
Farm Animals Are Long Away from Natural Behavior: Open Questions and Operative Consequences on Animal Welfare



Abstract
Simple Summary
Abstract
1. Introduction
2. Domestication Changed Animal Traits
3. Questions and Answers about Modern Farm Animals
4. Practical Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Donnell, P. Animals: Ethics, Rights & Law—A Transdisciplinary Bibliography. 2019. Available online: https://www.academia.edu/4843888/Animal_Ethics_Rights_and_Law_bibliography (accessed on 29 December 2020).
- The Future of Animal-Sourced Foods: Ethical Considerations. Anim. Front. 2020, 10, 1. [CrossRef]
- Pulina, G. Ethical meat: Respect for farm animals. Anim. Front. 2020, 10, 34–38. [Google Scholar] [CrossRef]
- European Commission. Attitudes of Europeans towards Animal Welfare. Spec. Eurobarometer 2016, 442, 22. [Google Scholar]
- Cambridge Dictionary. 2020. Available online: https://dictionary.cambridge.org/dictionary/english/welfare (accessed on 30 December 2020).
- Carenzi, C.; Verga, M. Animal welfare: Review of the scientific concept and definition. Ital. J. Anim. Sci. 2009, 8, 21–30. [Google Scholar] [CrossRef]
- Darwin, C. The Expression of Emotions in Man and Animals; Murray, J., Ed.; Oxford University Press: London, UK, 1872. [Google Scholar]
- Romanes, G. Animal Intelligence. Nature 1884, 30, 267. [Google Scholar] [CrossRef]
- Knoll, E. Dogs, darwinism and english sensibilities. In Anthropomorphism, Anecdotes, and Animals; Mitchell, R.W., Thompson, N.S., Miles, H.L., Eds.; State University of New York Press: Albany, NY, USA, 1997; pp. 12–21. [Google Scholar]
- LeDoux, J.E. The Deep History of Ourselves: The Four Billion Years Story of How We Got Conscious Brains; Ed. Penguin Books: New York, NY, USA, 2019. [Google Scholar]
- EU. Treaty of Lisbon, amending the treaty on European Union and the treaty establishing the European community. OJEU 2007, 17, 13. [Google Scholar]
- Morgante, M.; Vallortigara, G. Animal welfare: Neurocognitive approaches. Ital. J. Anim. Sci. 2009, 8 (Suppl. 1), 255–264. [Google Scholar] [CrossRef]
- Vallortigara, G. Sentience does not require “higher” cognition. Anim. Sentience 2017, 30, 6. [Google Scholar]
- Désiré, L.; Boissy, A.; Veissier, I. Emotions in farm animals: A new approach to animal welfare in applied ethology. Behav. Process 2002, 60, 165–180. [Google Scholar] [CrossRef]
- De Waal, F.B.M. What is an animal emotion? Ann. N. Y. Acad. Sci. 2011, 1224, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Belyaev, D.K. Destabilizing selection as a factor in domestication. J. Hered. 1979, 70, 301–308. [Google Scholar] [CrossRef]
- Kukekova, A.V.; Johnson, J.L.; Xiang, X.; Feng, S.; Liu, S.; Rando, H.M.; Kharlamova, A.V.; Herbeck, Y.; Serdyukova, N.A.; Xiong, Z.; et al. Red fox genome assembly identifies genomic regions associated with tame and aggressive behaviours. Nat. Ecol. Evol. 2018, 2, 1479–1491. [Google Scholar] [CrossRef]
- Price, E.O. Behavioral development in animals undergoing domestication. Appl. Anim. Behav. Sci. 1999, 65, 245–271. [Google Scholar] [CrossRef]
