
Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status

 , , , ,
, , , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Formulations
2.2. Rearing Conditions
2.3. Experimental Procedures
2.4. Hematological Procedures
2.5. Immune Parameters
2.6. Gene Expression Analysis
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Immune Parameters
3.3. Plasma Humoral Parameters
3.4. Skin Mucus Humoral Parameters
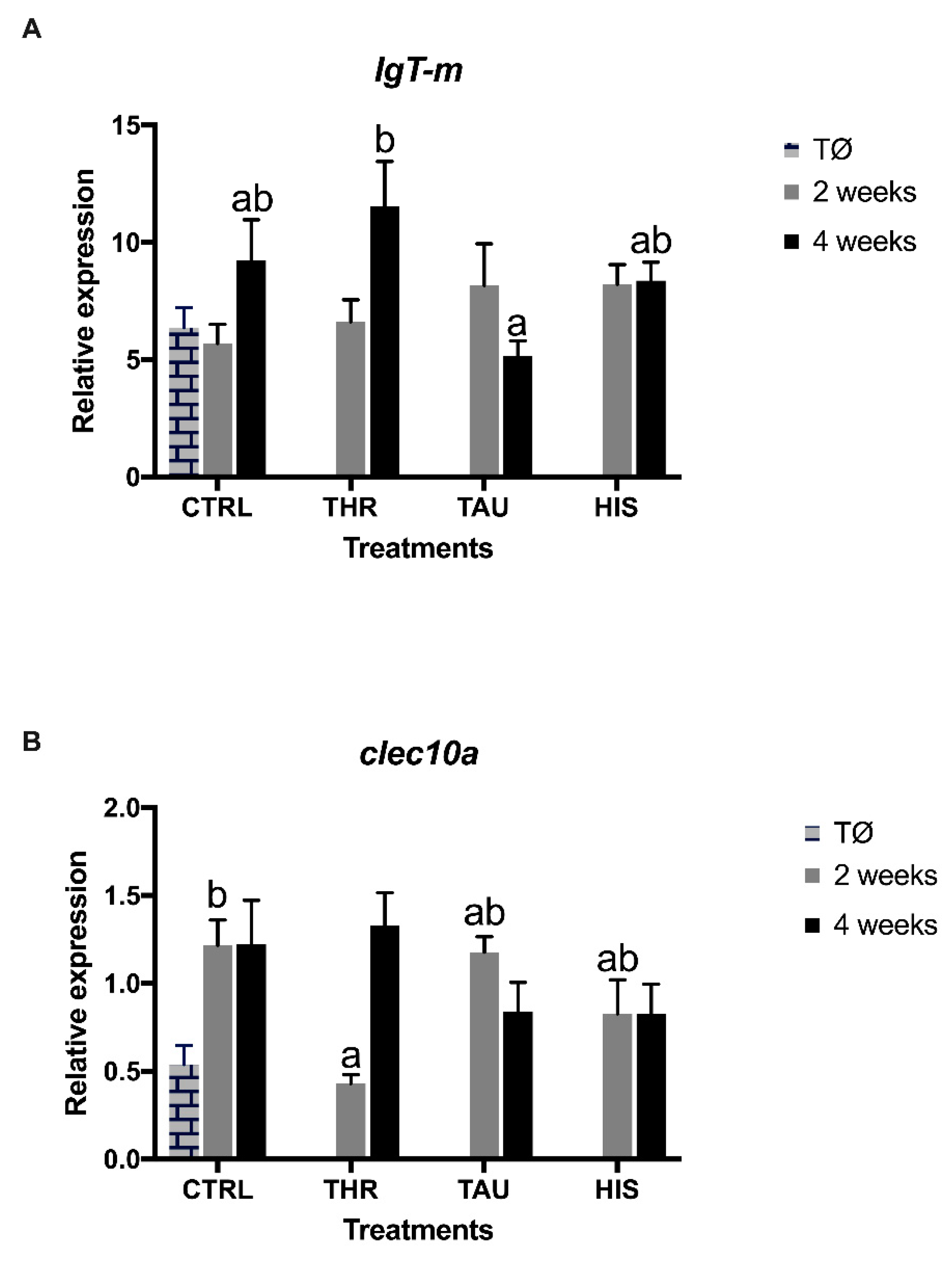
3.5. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quesada, S.P.; Paschoal, J.A.; Reyes, F.G. Considerations on the aquaculture development and on the use of veterinary drugs: Special issue for fluoroquinolones—A review. J. Food Sci. 2013, 78, R1321–R1333. [Google Scholar] [CrossRef]
- Oliva-Teles, A. Nutrition and health of aquaculture fish. J. Fish. Dis 2012, 35, 83–108. [Google Scholar] [CrossRef]
- Leal, M.C.; Calado, R. The key role of functional aquafeeds to achieve a more sustainable aquaculture. J. World Aquac. Soc. 2019, 50, 1044–1047. [Google Scholar] [CrossRef]
- Encarnação, P. 5-Functional feed additives in aquaculture feeds A2-Nates, Sergio, F. In Aquafeed Formulation; Academic Press: San Diego, CA, USA, 2016; pp. 217–237. [Google Scholar]
- Conceição, L.E.C.; Aragao, C.; Dias, J.; Costas, B.; Terova, G.; Martins, C.; Tort, L. Dietary nitrogen and fish welfare. Fish. Physiol Biochem. 2012, 38, 119–141. [Google Scholar] [CrossRef]
- Herrera, M.; Mancera, J.M.; Costas, B. The Use of Dietary Additives in Fish Stress Mitigation: Comparative Endocrine and Physiological Responses. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish. and Shrimp; The National Academies Press: Washington, DC, USA, 2011; p. 392. [Google Scholar]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Farhat; Khan, M.A. Effects of varying levels of dietary l-histidine on growth, feed conversion, protein gain, histidine retention, hematological and body composition in fingerling stinging catfish Heteropneustes fossilis (Bloch). Aquaculture 2013, 404–405, 130–138. [Google Scholar] [CrossRef]
- Waagbø, R.; Trosse, C.; Koppe, W.; Fontanillas, R.; Breck, O. Dietary histidine supplementation prevents cataract development in adult Atlantic salmon, Salmo salar L., in seawater. Br. J. Nutr. 2010, 104, 1460–1470. [Google Scholar] [CrossRef]
- Bender, D.A. Histidine. In Amino Acid Metabolism; John Wiley & Sons, Ltd.: New York, NY, USA, 2012; pp. 305–322. [Google Scholar]
- Lancha Junior, A.H.; Painelli Vde, S.; Saunders, B.; Artioli, G.G. Nutritional Strategies to Modulate Intracellular and Extracellular Buffering Capacity During High-Intensity Exercise. Sports Med. 2015, 45 Suppl 1, S71–S81. [Google Scholar] [CrossRef]
- Wade, A.M.; Tucker, H.N. Antioxidant characteristics of L-histidine 1. J. Nutr. Biochem. 1998, 9, 308–315. [Google Scholar] [CrossRef]
