
Effects of Different Parts on the Chemical Composition, Silage Fermentation Profile, In Vitro and In Situ Digestibility of Paper Mulberry

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Harvesting and Different Fractions of the Plants
2.2. Nutritional Composition of Fresh Samples
2.3. PM Silage
2.3.1. Silage Making
2.3.2. Silage Chemical Composition Determination
2.4. In Vitro Digestibility
2.5. In Situ Digestibility
2.6. Calculation and Statistical Analysis
2.6.1. Calculation of Gas Production Parameters for In Vitro Fermentation
2.6.2. Calculation of in Situ Digestibility
2.6.3. Statistical Analysis
3. Results
3.1. Yield and Chemical Composition
3.2. Silage Fermentation Profile
3.3. In Vitro Digestibility
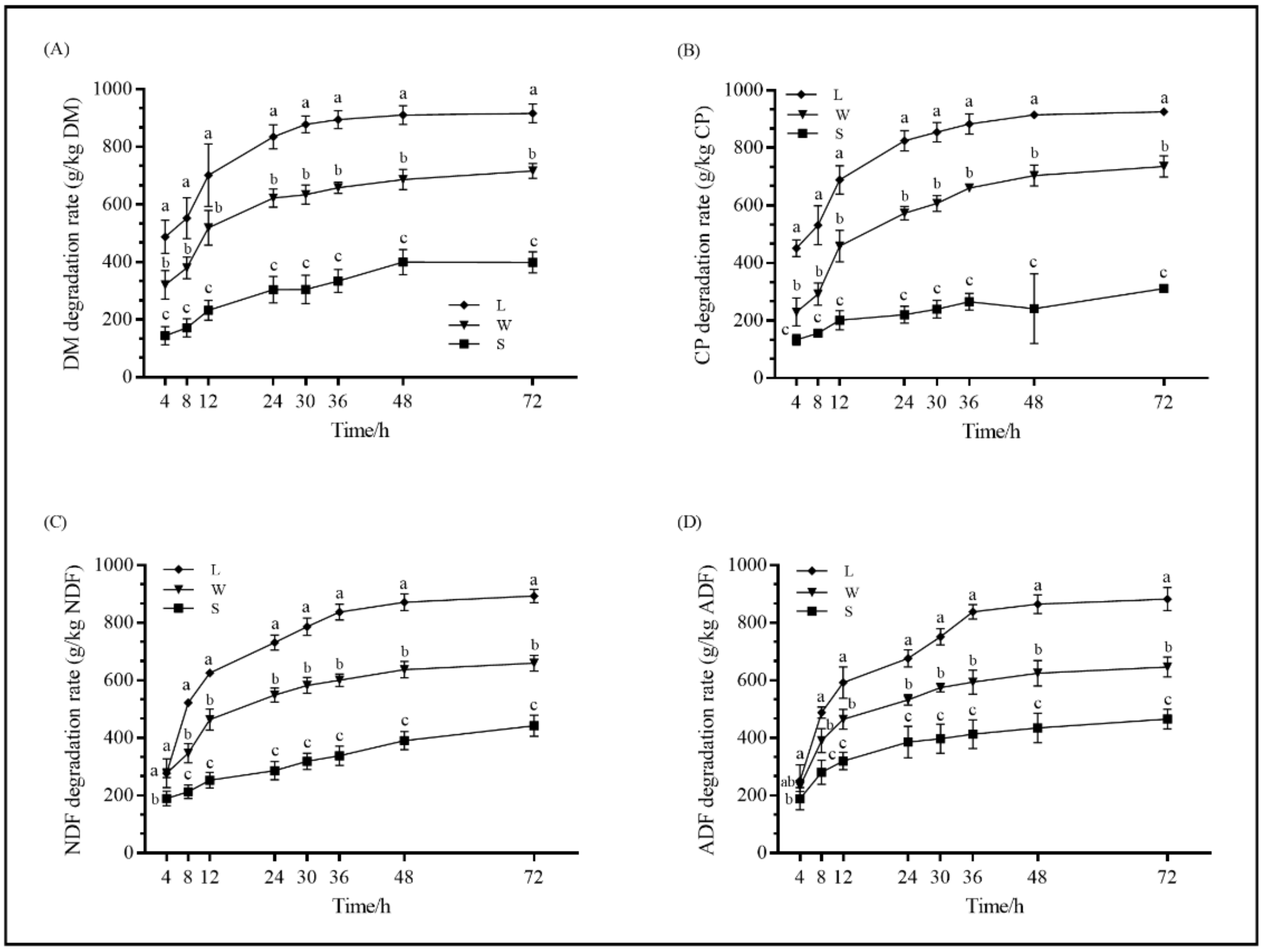
3.4. In Situ Digestibility
4. Discussion
4.1. Yield and Chemical Composition
4.2. Silage Fermentation Profile
4.3. Digestibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penailillo, J.; Olivares, G.; Moncada, X.; Payacan, C.; Chang, C.S.; Chung, K.F.; Matthews, P.J.; Seelenfreund, A.; Seelenfreund, D. Sex Distribution of Paper Mulberry (Broussonetia papyrifera) in the Pacific. PLoS ONE 2016, 11, e0161148. [Google Scholar] [CrossRef]
- Xianjun, P.; Yucheng, W.; Ruiping, H.; Meiling, Z.; Shihua, S. Global transcriptomics identification and analysis of transcriptional factors in different tissues of the paper mulberry. BMC Plant Biol. 2014, 14, 194. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, Z.F.; Luo, X.; Li, X. Effects of mulberry fruit (Morus alba L.) consumption on health outcomes: A mini-review. Antioxidants 2018, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.J.; Park, S. Mulberry extract supplements ameliorate the inflammation-related hematological parameters in carrageenan-induced arthritic rats. J. Med. Food. 2006, 9, 431–435. [Google Scholar] [CrossRef]
- Du, Q.; Zheng, J.; Xu, Y. Composition of anthocyanins in mulberry and their antioxidant activity. J. Food Compost. Anal. 2008, 21, 390–395. [Google Scholar] [CrossRef]
- Tang, F.; Chen, N.; Zhao, M.; Wang, Y.; He, R.; Peng, X.; Shen, S. Identification and functional divergence analysis of WOX gene family in paper mulberry. Int. J. Mol. Sci. 2017, 18, 1782. [Google Scholar] [CrossRef]
- Ouyang, J.; Wang, M.; Hou, Q.; Feng, D.; Pi, Y.; Zhao, W. Effects of dietary mulberry leaf powder in concentrate on the rumen fermentation and ruminal epithelium in fattening Hu sheep. Animals 2019, 9, 218. [Google Scholar] [CrossRef]