- Zeder, M.A. Core questions in domestication research. Proc. Natl. Acad. Sci. USA 2015, 112, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. The Origin of Species, 6th ed.; John Murray: London, UK, 1859; Volume 570. [Google Scholar]
- Vonholdt, B.M.; Pollinger, J.P.; Lohmueller, K.E.; Han, E.; Parker, H.G.; Quignon, P.; Degenhardt, J.D.; Boyko, A.R.; Earl, D.A.; Auton, A.; et al. Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication. Nature 2010, 464, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Cagan, A.; Blass, T. Identification of genomic variants putatively targeted by selection during dog domestication. BMC Evol. Biol. 2016, 16, 10. [Google Scholar] [CrossRef]
- MacHugh, D.E.; Larson, G.; Orlando, L. Taming the past: Ancient DNA and the study of animal domestication. Ann. Rev. Anim. Biosci. 2017, 5, 329–351. [Google Scholar] [CrossRef]
- Conolly, J.; Colledge, S.; Dobney, K.; Vigne, J.D.; Peters, J.; Stopp, B.; Manning, K.; Shennan, S. Meta-analysis of zooarchaeological data from SW Asia and SE Europe provides insight into the origins and spread of animal husbandry. J. Archaeol. Sci. 2011, 38, 538–545. [Google Scholar] [CrossRef]
- Asouti, E.; Fuller, D.Q. A contextual approach to the emergence of agriculture in Southwest Asia reconstructing early Neolithic plant-food production. Curr. Anthr. 2013, 54, 299–345. [Google Scholar] [CrossRef]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Vigueira, C.C.; Denham, T.; Dobney, K.; et al. Current perspectives and the future of domestication studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef] [PubMed]
- Gepts, P.; Papa, R. Evolution during domestication. In Encyclopedia of Life Sciences; Nature Publishing Group, Ed.; Macmillan Publishers: New York, NY, USA, 2001. [Google Scholar]
- Clutton-Brock, J.A. Natural History of Domesticated Mammals; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Carneiro, M.; Afonso, S.; Geraldes, A.; Garreau, H.; Bolet, G.; Boucher, S.; Tircazes, A.; Queney, G.; Nachman, M.W.; Ferrand, N. The genetic structure of domestic rabbits. Mol. Biol. Evol. 2011, 28, 1801–1816. [Google Scholar] [CrossRef] [PubMed]
- Rasali, D.P.; Shrestha, J.N.B.; Crow, G.H. Development of composite sheep breeds in the world: A review. Can. J. Anim. Sci. 2008, 86, 1–24. [Google Scholar] [CrossRef]
- Felius, M.; Beerling, M.L.; Buchanan, D.S.; Theunissen, B.; Koolmees, P.A.; Lenstra, J.A. On the history of cattle genetic resources. Diversity 2014, 6, 705–750. [Google Scholar] [CrossRef]
- FAO. Status of Animal Genetic Resources. Available online: www.fao.org/docrep/pdf/010/a1250e/a1250e02.pdf (accessed on 2 December 2020).
- Ryder, M.L. Sheep and Man; Gerald Duckworth & Co. Ltd.: London, UK, 1983. [Google Scholar]
- Loftus, R.T.; MacHugh, D.E.; Bradley, D.G.; Sharp, P.M.; Cunningham, P. Evidence for two independent domestications of cattle. Proc. Natl. Acad. Sci. USA 1994, 91, 2757–2761. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, M.R.; Chen, W.; Lenstra, J.A.; Goderie, C.R.J.; MacHugh, D.E.; Park, S.D.E.; Magee, D.A.; Matassino, D.; Ciani, F.; Megens, H.J.; et al. Genetic origin, admixture and population history of aurochs (Bos primigenius) and primitive European cattle. Heredity 2016, 118, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zeder, M. Domestication and early agriculture in the Mediterranean basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11592–11604. [Google Scholar] [CrossRef] [PubMed]