- Hobart, L.J.; Seibel, I.; Yeargans, G.S.; Seidler, N.W. Anti-crosslinking properties of carnosine: Significance of histidine. Life Sci. 2004, 75, 1379–1389. [Google Scholar] [CrossRef]
- Bellia, F.; Amorini, A.M.; La Mendola, D.; Vecchio, G.; Tavazzi, B.; Giardina, B.; Di Pietro, V.; Lazzarino, G.; Rizzarelli, E. New glycosidic derivatives of histidine-containing dipeptides with antioxidant properties and resistant to carnosinase activity. Eur. J. Med. Chem. 2008, 43, 373–380. [Google Scholar] [CrossRef]
- Gao, Y.-J.; Liu, Y.-J.; Chen, X.-Q.; Yang, H.-J.; Li, X.-F.; Tian, L.-X. Effects of graded levels of histidine on growth performance, digested enzymes activities, erythrocyte osmotic fragility and hypoxia-tolerance of juvenile grass carp Ctenopharyngodon idella. Aquaculture 2016, 452, 388–394. [Google Scholar] [CrossRef]
- Small, B.C.; Soares, J.H. Quantitative Dietary Threonine Requirement of Juvenile Striped Bass Morone saxatilis. J. World Aquac. Soc. 1999, 30, 319–323. [Google Scholar] [CrossRef]
- Tibaldi, E.; Tulli, F. Dietary threonine requirement of juvenile european sea bass (Dicentrarchus labrax). Aquaculture 1999, 175, 155–166. [Google Scholar] [CrossRef]
- Fatma Abidi, S.; A Khan, M. Dietary threonine requirement of fingerling Indian major carp, Labeo rohita (Hamilton). Aquac. Res. 2008, 39, 1498–1505. [Google Scholar] [CrossRef]
- Gao, Y.-J.; Yang, H.-J.; Liu, Y.-J.; Chen, S.-J.; Guo, D.-Q.; Yu, Y.-y.; Tian, L.-X. Effects of graded levels of threonine on growth performance, biochemical parameters and intestine morphology of juvenile grass carp Ctenopharyngodon idella. Aquaculture 2014, 424–425, 113–119. [Google Scholar] [CrossRef]
- Duval, D.; Demangel, C.; Munier-Jolain, K.; Miossec, S.; Geahel, I. Factors controlling cell proliferation and antibody production in mouse hybridoma cells: I. Influence of the amino acid supply. Biotechnol. Bioeng. 1991, 38, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Corzo, A.; Kidd, M.T.; Dozier, W.A.; Pharr, G.T.; Koutsos, E.A. Dietary Threonine Needs for Growth and Immunity of Broilers Raised Under Different Litter Conditions. J. Appl Poult Res. 2007, 16, 574–582. [Google Scholar] [CrossRef]
- Hong, Y.; Jiang, W.; Kuang, S.; Hu, K.; Tang, L.; Liu, Y.; Jiang, J.; Zhang, Y.; Zhou, X.; Feng, L. Growth, digestive and absorptive capacity and antioxidant status in intestine and hepatopancreas of sub-adult grass carp Ctenopharyngodonidella fed graded levels of dietary threonine. J. Anim. Sci. Biotechnol. 2015, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Faure, M.; Moennoz, D.; Montigon, F.; Mettraux, C.; Breuille, D.; Ballevre, O. Dietary threonine restriction specifically reduces intestinal mucin. J. Nutr. 2005, 135. [Google Scholar] [CrossRef]