- Si, B.; Tao, H.; Zhang, X.; Guo, J.; Cui, K.; Tu, Y.; Diao, Q. Effect of Broussonetia papyrifera L. (paper mulberry) silage on dry matter intake, milk composition, antioxidant capacity and milk fatty acid profile in dairy cows. Asian-Australas J. Anim. Sci. 2018, 31, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Bruschi, S.; Fortunati, P.; Masoero, F. Use of principal factor analysis to generate a corn silage fermentative quality index to rank well- or poorly preserved forages. J. Sci. Food Agric. 2016, 96, 1686–1696. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Olof, T.; Åman, P.; Eric, W.; Roger, A.; Dan, P. Total dietary fiber determined as neutral sugar residues, uronic acid residues, and Klason lignin (the Uppsala method): Collaborative study. J. AOAC Int. 1995, 78, 1030–1044. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Murphy, R.P. A method for the extraction of plant samples and the determination of total soluble carbohydrates. J. Sci. Food Agric. 1958, 9, 714–717. [Google Scholar] [CrossRef]
- Playne, M.J.; Mcdonald, P. Buffering constituents of herbage and of silage. J. Sci. Food Agric. 1966, 17, 264–268. [Google Scholar] [CrossRef]
- Li, R.; Hou, G.; Jiang, X.; Song, Z.; Fan, Z.; Hou, D.X.; He, X. Different dietary protein sources in low protein diets regulate colonic microbiota and barrier function in a piglet model. Food Funct. 2019, 10, 6417–6428. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Sun, Q.; Li, F.; Xu, C.; Cai, Y. Comparative analysis of ensiling characteristics and protein degradation of alfalfa silage prepared with corn or sweet sorghum in semiarid region of Inner Mongolia. Anim. Sci. J. 2020, 91, e13321. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.F.; Kang, J.H.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 46–75. [Google Scholar] [CrossRef]
- Yuan, X.J.; Wen, A.Y.; Wang, J.; Desta, S.T.; Dong, Z.H.; Shao, T. Effects of four short-chain fatty acids or salts on fermentation characteristics and aerobic stability of alfalfa (Medicago sativa L.) silage. J. Sci. Food Agric. 2018, 98, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Cao, Z.J.; Cao, B.B.; Yang, H.J.; Li, S.L.; Liu, J.X. Effects of different forage combinations in total mixed rations on in vitro gas production kinetics, ruminal and milk fatty acid profiles of lactating cows. Anim. Sci. J. 2018, 89, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H.; Stengass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Ryan, J.P. Determination of volatile fatty acids and some related compounds in ovine rumen fluid, urine, and blood plasma, by gas-liquid chromatography. Anal. Biochem. 1980, 108, 374–384. [Google Scholar] [CrossRef]