- Harri, M.; Mononen, J.; Ahola, L.; Plyusnina, I.; Rekilä, T. Behavioural and physiological differences between silver foxes selected and not selected for domestic behaviour. Anim. Welf. 2003, 12, 305–314. [Google Scholar]
- Thrope, W.H. The Origins and Rise of Ethology; Praeger: London, UK, 1979. [Google Scholar]
- Jensen, P. Domestication—From behaviour to genes and back again. Appl. Anim. Behav. Sci. 2006, 97, 3–15. [Google Scholar] [CrossRef]
- Kumar, A.; Faiq, M.A.; Pareek, V.; Kulandhasamy, M. Heritability of Behavior. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T.K., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Dochtermann, N.A.; Schwab, T.; Anderson Berdal, M.; Dalos, J.; Royauté, R. The heritability of behavior: A meta-analysis. J. Hered. 2019, 110, 403–410. [Google Scholar] [CrossRef]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “domestication syndrome” in mammals: A unified explanation based on neural crest cell behavior and genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.S. A striking example of developmental bias in an evolutionary process: The “domestication syndrome”. Evol. Dev. 2020, 22, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.A.; Jones, M.K.; Powell, W.; Allaby, R.G. The complex origins of domesticated crops in the Fertile Crescent. Trends Ecol. Evol. 2009, 24, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Villagra, M.R.; Geiger, M.; Schneider, R.A. The taming of the neural crest: A developmental perspective on the origins of morphological covariation in domesticated mammals. R. Soc. Open Sci. 2016, 3, 160107. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M.J.; Price, E.G. Comparative evolution of cereals. Evolution 1973, 27, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Greenspan, R.J. The nature of genetic influences on behavior: Lessons from “simpler” organisms. Am. J. Psychiatry 2006, 163, 1683–1694. [Google Scholar] [CrossRef]
- Jensen, P. Behavior genetics and the domestication of animals. Annu. Rev. Anim. Biosci. 2014, 2, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Price, E.O. Animal Domestication and Behavior; CABI Publishers: Wallingford, UK, 2002. [Google Scholar]
- York, R.A. Assessing the genetic landscape of animal behavior. Genetics 2018, 209, 223–232. [Google Scholar] [CrossRef]
- Cesarani, A.; Sechi, T.; Gaspa, G.; Usai, M.G.; Sorbolini, S.; Macciotta, N.P.P.; Carta, A. Investigation of genetic diversity and selection signatures between Sarda and Sardinian Ancestral black, two related sheep breeds with evident morphological differences. Small Rumin. Res. 2019, 177, 68–75. [Google Scholar] [CrossRef]
- Luquetti, D.V.; Heike, C.L.; Hing, A.V.; Cunningham, M.L.; Cox, T.C. Microtia: Epidemiology and genetics. Am. J. Med. Genet. 2012, 158A, 124–139. [Google Scholar] [CrossRef]