- Feng, L.; Peng, Y.; Wu, P.; Hu, K.; Jiang, W.D.; Liu, Y. Threonine affects intestinal function, protein synthesis and gene expression of TOR in Jian carp (Cyprinus carpio var. Jian). PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Salze, G.P.; Davis, D.A. Taurine: A critical nutrient for future fish feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- Goto, T.; Matsumoto, T.; Murakami, S.; Takagi, S.; Hasumi, F. Conversion of cysteate into taurine in liver of fish. Fisheries Sci. 2003, 69, 216–218. [Google Scholar] [CrossRef]
- Gibson, G.T.; Barrows, F.T.; Teague, A.M.; Johansen, K.A.; Overturf, K.E.; Shepherd, B. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 269, 514–524. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Hayashi, M.; Hatate, H.; Endo, M.; Yamashita, H.; Ukawa, M. Hemolytic suppression roles of taurine in yellowtail Seriola quinqueradiata fed non-fishmeal diet based on soybean protein. Fisheries Sci. 2006, 72, 546–555. [Google Scholar] [CrossRef]
- Maita, M.; Maekawa, J.; Satoh, K.-i.; Futami, K.; Satoh, S. Disease resistance and hypocholesterolemia in yellowtail Seriola quinqueradiata fed a non-fishmeal diet. Fisheries Sci. 2006, 72, 513–519. [Google Scholar] [CrossRef]
- Colorni, A.P.F. Diseases and health management. In Sparidae: Biology and Aquaculture of Gilthead Sea Bream and other Species; Pavlidis, M.A., Mylonas, C.C., Eds.; Wiley-Blackwell: Oxford, UK, 2011. [Google Scholar]
- Ramos-Pinto, L.; Azeredo, R.; Silva, C.; Conceição, L.E.C.; Dias, J.; Montero, D.; Torrecillas, S.; Silva, T.S.; Costas, B. Short-Term Supplementation of Dietary Arginine and Citrulline Modulates Gilthead Seabream (Sparus aurata) Immune Status. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Pinto, L.; Martos-Sitcha, J.A.; Reis, B.; Azeredo, R.; Fernandez-Boo, S.; Pérez-Sánchez, J.; Calduch-Giner, J.A.; Engrola, S.; Conceição, L.E.C.; Dias, J.; et al. Dietary tryptophan supplementation induces a transient immune enhancement of gilthead seabream (Sparus aurata) juveniles fed fishmeal-free diets. Fish Shellfish Immunol. 2019, 93, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.J. Whole body amino acid composition of European seabass (Dicentrarchus labrax), gilthead seabream (Sparus aurata) and turbot (Psetta maxima) with an estimation of their IAA requirement profiles. Aquat. Living Resour. 1998, 11, 355–358. [Google Scholar] [CrossRef]
- Peres, H.; Oliva-Teles, A. The optimum dietary essential amino acid profile for gilthead seabream (Sparus aurata) juveniles. Aquaculture 2009, 296, 81–86. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Diaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef]
- Afonso, A.; Lousada, S.; Silva, J.; Ellis, A.E.; Silva, M.T. Neutrophil and macrophage responses to inflammation in the peritoneal cavity of rainbow trout Oncorhynchus mykiss. A light and electron microscopic cytochemical study. Dis Aquat Organ. 1998, 34, 27–37. [Google Scholar] [CrossRef]