- Nocek, J.E. In situ and other methods to estimate ruminal protein and energy digestibility: A review. J. Dairy Sci. 1988, 91, 2051–2069. [Google Scholar] [CrossRef]
- Groot, J.C.J.; Cone, J.W.W.; Illiams, B.A.; Debersaques, F.M.A.; Lantinga, E.A. Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- Yang, H.J.; Tamminga, S.; Williams, B.A.; Dijkstra, J.; Boer, H. In vitro gas and volatile fatty acids production profiles of barley and maize and their soluble and washout fractions after feed processing. Anim. Feed Sci. Technol. 2005, 120, 125–140. [Google Scholar] [CrossRef]
- Orskov, E.R.; Mcdonald, I. Estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef]
- Batajoo, K.K.; Shaver, R.D. Impact of nonfiber carbohydrate on intake, digestion, and milk production by dairy cows. J. Dairy Sci. 1994, 77, 1580–1588. [Google Scholar] [CrossRef]
- Hall, M.B.; Mertens, D.R. A 100-year review: Carbohydrates-characterization, digestion, and utilization. J. Dairy Sci. 2017, 100, 10078–10093. [Google Scholar] [CrossRef]
- Grant, R.J.; Ferraretto, L.F. Silage review: Silage feeding management: Silage characteristics and dairy cow feeding behavior. J. Dairy Sci. 2018, 101, 4111–4121. [Google Scholar] [CrossRef]
- Kanjanapruthipong, J.; Junlapho, W.; Karnjanasirm, K. Feeding and lying behavior of heat-stressed early lactation cows fed low fiber diets containing roughage and nonforage fiber sources. J. Dairy Sci. 2015, 98, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Schulze, A.K.; Weisbjerg, M.R.; Norgaard, P. Effects of feeding level and NDF content of grass-clover silages on chewing activity, fecal particle size and NDF digestibility in dairy heifers. Animal 2014, 8, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Hall, M.B.; Gallo-Meagher, M.; Smith, R.L. Improvement of forage quality by downregulation of maize o-methyltransferase. Crop. Sci. 2003, 43, 2240–2251. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Tang, W.; Wu, X.; Huang, C.; Huang, C.; Lai, C.; Yong, Q. Enhancing enzymatic digestibility of waste wheat straw by presoaking to reduce the ash-influencing effect on autohydrolysis. Biotechnol. Biofuels 2019, 12, 222. [Google Scholar] [CrossRef]