- Bartel-Friedrich, S. Congenital auricular malformations: Description of anomalies and syndromes. Facial Plast. Surg. 2015, 31, 567–580. [Google Scholar] [CrossRef]
- Soundararajan, C.; Nagarajan, K.; Prakash, M.A. Occurrence of Microtia in Madras red Sheep-A study of 12 flocks. Intas. Polivet. 2016, 17, 255–257. [Google Scholar]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Neto, L.R.P.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-wide analysis of the world’s sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [PubMed]
- Saetre, P.; Lindberg, J.; Leonard, J.A.; Olsson, K.; Pettersson, U.; Ellegren, H.; Bergström, T.F.; Vilà, C.; Jazin, E. From wild wolf to domestic dog: Gene expression changes in the brain. Brain Res. Mol. Brain Res. 2004, 126, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Albert, F.W.; Somel, M.; Carneiro, M.; Aximu-Petri, A.; Halbwax, M.; Thalmann, O.; Blanco-Aguiar, J.A.; Plyusnina, I.Z.; Trut, L.; Villafuerte, R.; et al. A comparison of brain gene expression levels in domesticated and wild animals. PLoS Genet. 2012, 8, e1002962. [Google Scholar] [CrossRef]
- Nätt, D.; Rubin, C.J.; Wright, D.; Johnsson, M.; Beltéky, J.; Andersson, L.; Jensen, P. Heritable genome-wide variation of gene expression and promoter methylation between wild and domesticated chickens. BMC Genom. 2012, 13, 1–12. [Google Scholar] [CrossRef]
- Carneiro, M.; Piorno, V.; Rubin, C.J.; Alves, J.M.; Ferrand, N.; Alves, P.C.; Andersson, L. Candidate genes underlying heritable differences in reproductive seasonality between wild and domestic rabbits. Anim. Genet. 2015, 46, 418–425. [Google Scholar] [CrossRef]
- Christie, M.; Marine, M.; Fox, S.; French, R.A.; Blouin, M.S. A single generation of domestication heritably alters the expression of hundreds of genes. Nat. Commun. 2016, 7, 10676. [Google Scholar] [CrossRef]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, A.; Lindblad-Toh, K. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef]
- Carneiro, M.; Rubin, C.J.; Di Palma, F.; Albert, F.W.; Alföldi, J.; Barrio, A.M.; Pielberg, G.; Rafati, N.; Sayyab, S.; Turner-Maier, J.; et al. Rabbit genome analysis reveals a polygenic basis for phenotypic change during domestication. Science 2014, 345, 1074–1079. [Google Scholar] [CrossRef]
- Montague, M.J.; Li, G.; Gandolfi, B.; Khan, R.; Aken, B.L.; Searle, S.M.J.; Minx, P.; Hillier, L.W.; Koboldt, D.C.; Davis, B.W.; et al. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 17230–17235. [Google Scholar] [CrossRef]
- Mäkinen, H.; Vasemägi, A.; McGinnity, P.; Cross, T.F.; Primmer, C.R. Population genomic analyses of early-phase Atlantic Salmon (Salmo salar) domestication/captive breeding. Evol. Appl. 2015, 8, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Nunney, L. The cost of natural selection revisited. Ann. Zool. Fenn. 2003, 40, 185–194. [Google Scholar]