- Graham, S.; Jeffries, A.H.; Secombes, C.J. A novel assay to detect macrophage bactericidal activity in fish: Factors influencing the killing of Aeromonas salmonicida. J. Fish Dis. 1988, 11, 389–396. [Google Scholar] [CrossRef]
- Ellis, A.E. Serum antiproteases in fish. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., van Muiswinkel, W.B., Eds.; SOS Publications: Fair Haven, NJ, USA, 1990. [Google Scholar]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunop. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemometr. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Li, H.; Ma, M.-L.; Luo, S.; Zhang, R.-M.; Han, P.; Hu, W. Metabolic responses to ethanol in Saccharomyces cerevisiae using a gas chromatography tandem mass spectrometry-based metabolomics approach. Int. J. Biochem. Cell Biol. 2012, 44, 1087–1096. [Google Scholar] [CrossRef]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; Martin, R.J.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol. Renal. Physiol. 2016, 310, F857–F871. [Google Scholar] [CrossRef] [PubMed]
- Bouckenooghe, T.; Remacle, C.; Reusens, B. Is taurine a functional nutrient? Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 728–733. [Google Scholar] [CrossRef]
- do Vale, A.; Afonso, A.; Silva, M.T. The professional phagocytes of sea bass (Dicentrarchus labrax L.): Cytochemical characterisation of neutrophils and macrophages in the normal and inflamed peritoneal cavity. Fish Shellfish Immunol. 2002, 13, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M.; Kato, I. Protein Inhibitors of Proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Agbowuro, A.A.; Huston, W.M.; Gamble, A.B.; Tyndall, J.D.A. Proteases and protease inhibitors in infectious diseases. Med. Res. Rev. 2018, 38, 1295–1331. [Google Scholar] [CrossRef]
- Gonzalez-Silvera, D.; Herrera, M.; Giráldez, I.; Esteban, M. Effects of the Dietary Tryptophan and Aspartate on the Immune Response of Meagre (Argyrosomus regius) after Stress. Fishes 2018, 3, 6. [Google Scholar] [CrossRef]
- Potempa, J.; Pike, R.N. Corruption of innate immunity by bacterial proteases. J. Innate. Immun. 2009, 1, 70–87. [Google Scholar] [CrossRef]
- Dash, S.; Das, S.K.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar]
- Ellis, A.E. Immunity to bacteria in fish. Fish Shellfish Immunol. 1999, 9, 291–308. [Google Scholar] [CrossRef]
- Han, Y.; Koshio, S.; Jiang, Z.; Ren, T.; Ishikawa, M.; Yokoyama, S.; Gao, J. Interactive effects of dietary taurine and glutamine on growth performance, blood parameters and oxidative status of Japanese flounder Paralichthys olivaceus. Aquaculture 2014, 434, 348–354. [Google Scholar] [CrossRef]
- Magalhães, R.; Martins, N.; Martins, S.; Lopes, T.; Diáz-Rosales, P.; Pousão-Ferreira, P.; Oliva-Teles, A.; Peres, H. Is dietary taurine required for white seabream (Diplodus sargus) juveniles? Aquaculture 2019, 502, 296–302. [Google Scholar] [CrossRef]