- Dong, X.; Zhou, Z.; Saremi, B.; Helmbrecht, A.; Wang, Z.; Loor, J.J. Varying the ratio of Lys:Met while maintaining the ratios of Thr:Phe, Lys:Thr, Lys:His, and Lys:Val alters mammary cellular metabolites, mammalian target of rapamycin signaling, and gene transcription. J. Dairy Sci. 2018, 101, 1708–1718. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.S.; Ji, P.; Drackley, J.K.; Luchini, D.; Loor, J.J. Supplemental smartamine M or metaSmart during the transition period benefits postpartal cow performance and blood neutrophil function. J. Dairy Sci. 2013, 96, 6248–6263. [Google Scholar] [CrossRef] [PubMed]
- Giallongo, F.; Hristov, A.N.; Oh, J.; Frederick, T.; Weeks, H.; Werner, J.; Lapierre, H.; Patton, R.A.; Gehman, A.; Parys, C. Effects of slow-release urea and rumen-protected methionine and histidine on performance of dairy cows. J. Dairy Sci. 2015, 98, 3292–3308. [Google Scholar] [CrossRef]
- Lee, C.; Hristov, A.N.; Cassidy, T.W.; Heyler, K.S.; Lapierre, H.; Varga, G.A.; de Veth, M.J.; Patton, R.A.; Parys, C. Rumen-protected lysine, methionine, and histidine increase milk protein yield in dairy cows fed a metabolizable protein-deficient diet. J. Dairy Sci. 2012, 95, 6042–6056. [Google Scholar] [CrossRef]
- Giallongo, F.; Harper, M.T.; Oh, J.; Lopes, J.C.; Lapierre, H.; Patton, R.A.; Parys, C.; Shinzato, I.; Hristov, A.N. Effects of rumen-protected methionine, lysine, and histidine on lactation performance of dairy cows. J. Dairy Sci. 2016, 99, 4437–4452. [Google Scholar] [CrossRef]
- Van Amburgh, M.E.; Collao-Saenz, E.A.; Higgs, R.J.; Ross, D.A.; Recktenwald, E.B.; Raffrenato, E.; Chase, L.E.; Overton, T.R.; Mills, J.K.; Foskolos, A. The Cornell Net Carbohydrate and Protein System: Updates to the model and evaluation of version 6.5. J. Dairy Sci. 2015, 98, 6361–6380. [Google Scholar] [CrossRef]
- Fleming, A.J.; Lapierre, H.; White, R.R.; Tran, H.; Kononoff, P.J.; Martineau, R.; Weiss, W.P.; Hanigan, M.D. Predictions of ruminal outflow of essential amino acids in dairy cattle. J. Dairy Sci. 2019, 102, 10947–10963. [Google Scholar] [CrossRef]
- Silva, V.P.; Pereira, O.G.; Leandro, E.S.; da Silva, T.C.; Ribeiro, K.G.; Mantovani, H.C.; Santos, S.A. Effects of lactic acid bacteria with bacteriocinogenic potential on the fermentation profile and chemical composition of alfalfa silage in tropical conditions. J. Dairy Sci. 2016, 99, 1895–1902. [Google Scholar] [CrossRef] [PubMed]
- Winters, A.L.; Cockburn, J.E.; Dhanoa, M.S.; Merry, R.J. Effects of lactic acid bacteria in inoculants on changes in amino acid composition during ensilage of sterile and non-sterile ryegrass. J. Appl. Microbiol. 2000, 89, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Mahanna, B. Proper management assures high-quality silage, grains. Feedstuffs 1994, 66, 17–20. [Google Scholar]