- Roberge, C.; Normandeau, É.; Einum, S.; Guderley, H.; Bernatchez, L. Genetic consequences of interbreeding between farmed and wild Atlantic salmon: Insights from the transcriptome. Mol. Ecol. 2008, 17, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.R.; Ford, M.J.; Blouin, M.S. On the reproductive success of early-generation hatchery fish in the wild. Evol. Appl. 2014, 7, 883–896. [Google Scholar] [CrossRef]
- Floridi, L. The Logic of Information. A Theory of Philosophy as Conceptual Design; Oxford University Press: Oxford, UK, 2019; p. 240. [Google Scholar]
- Khun, T.S. The Structure of Scientific Revolutions; University of Chicago Press: Chicago, IL, USA, 1962; p. 264. [Google Scholar]
- Geffroy, B.; Sadoul, B.; Putman, B.J.; Berger-Tal, O.; Garamszegi, L.Z.; Møller, A.P.; Blumstein, D.T. Evolutionary dynamics in the Anthropocene: Life history and intensity of human contac shape antipredator responses. PLoS Biol. 2020, 18, e3000818. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/3/i2414e/i2414e05.pdf (accessed on 2 March 2021).
- Leonard, S.M.; Xin, H.; Brown-Brandl, T.M.; Ramirez, B.C.; Dutta, S.; Rohrer, G.A. Effects of Farrowing Stall Layout and Number of Heat Lamps on Sow and Piglet Production Performance. Animals 2020, 10, 348. [Google Scholar] [CrossRef]
- Rollin, B. The Frankenstein Syndrome: Ethical and Social Issues in the Genetic Engineering of Animals; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1995; p. 245. [Google Scholar]
- Shriver, A. Prioritizing the protection of welfare in gene-edited livestock. Anim. Front. 2020, 10, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.P.; White, I.M.S.; Brotherstone, S.; Farnworth, M.J.; Knap, P.W.; Penny, P.; Mendl, M.; Lawrence, A.B. Heritability of post-mixing aggressiveness in grower-stage pigs and its relationship with production traits. Anim. Sci. 2006, 82, 615–620. [Google Scholar] [CrossRef]
- Meffert, L.M.; Hicks, S.K.; Regan, J.L. Nonadditive genetic effects in animal behavior. Am. Nat. 2002, 160 (Suppl. 6), S198–S213. [Google Scholar] [CrossRef]
- Angvall, B.; Jöngren, M.; Strandberg, E.; Jensen, P. Heritability and genetic correlations of fear-related behaviour in red junglefowl—possible implications for early domestication. PLoS ONE 2012, 7, e35162. [Google Scholar] [CrossRef]
- Kjaer, J.B.; Sørensen, P. Feather pecking behaviour in White Leghorn chickens, a genetic study. Br. Poult. Sci. 1997, 38, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Riber, A.B.; Hinrichsen, L.K. Welfare consequences of omitting beak trimming in barn layers. Front. Vet. Sci. 2017, 4, 222. [Google Scholar] [CrossRef]
- Ding, Y.; Berrocal, A.; Morita, T.; Longden, K.D.; Stern, D.L. Natural courtship song variation caused by an intronic retroelement in an ion channel gene. Nature 2016, 536, 329–332. [Google Scholar] [CrossRef]
- Boissy, A.; Bouix, J.; Orgeur, P.; Poindron, P.; Bibé, B.; Le Neindre, P. Genetic analysis of emotional reactivity in sheep: Effects of the genotypes of the lambs and of their dams. Genet. Sel. Evol. 2005, 37, 381–401. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.P.; Roehe, R.; Mekkawy, W.; Farnworth, M.J.; Knap, P.W.; Lawrence, A.B. Bayesian analysis of genetic associations of skin lesions and behavioural traits to identify genetic components of individual aggressiveness in pigs. Behav. Genet. 2008, 38, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Drews, C. The concept and definition of dominance in animal behaviour. Behaviour 1993, 125, 283–313. [Google Scholar] [CrossRef]