- Li, M.; Lai, H.; Li, Q.; Gong, S.; Wang, R. Effects of dietary taurine on growth, immunity and hyperammonemia in juvenile yellow catfish Pelteobagrus fulvidraco fed all-plant protein diets. Aquaculture 2016, 450, 349–355. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Trede, N.S. Fish immunology. Curr. Biol. 2009, 19, R678–R682. [Google Scholar] [CrossRef]
- Perez-Sanchez, J.; Benedito-Palos, L.; Estensoro, I.; Petropoulos, Y.; Calduch-Giner, J.A.; Browdy, C.L.; Sitja-Bobadilla, A. Effects of dietary NEXT ENHANCE(R)150 on growth performance and expression of immune and intestinal integrity related genes in gilthead sea bream (Sparus aurata L.). Fish Shellfish Immunol. 2015, 44, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Vasta, G.R. Roles of galectins in infection. Nat. Rev. Microbiol. 2009, 7, 424–438. [Google Scholar] [CrossRef]
- Vasta, G.R.; Ahmed, H.; Odom, E.W. Structural and functional diversity of lectin repertoires in invertebrates, protochordates and ectothermic vertebrates. Curr. Opin. Struct. Biol. 2004, 14, 617–630. [Google Scholar] [CrossRef]
- Mayer, S.; Raulf, M.K.; Lepenies, B. C-type lectins: Their network and roles in pathogen recognition and immunity. Histochem. Cell Biol. 2017, 147, 223–237. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Fontinha, F.; Fernandez-Boo, S.; Conceicao, L.E.C.; Dias, J.; Costas, B. Dietary Methionine Improves the European Seabass (Dicentrarchus labrax) Immune Status, Inflammatory Response, and Disease Resistance. Front. Immunol. 2018, 9, 2672. [Google Scholar] [CrossRef]
- Azeredo, R.; Machado, M.; Fontinha, F.; Fernández-Boo, S.; Conceição, L.E.C.; Dias, J.; Costas, B. Dietary arginine and citrulline supplementation seem to negatively modulate the immune condition and inflammatory response of the European seabass. Fish Shellfish Immunol. 2020, 106, 451–463. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Ingredients (% Feed Basis) | Experimental Diets | |||
|---|---|---|---|---|
| CTRL | HIS | TAU | THR | |
| Fishmeal LT70 (NORVIK) a | 12.00 | 12.00 | 12.00 | 12.00 |
| Poultry meal 65 b | 5.00 | 5.00 | 5.00 | 5.00 |
| Soy protein concentrate (Soycomil) c | 8.50 | 8.50 | 8.50 | 8.50 |
| Wheat gluten d | 7.37 | 7.37 | 7.37 | 7.37 |
| Corn gluten e | 8.00 | 8.00 | 8.00 | 8.00 |
| Soybean meal 48 f | 7.68 | 7.68 | 7.68 | 7.68 |
| Soybean meal 44 g | 15.00 | 15.00 | 15.00 | 15.00 |
| Rapeseed meal h | 5.00 | 5.00 | 5.00 | 5.00 |
| Wheat meal i | 14.30 | 14.30 | 14.30 | 14.30 |
| Sardine oil j | 4.65 | 4.65 | 4.65 | 4.65 |
| Rapeseed oil k | 10.85 | 10.85 | 10.85 | 10.85 |
| Vit & Min Premix INVIVO 1% l | 1.00 | 1.00 | 1.00 | 1.00 |
| Antioxidant m | 0.20 | 0.20 | 0.20 | 0.20 |
| Sodium propionate n | 0.10 | 0.10 | 0.10 | 0.10 |
| MCP o | 0.20 | 0.20 | 0.20 | 0.20 |