- Li, Q.; Xue, B.; Zhao, Y.; Wu, T.; Liu, H.; Yi, X.; Sun, C.; Wang, Z.; Zou, H.; Yan, T. In situ degradation kinetics of 6 roughages and the intestinal digestibility of the rumen undegradable protein. J. Anim. Sci. 2018, 96, 4835–4844. [Google Scholar] [CrossRef]
- Krizsan, S.J.; Nyholm, L.; Nousiainen, J.; Südekum, K.-H.; Huhtanen, P. Comparison of in vitro and in situ methods in evaluation of forage digestibility in ruminants1. J. Anim. Sci. 2012, 90, 3162–3173. [Google Scholar] [CrossRef]
- Hao, Y.; Huang, S.; Si, J.; Zhang, J.; Gaowa, N.; Sun, X.; Lv, J.; Liu, G.; He, Y.; Wang, W.; et al. Effects of paper mulberry silage on the milk production, apparent digestibility, antioxidant capacity, and fecal bacteria composition in Holstein dairy cows. Animals 2020, 10, 1152. [Google Scholar] [CrossRef]
- Getachew, G.; Robinson, P.H.; DePeters, E.J.; Taylor, S.J. Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Anim. Feed Sci. Technol. 2004, 111, 57–71. [Google Scholar] [CrossRef]
- Kamalak, A.; Canbolat, O.; Gurbuz, Y.; Ozay, O. Comparison of in vitro gas production technique with in situ nylon bag technique to estimate dry matter degradation. Czech. J. Anim. Sci. 2011, 50, 60–67. [Google Scholar] [CrossRef]
- Grant, R.J. Influence of corn and sorghum starch on the in vitro kinetics of forage fiber digestion. J. Dairy Sci. 1994, 77, 1563–1569. [Google Scholar] [CrossRef]
- Hess, H.D.; Monsalve, L.M.; Lascano, C.E.; Carulla, J.E.; Díaz, T.E.; Kreuzer, M. Supplementation of a tropical grass diet with forage legumes and Sapindus saponaria fruits: Effects on in vitro ruminal nitrogen turnover and methanogenesis. Aust. J. Agric. Res. 2003, 54, 703–713. [Google Scholar] [CrossRef]
- Jaworski, N.W.; Lrke, H.N.; Bach Knudsen, K.E.; Stein, H.H. Carbohydrate composition and in vitro digestibility of dry matter and nonstarch polysaccharides in corn, sorghum, and wheat and coproducts from these grains. J. Anim. Sci. 2015, 93, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Rinker, M.; McLeod, K.R.; Harmon, D.L. Yucca schidigera extract decreases in vitro methane production in a variety of forages and diets. Anim. Feed Sci. Technol. 2010, 159, 18–26. [Google Scholar] [CrossRef]
- Getachew, G.; Laca, E.A.; Putnam, D.H.; Witte, D.; McCaslin, M.; Ortega, K.P.; DePeters, E.J. The impact of lignin downregulation on alfalfa yield, chemical composition, and in vitro gas production. J. Sci. Food Agric. 2018, 98, 4205–4215. [Google Scholar] [CrossRef]