- Phillips, C.J.C. Cattle Behaviour; Farming Press Books: Ipswich, UK, 1993. [Google Scholar]
- Knap, P.W.; Merks, J.W.M. A note on the genetics of aggressiveness of primiparous sows towards their piglets. Livest. Prod. Sci. 1987, 17, 161–167. [Google Scholar] [CrossRef]
- Hofmeyr, I. The fighting bulls of Portugal: Genetics & handling. Stockfarm 2016, 6, 34–35. [Google Scholar]
- Ulloa-Arvizu, R.; Gayosso-Vázquez, A.; Ramos-Kuri, M.; Estrada, F.J.; Montaño, M.; Alonso, R. A Genetic analysis of Mexican Criollo cattle populations. J. Anim. Breed. Genet. 2008, 125, 351–359. [Google Scholar] [CrossRef]
- Buchanan, D.S.; Lenstra, J.A. Breeds of cattle. In The Genetics of Cattle, 2nd ed.; Garrick, D.J., Ruvinsky, A., Eds.; CABI Publishers: Wallingford, UK, 2015; p. 33. [Google Scholar]
- Sartori, C.; Mantovani, R. Effects of inbreeding on fighting ability measured in Aosta Chestnut and Aosta Black Pied cattle. J. Anim. Sci. 2012, 90, 2907–2915. [Google Scholar] [CrossRef]
- Sartori, C.; Manser, M.B.; Mantovani, R. Relationship between number and intensity of fighting: Evidence from cow fighting tournaments in Valdostana cattle. Ital. J. Anim. Sci. 2014, 13, 2907–2915. [Google Scholar] [CrossRef]
- Sartori, C.; Guzzo, N.; Mantovani, R. Genetic correlations of fighting ability with somatic cells and longevity in cattle. Animal 2020, 14, 13–21. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Ben Jemaa, S.; Ciani, E.; Sottile, G.; Moscarelli, A.; Boussaha, M.; Montedoro, M.; Pilia, F.; Cassandro, M. Genome-wide detection of signatures of selection in three Valdostana cattle populations. J. Anim. Breed. Genet. 2020, 137, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Misztal, I. Breeding and genetics symposium: Resilience and lessons from studies in genetics of heat stress. J. Anim. Sci. 2017, 95, 1780–1787. [Google Scholar] [CrossRef] [PubMed]
- Blasco, A. Animal breeding methods and sustainability. Am. Nat. 2012, 139, 749–770. [Google Scholar]
- Frankham, R. Genetic adaptation to captivity in species conservation programs. Mol. Ecol. 2008, 17, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Crowell-Davis, S.L. Behavior problems in pet rabbits. J. Exot. Pet Med. 2007, 16, 38–44. [Google Scholar] [CrossRef]
- Scott, J.P.; Fuller, J.L. Genetics and the Social Behavior of the Dog; University of Chicago Press: Chicago, IL, USA, 2012; Volume 570. [Google Scholar]
- Ilska, J.; Haskell, M.J.; Blott, S.C.; Sánchez-Molano, E.; Polgar, Z.; Lofgren, S.E.; Clements, D.N.; Wiener, P. Genetic characterization of dog personality traits. Genetics 2017, 206, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- McDonnel, S. The Equid Ethogram: A Practical Field Guide to Horse Behaviour; The Blood Horse Inc.: Lexington, KY, USA, 2003. [Google Scholar]