| DL-Methionine p | 0.15 | 0.15 | 0.15 | 0.15 |
| L-Threonine q | 0.75 | |||
| L-Histidine r | 0.40 | |||
| Taurine s | 0.50 | |||
| Proximate analyses | ||||
| Dry matter (% feed) | 94.6 | 94.3 | 94.2 | 94.3 |
| Crude protein (% dry weight) | 42.62 | 41.92 | 42.24 | 42.23 |
| Crude lipid (% dry weight) | 18.3 | 19.1 | 18 | 18.3 |
| Ash (% dry weight) | 6.4 | 6.1 | 6.3 | 6.1 |
| Gross Energy (kJ g−1 DM) | 21.83 | 21.75 | 21.37 | 22.00 |
| AA | Experimental Diets | |||
|---|---|---|---|---|
| CTRL | HIS | TAU | THR | |
| Arginine | 7.6 | 6.1 | 6.3 | 5.7 |
| Histidine | 2.1 | 3.4 | 2.6 | 2.4 |
| Lysine | 5.3 | 6.3 | 6.7 | 6.2 |
| Threonine | 3.8 | 3.5 | 3.6 | 4.3 |
| Isoleucine | 4.5 | 4.2 | 4.3 | 4.1 |
| Leucine | 8.4 | 7.2 | 7.1 | 6.9 |
| Valine | 4.8 | 4.9 | 5.0 | 4.7 |
| Methionine | 2.3 | 2.5 | 2.9 | 2.2 |
| Phenylalanine | 5.3 | 4.0 | 4.0 | 3.8 |
| Cystine | 0.6 | 1.3 | 1.3 | 1.3 |
| Tyrosine | 4.0 | 3.4 | 3.5 | 3.1 |
| Aspartic acid + Asparagine | 7.4 | 9.6 | 10.1 | 9.5 |
| Glutamic acid + Glutamine | 19.0 | 21.8 | 19.0 | 21.9 |
| Alanine | 4.8 | 4.8 | 5.0 | 4.6 |
| Glycine | 5.1 | 4.4 | 4.9 | 4.2 |
| Proline | 6.7 | 6.6 | 6.5 | 6.4 |
| Serine | 4.5 | 4.4 | 4.5 | 4.4 |
| Taurine | 0.3 | 0.3 | 1.0 | 0.2 |
| Gene Name/Category | Symbol | Gene Name/Category | Symbol |
|---|---|---|---|
| Interleukins and cytokines | T-cell markers | ||
| Interleukin-1 beta | il-1β | Cluster of differentiation 3 zeta chain | cd3x |
| Interleukin-6 | il-6 | CD4-full | cd4-full |
| Interleukin-7 | il-7 | Cluster of differentiation 8 alpha | cd8a |
| Interleukin-8 | il-8 | Cluster of differentiation 8 beta | cd8b |
| Interleukin-10 | il-10 | Zeta-chain-associated protein kinase 70 | zap70 |
| Interleukin 12 subunit beta | il12 | ||
| Interleukin-15 | il-15 | ||
| Interleukin-34 | il-34 | Pattern recognition receptors | |
| Tumor necrosis factor-alpha | tnf α | Toll-like receptor 2 | tlr2 |
| Toll-like receptor 5 | tlr5 | ||
| Macrophages and monocytes chemokines | Toll-like receptor 9 | tlr9 | |
| Macrophage colony-stimulating factor 1 receptor 1 | csf1r1 | Macrophage mannose receptor 1 | mrc1 |
| C-C chemokine receptor type 3 | ccr3 | ||
| C-C chemokine CK8/C-C motif chemokine 20 | ck8/ccl20 | Antimicrobial peptide/Iron recycling | |
| Hepcidin | hepc | ||
| Immunoglobulins | |||
| Immunoglobulin M | sIgM | Complement pathways | |
| Immunoglobulin M membrane-bound form | IgM-m | Complement factor 3 | c3 |
| Immunoglobulin T | sIgT | ||
| Immunoglobulin T membrane-bound form | IgT-m | C-type lectin receptor signaling | |
| C-type lectin domain family 10 member A | clec10a | ||
| Apoptosis | |||
| Caspase 3 | casp3 | ||
| Parameters | CTRL | THR | TAU | HIS | ||||||
| 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | |||
| FBW (g fish-1) | 11.07 ± 0.23 | 14.49 ± 2.10 | 10.89 ± 0.30 | 14.04 ± 1.49 | 11.08 ± 0.33 | 13.89 ± 1.08 | 11.13 ± 0.49 | 13.70 ± 0.10 | ||