- Beuvink, J.M.W.; Spoelstra, S.F. Interactions between substrate, fermentation end-products, buffering systems and gas production upon fermentation of different carbohydrates by mixed rumen microorganisms in vitro. Appl. Microbiol. Biotechnol. 1992, 37, 505–509. [Google Scholar] [CrossRef]
- Getachew, G.; Makkar, H.; Becker, K. Tropical browses: Contents of phenolic compounds, in vitro gas production and stoichiometric relationship between short chain fatty acid and in vitro gas production. J. Agric. Sci. 2002, 139, 341–352. [Google Scholar] [CrossRef]


{kind=link}
| Items 1 | Groups | ||
|---|---|---|---|
| Leaf | Stem | Whole Plant | |
| FW yield (kg/per plant) | 1.70 ± 0.05 b | 1.17 ± 0.04 c | 2.79 ± 0.09 a |
| DM (g/kg) | 261.05 ± 4.00 | 271.06 ± 8.06 | 266 ± 6.07 |
| CP (g/kg DM) | 241.24 ± 0.98 a | 84.93 ± 3.29 c | 169.40 ± 4.23 b |
| NDF (g/kg DM) | 447.09 ± 31.61 c | 680.63 ± 14.60 a | 502.79 ± 11.07 b |
| ADF (g/kg DM) | 239.00 ± 13.86 c | 565.91 ± 9.81 a | 348.88 ± 5.57 b |
| Lignin (g/kg DM) | 33.45 ± 5.79 c | 75.67 ± 2.12 a | 59.61 ± 3.11 b |
| EE (g/kg DM) | 29.24 ± 0.04 a | 22.82 ± 0.25 b | 22.54 ± 2.88 b |
| WSC (g/kg DM) | 45.82 ± 4.62 a | 11.57 ± 0.77 c | 33.24 ± 1.32 b |
| Ash (g/kg DM) | 116.62 ± 0.73 a | 47.31 ± 0.41 c | 84.95 ± 5.38 b |
| Calcium (g/kg DM) | 26.47 ± 1.79 a | 9.09 ± 1.07 c | 15.55 ± 0.92 b |
| Phosphorus (g/kg DM) | 3.01 ± 0.11 a | 2.50 ± 0.23 b | 2.26 ± 0.28 b |
| Buffer capacity (mE/Kg · DM) | 418.30 ± 21.25 a | 187.97 ± 5.51 c | 297.94 ± 26.86 b |
| Items | Groups | ||
|---|---|---|---|
| Leaf | Stem | Whole Plant | |
| Animo acid (g/kg DM) | |||
| Tryptophan | 3.74 ± 0.04 a | 0.83 ± 0.04 c | 2.41 ± 0.02 b |
| Cysteine | 2.21 ± 0.05 a | 0.51 ± 0.02 c | 1.63 ± 0.07 b |
| Methionine | 3.92 ± 0.15 a | 1.04 ± 0.04 c | 2.53 ± 0.23 b |
| Aspartic acid | 23.42 ± 0.71 a | 9.83 ± 0.20 c | 17.81 ± 0.43 b |
| Threonine | 11.12 ± 0.12 a | 2.74 ± 0.05c | 7.23 ± 0.11 b |
| Serine | 9.73 ± 0.23 a | 2.84 ± 0.05 c | 6.89 ± 0.17 b |
| Glutamic acid | 24.68 ± 0.28 a | 6.33 ± 0.10 c | 16.53 ± 0.49 b |
| Proline | 15.72 ± 0.37a | 5.30 ± 0.17 c | 10.63 ± 0.62 b |
| Glycine | 11.32 ± 0.18 a | 2.58 ± 0.07 c | 7.59 ± 0.20 b |
| Alanine | 10.29 ± 0.20 a | 2.54 ± 0.04 c | 7.04 ± 0.18 b |
| Valine | 13.41 ± 0.38 a | 3.46 ± 0.04 c | 9.03 ± 0.23 b |
| Isoleucine | 10.40 ± 0.21 a | 2.45 ± 0.11 c | 7.03 ± 0.18 b |
| Leucine | 18.82 ± 0.22 a | 4.21 ± 0.15 c | 12.52 ± 0.24 b |
| Tyrosine | 7.48 ± 0.18 a | 0.91 ± 0.02 c | 4.47 ± 0.17 b |