- Deesing, M.J.; Grandin, T. Behavior Genetics of the Horse (Equus caballus). In Genetics and the Behavior of Domestic Animals, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 237–290. [Google Scholar]
- Pulina, G.; Francesconi, A.H.D.; Stefanon, B.; Sevi, A.; Calamari, L.; Lacetera, N.; Dell’Orto, V.; Pilla, F.; Marsan, P.A.; Mele, M.; et al. Sustainableruminant production to help feed the planet. Ital. J. Anim. Sci. 2017, 16, 140–171. [Google Scholar] [CrossRef]
- Pulina, G.; Tondo, A.; Danieli, P.P.; Primi, R.; Crovetto, G.M.; Fantini, A.; Macciotta, N.P.P.; Atzori, A.S. How to manage cows yielding 20,000 kg of milk: Technical challenges and environmental implications. Ital. J. Anim. Sci. 2020, 19, 865–879. [Google Scholar] [CrossRef]
- EU Commission, 2016. Commission recommendation (EU) 2016/336 of 8 March 2016 on the application of Council Directive 2008/120/EC laying down minimum standards for the protection of pigs as regards measures to reduce the need for tail-docking. OJEU 9.3 2016, 62, 20–22. [Google Scholar]
- Van de Weerd, H.; Ison, S. Providing Effective Environmental Enrichment to Pigs: How Far Have We Come? Animals 2019, 9, 254. [Google Scholar] [CrossRef] [PubMed]
- Norton, T.; Chen, C.; Larsen, M.L.V.; Berckmans, D. Review: Precision livestock farming: Building ‘digital representations’ to bring the animals closer to the farmer. Animal 2019, 13, 3009–3017. [Google Scholar] [CrossRef] [PubMed]
- Neethirajan, S. The role of sensors, big data and machine learning in modern animal farming. Sens. Bio-Sens. Res. 2020, 29. [Google Scholar] [CrossRef]
- Word Organization for Animal Health. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahc/current/chapitre_aw_introduction.pdf (accessed on 2 March 2021).


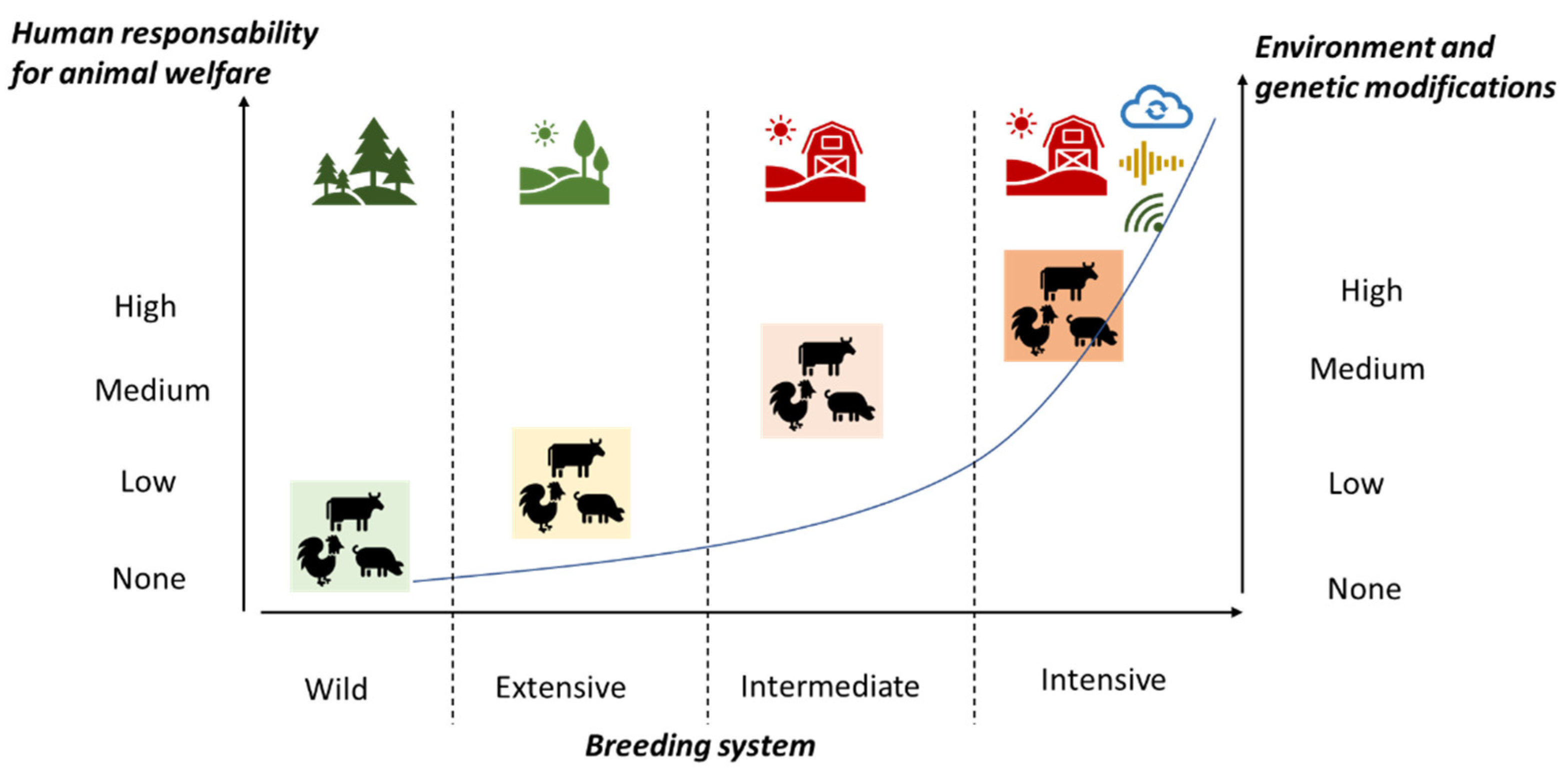{kind=link}
| Trait | Livestock Species |
|---|---|
| Curly tails | Dog, pig |
| Depigmentation | Cattle, dog, goat, horse, pig, rabbit |
| Docility | Cattle, dog, donkey, goat, horse, pig, rabbit, sheep |
| Floppy ears | Cattle, donkey, dog, rabbit |
| More frequent estrous cycles | Dog, goat |
| Neotenous (juvenile) behavior | Dog |
| Reduced ears | Dog |
| Shorter muzzles | Cattle, dog, goat, pig, sheep |
| Smaller brain or cranial capacity | Cattle, dog, goat, horse, pig, rabbit |
| Smaller teeth | Dog, pig |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cesarani, A.; Pulina, G. Farm Animals Are Long Away from Natural Behavior: Open Questions and Operative Consequences on Animal Welfare. Animals 2021, 11, 724. https://doi.org/10.3390/ani11030724
Cesarani A, Pulina G. Farm Animals Are Long Away from Natural Behavior: Open Questions and Operative Consequences on Animal Welfare. Animals. 2021; 11(3):724. https://doi.org/10.3390/ani11030724
Chicago/Turabian StyleCesarani, Alberto, and Giuseppe Pulina. 2021. "Farm Animals Are Long Away from Natural Behavior: Open Questions and Operative Consequences on Animal Welfare" Animals 11, no. 3: 724. https://doi.org/10.3390/ani11030724
APA StyleCesarani, A., & Pulina, G. (2021). Farm Animals Are Long Away from Natural Behavior: Open Questions and Operative Consequences on Animal Welfare. Animals, 11(3), 724. https://doi.org/10.3390/ani11030724