| RGR (RGR, % day-1) | 1.63 ± 0.28 | 1.76 ± 0.50 | 1.65 ± 0.51 | 1.73 ± 0.35 | 1.54 ± 0.18 | 1.58 ± 0.32 | 1.84 ± 0.48 | 1.67 ± 0.09 | ||
| Two-way ANOVA | ||||||||||
| Parameters | Diet | Time | Time × diet | Time | Diet | |||||
| 2 weeks | 4 weeks | CTRL | THR | TAU | HIS | |||||
| BW | 0.925 | <0.001 | 0.893 | A | B | - | - | - | - | |
| RGR | 0.814 | 0.911 | 0.898 | - | - | - | - | - | - | |
| Parameters | CTRL | CTRL | THR | TAU | HIS | |||||||
| TØ | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | ||||
| Hemoglobin | (g dL−1) | 0.60 ± 0.10 | 0.60 ± 0.06 | 0.60 ± 0.05 | 0.65 ± 0.09 | 0.65 ± 0.04 | 0.57 ± 0.03 | 0.56 ± 0.05 | 0.54 ± 0.06 | 0.67 ± 0.10 | ||
| WBC | (×104 µL) | 4.66 ± 0.33 b | 4.53 ± 0.60 b | 6.21 ± 0.36 | 3.34 ± 0.20 a | 7.11 ± 0.56 | 4.85 ± 0.28 b | 7.00 ± 0.40 | 4.94 ± 0.47 b | 5.94 ± 0.56 | ||
| RBC | (×106 µL) | 1.55 ± 0.15 | 1.14 ± 0.09 | 1.44 ± 0.10 | 1.28 ± 0.11 | 1.58 ± 0.04 | 1.00 ± 0.06 | 1.42 ± 0.09 | 1.24 ± 0.11 | 1.66 ± 0.12 | ||
| Absolute peripheral blood leucocytes | ||||||||||||
| Thrombocytes | (×104 µL) | 3.31 ± 0.23 | 3.46 ± 0.57 | 4.54 ± 0.26 | 2.46 ± 0.18 | 4.77 ± 0.36 | 3.62 ± 0.27 | 5.19 ± 0.40 | 3.56 ± 0.35 | 4.60 ± 0.47 | ||
| Lymphocytes | (×104 µL) | 1.21 ± 0.23 ab | 0.93 ± 0.14 | 1.37 ± 0.18 ab | 0.70 ± 0.34 * | 1.95 ± 0.49 b# | 0.98 ± 0.46 | 1.49 ± 0.48 ab | 1.22 ± 0.49 | 1.11 ± 0.43 a | ||
| Monocytes | (×104 µL) | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.06 ± 0.02 | 0.02 ± 0.01 | 0.06 ± 0.02 | 0.04 ± 0.01 | 0.08 ± 0.02 | 0.07 ± 0.02 | 0.06 ± 0.02 | ||
| Neutrophils | (×104 µL) | 0.13 ± 0.03 | 0.07 ± 0.02 | 0.22 ± 0.04 | 0.08 ± 0.03 | 0.18 ± 0.02 | 0.18 ± 0.03 | 0.25 ± 0.02 | 0.09 ± 0.01 | 0.15 ± 0.04 | ||
| Two-way ANOVA | ||||||||||||
| Parameters | Diet | Time | Time × diet | Time | Diet | |||||||
| TØ | 2 weeks | 4 weeks | CTRL | THR | TAU | HIS | ||||||
| Hemoglobin | (g dl −1) | 0.633 | 0.538 | 0.699 | - | - | - | - | - | - | - | |
| WBC | (×104 µL) | 0.406 | <0.001 | 0.017 | A | A | B | - | - | - | - | |
| RBC | (×106 µL) | 0.286 | <0.001 | 0.898 | B | A | B | - | - | - | - | |
| Absolute peripheral blood leucocytes | ||||||||||||
| Thrombocytes | (×104 µL) | 0.214 | <0.001 | 0.298 | A | A | B | - | - | - | - | |
| Lymphocytes | (×104 µL) | 0.74 | <0.001 | 0.003 | AB | A | B | - | - | - | - | |
| Monocytes | (×104 µL) | 0.071 | 0.003 | 0.116 | AB | A | B | - | - | - | - | |
| Neutrophils | (×104 µL) | 0.006 | <0.001 | 0.287 | A | A | B | AB | AB | B | A | |
| Parameters | CTRL | CTRL | THR | TAU | HIS | ||||||||
| TØ | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | 2 Weeks | 4 Weeks | |||||
| Plasma | |||||||||||||
| Bactericidal activity | (%) | 28.57 ± 7.31 | 43.75 ± 2.14 | 47.56 ± 3.01 | 37.65 ± 2.56 | 48.25 ± 2.76 | 38.47 ± 2.20 | 39.92 ± 3.30 | 42.39 ± 3.40 | 43.54 ± 2.90 | |||