| Phenylalanine | 11.67 ± 0.32 a | 2.58 ± 0.10 c | 7.66 ± 0.08 b |
| Histidine | 4.51 ± 0.08 a | 1.26 ± 0.03 c | 3.17 ± 0.12 b |
| Lysine | 13.53 ± 0.28 a | 3.48 ± 0.10 c | 9.68 ± 0.24 b |
| Arginine | 12.81 ± 0.22 a | 2.43 ± 0.04 c | 8.27 ± 0.10 b |
| Items 1 | Groups | ||
|---|---|---|---|
| Leaf | Stem | Whole Plant | |
| Chemical Composition | |||
| DM (g/kg) | 261.04 ± 4.00 ab | 271.06 ± 8.06 a | 256.80 ± 1.70 b |
| CP (g/kg DM) | 252.87 ± 11.57 a | 92.28 ± 1.90 c | 190.76 ± 6.26 b |
| NDF (g/kg DM) | 304.25 ± 6.06 c | 670.19 ± 2.68 a | 415.45 ± 12.91 b |
| ADF (g/kg DM) | 158.48 ± 14.29 c | 580.47 ± 8.57 a | 323.72 ± 23.90 b |
| Fermentation Profile | |||
| pH | 5.09 ± 0.06 a | 3.81 ± 0.01 c | 4.80 ± 0.20 b |
| NH3-N (g/kg TN) | 44.07 ± 7.37 a | 20.91 ± 3.23 b | 43.94 ± 10.87 a |
| Lactate (g/kg DM) | 99.77 ± 2.29 a | 52.07 ± 6.62 b | 90.65 ± 13.92 a |
| Acetate (g/kg DM) | 39.63 ± 4.71 a | 6.43 ± 1.78 c | 22.71 ± 2.52 b |
| Propionate (g/kg DM) | 21.03 ± 2.27 a | ND b | 23.96 ± 6.03 a |
| Butyrate (g/kg DM) | ND | ND | ND |
| Items 1 | Groups | ||
|---|---|---|---|
| Leaf | Stem | Whole Plant | |
| IVDMD (g/kg DM) | 909.00 ± 22.73 a | 484.31 ± 22.16 c | 683.51 ± 8.6 b |
| pH | 6.94 ± 0.02 | 6.93 ± 0.07 | 6.97 ± 0.05 |
| NH3-N (mg/dL) | 12.08 ± 0.45 | 11.10 ± 0.83 | 11.94 ± 0.30 |
| Acetate (mmol/L) | 53.03 ± 0.59 a | 48.25 ± 1.79 b | 52.99 ± 1.59 a |
| Propionate (mmol/L) | 24.67 ± 1.04 a | 19.09 ± 0.98 b | 20.07 ± 0.87 b |
| Isobutyrate (mmol/L) | 0.72 ± 0.17 b | 1.57 ± 0.08 a | 1.67 ± 0.15 a |
| Butyrate (mmol/L) | 9.76 ± 0.76 | 8.32 ± 0.39 | 9.28 ± 0.52 |
| Isovalerate (mmol/L) | 2.39 ± 0.28 | 2.04 ± 0.15 | 2.44 ± 0.21 |
| Valerate (mmol/L) | 2.05 ± 0.07 a | 1.65 ± 0.01 b | 1.92 ± 0.10 ab |
| Total VFA (mmol/L) | 92.63 ± 2.83 a | 80.92 ± 1.36 b | 88.37 ± 1.89ab |
| GP48 (mL/g) | 90.05 ± 7.74 | 74.24 ± 12.28 | 82.40 ± 8.31 |
| A | 95.65 ± 7.72 a | 75.92 ± 12.53 b | 89.56 ± 8.50 b |
| B | 1.44 ± 0.10 a | 1.28 ± 0.04 b | 1.27 ± 0.05 b |
| C | 3.95 ± 0.33 | 4.00 ± 0.57 | 4.09 ± 0.60 |
| TRmaxG | 1.21 ± 0.31 | 0.78 ± 0.09 | 0.78 ± 0.25 |
| RmaxG | 15.05 ± 2.10 | 12.28 ± 1.95 | 13.38 ± 2.60 |
| TRmaxS | 2.07 ± 0.54 | 1.56 ± 0.02 | 1.60 ± 0.48 |
| RmaxS | 0.20 ± 0.02 | 0.19 ± 0.03 | 0.19 ± 0.03 |
| Items 1 | Groups | ||
|---|---|---|---|
| Leaf | Stem | Whole Plant | |
| DM | |||
| a (g/kg DM) | 337.91 ± 65.14 a | 104.96 ± 7.63 b | 147.74 ± 39.18 b |
| b (g/kg DM) | 601.13 ± 43.94 a | 345.70 ± 37.62 b | 544.90 ± 29.24 a |
| c (g/kg DM) | 63.09 ± 11.86 a | 30.71 ± 4.63 b | 78.72 ± 4.98 a |