| Antiprotease | (%) | ⨂ | 92.02 ± 0.99 | 92.69 ± 1.43 | 94.60 ± 0.59 | 95.45 ± 1.01 | 90.06 ± 0.85 | 94.56 ± 0.59 | 91.71 ± 1.04 | 95.97 ± 0.50 | |||
| Peroxidase | (U/mL) | ⨂ | 6.29 ± 0.71 | 7.07 ± 0.71 | 6.60 ± 0.79 | 7.18 ± 0.55 | 6.34 ± 0.64 | 6.44 ± 0.43 | 6.95 ± 0.43 | 7.94 ± 1.00 | |||
| IgM | (abs) | 0.80 ± 0.11 | 0.85 ± 0.02 | 0.72 ± 0.05 | 0.77 ± 0.04 | 0.68 ± 0.06 | 0.66 ± 0.04 | 0.74 ± 0.04 | 0.81 ± 0.05 | 0.66 ± 0.05 | |||
| Mucus | |||||||||||||
| Bactericidal activity | (%) | 60.53 ± 1.02 a | 66.41 ± 0.63 *b | 55.83 ± 1.76 # | 64.57 ± 0.89 ab | 59.11 ± 1.59 | 60.80 ± 1.34 a | 59.08 ± 1.19 | 59.40 ± 1.26 a | 57.69 ± 1.49 | |||
| Antiprotease | (%) | 29.45 ± 2.37 | 25.52 ± 2.70 | 26.99 ± 1.33 | 28.10 ± 1.97 | 26.88 ± 1.31 | 28.36 ± 2.10 | 28.69 ± 2.49 | 27.52 ± 0.90 | 30.62 ± 2.6 | |||
| Peroxidase activity | (U/mg) | 0.66 ± 0.06 | 0.73 ± 0.09 | 0.69 ± 0.09 | 0.89 ± 0.10 | 0.78 ± 0.11 | 1.00 ± 0.12 | 0.71 ± 0.08 | 0.81 ± 0.11 | 0.78 ± 0.41 | |||
| Two-way ANOVA | |||||||||||||
| Parameters | Diet | Time | Time × diet | Time | Diet | ||||||||
| TØ | 2 weeks | 4 weeks | CTRL | THR | TAU | HIS | |||||||
| Plasma | |||||||||||||
| Bactericidal activity | (%) | 0.151 | 0.037 | 0.343 | A | AB | B | - | - | - | - | ||
| Antiprotease | (%) | 0.013 | <0.001 | 0.059 | - | A | B | A | B | A | AB | ||
| Peroxidase | (U/mL) | 0.471 | 0.208 | 0.926 | - | - | - | - | - | - | - | ||
| IgM | (abs) | 0.302 | 0.023 | 0.045 | B | B | A | - | - | - | - | ||
| Mucus | |||||||||||||
| Bactericidal activity | (%) | 0.053 | <0.001 | 0.001 | AB | B | A | - | - | - | - | ||
| Antiprotease | (%) | 0.012 | 0.149 | 0.102 | - | - | - | A | AB | AB | B | ||
| Peroxidase activity | (U/mg) | 0.452 | 0.1 | 0.541 | - | - | - | - | - | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Pinto, L.; Machado, M.; Calduch-Giner, J.; Pérez-Sánchez, J.; Dias, J.; Conceição, L.E.C.; Silva, T.S.; Costas, B. Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status. Animals 2021, 11, 1193. https://doi.org/10.3390/ani11051193
Ramos-Pinto L, Machado M, Calduch-Giner J, Pérez-Sánchez J, Dias J, Conceição LEC, Silva TS, Costas B. Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status. Animals. 2021; 11(5):1193. https://doi.org/10.3390/ani11051193
Chicago/Turabian StyleRamos-Pinto, Lourenço, Marina Machado, Josep Calduch-Giner, Jaume Pérez-Sánchez, Jorge Dias, Luís E. C. Conceição, Tomé S. Silva, and Benjamín Costas. 2021. "Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status" Animals 11, no. 5: 1193. https://doi.org/10.3390/ani11051193
APA StyleRamos-Pinto, L., Machado, M., Calduch-Giner, J., Pérez-Sánchez, J., Dias, J., Conceição, L. E. C., Silva, T. S., & Costas, B. (2021). Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status. Animals, 11(5), 1193. https://doi.org/10.3390/ani11051193