| a + b (g/kg DM) | 939.04 ± 23.73 a | 450.66 ± 34.85 c | 702.64 ± 15.41 b |
| ED (g/kg DM) | 621.96 ± 19.15 a | 208.92 ± 6.12 c | 439.59 ± 27.43 b |
| CP | |||
| a (g/kg DM) | 306.21 ± 41.64 a | 155.68 ± 16.15 b | 174.71 ± 5.73 b |
| b (g/kg DM) | 626.80 ± 32.98 a | 293.78 ± 99.84 b | 629.84 ± 29.96 a |
| c (g/kg DM) | 89.14 ± 1.97 a | 50.47 ± 7.34 b | 59.41 ± 17.28 b |
| a + b (g/kg DM) | 933.01 ± 15.72 a | 409.45 ± 116.09 b | 804.56 ± 30.77 a |
| ED (g/kg DM) | 612.85 ± 23.25 a | 191.74 ± 10.65 c | 455.05 ± 42.93 b |
| NDF | |||
| a (g/kg DM) | 87.82 ± 15.49 c | 162.08 ± 15.59 b | 214.19 ± 23.66 a |
| b (g/kg DM) | 788.51 ± 11.68 a | 414.21 ± 11.61 b | 458.06 ± 23.19 b |
| c (g/kg DM) | 87.76 ± 4.07 a | 20.56 ± 8.61 c | 54.43 ± 7.84 b |
| a + b (g/kg DM) | 876.33 ± 7.00 a | 576.29 ± 124.42 b | 672.25 ± 44.54 b |
| ED (g/kg DM) | 526.38 ± 3.03 a | 244.92 ± 10.83 c | 412.93 ± 18.68 b |
| ADF | |||
| a (g/kg DM) | 134.26 ± 4.39 | 137.02 ± 22.41 | 102.44 ± 18.32 |
| b (g/kg DM) | 736.82 ± 35.47 a | 315.26 ± 49.50 c | 524.71 ± 12.98 b |
| c (g/kg DM) | 69.08 ± 5.28 | 58.41 ± 17.13 | 79.41 ± 2.79 |
| a + b (g/kg DM) | 871.09 ± 14.80 a | 452.27 ± 29.12 c | 627.15 ± 10.90 b |
| ED (g/kg DM) | 500.23 ± 14.13 a | 280.17 ± 20.50 c | 381.29 ± 14.32 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Y.; Huang, S.; Liu, G.; Zhang, J.; Liu, G.; Cao, Z.; Wang, Y.; Wang, W.; Li, S. Effects of Different Parts on the Chemical Composition, Silage Fermentation Profile, In Vitro and In Situ Digestibility of Paper Mulberry. Animals 2021, 11, 413. https://doi.org/10.3390/ani11020413
Hao Y, Huang S, Liu G, Zhang J, Liu G, Cao Z, Wang Y, Wang W, Li S. Effects of Different Parts on the Chemical Composition, Silage Fermentation Profile, In Vitro and In Situ Digestibility of Paper Mulberry. Animals. 2021; 11(2):413. https://doi.org/10.3390/ani11020413
Chicago/Turabian StyleHao, Yangyi, Shuai Huang, Gaokun Liu, Jun Zhang, Gang Liu, Zhijun Cao, Yajing Wang, Wei Wang, and Shengli Li. 2021. "Effects of Different Parts on the Chemical Composition, Silage Fermentation Profile, In Vitro and In Situ Digestibility of Paper Mulberry" Animals 11, no. 2: 413. https://doi.org/10.3390/ani11020413
APA StyleHao, Y., Huang, S., Liu, G., Zhang, J., Liu, G., Cao, Z., Wang, Y., Wang, W., & Li, S. (2021). Effects of Different Parts on the Chemical Composition, Silage Fermentation Profile, In Vitro and In Situ Digestibility of Paper Mulberry. Animals, 11(2), 413. https://doi.org/10.3390/ani11020413