
Utero-Placental Immune Milieu during Normal and Aglepristone-Induced Parturition in the Dog

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Samples Collection and Preservation
2.2. Total RNA Isolation, High Capacity Reverse Transcription, Pre-Amplification of cDNA and Semi-Quantitative Real-Time TaqMan PCR
2.3. Immunohistochemical Staining
3. Results
3.1. Pregnancy and Term (Prepartum Luteolysis)
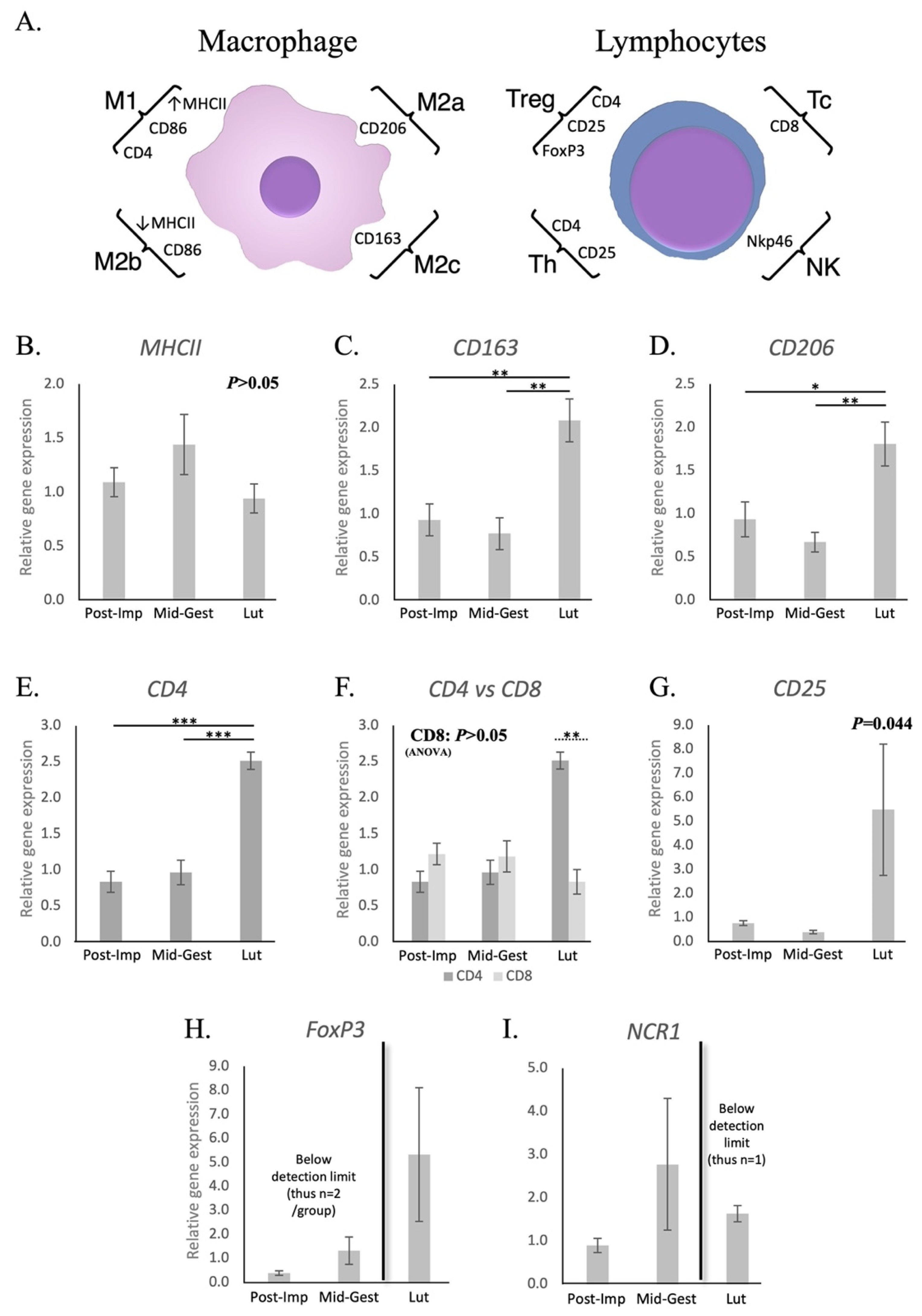
3.1.1. Macrophages and Lymphocytes
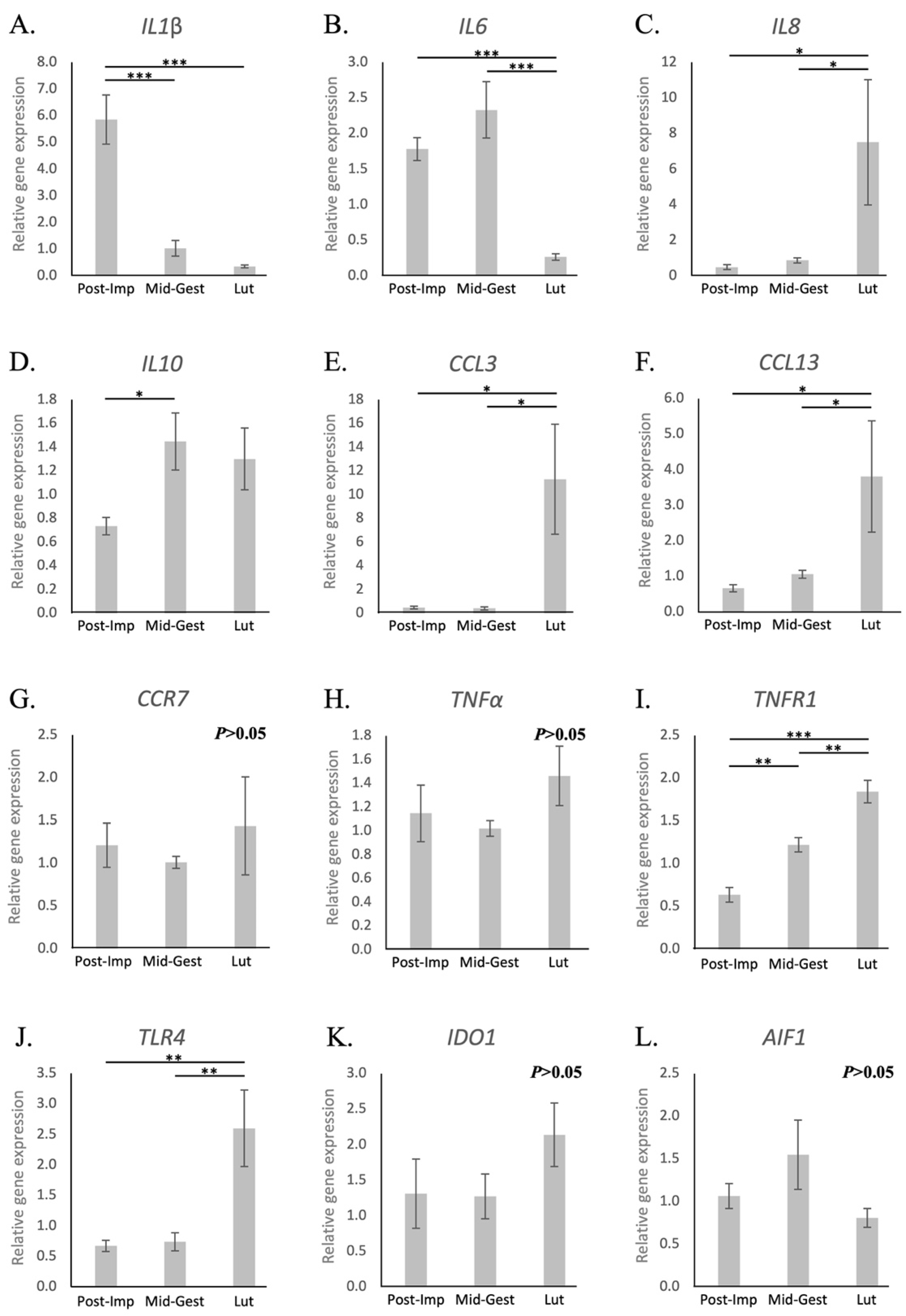
3.1.2. Cytokines and Other Immunomodulators
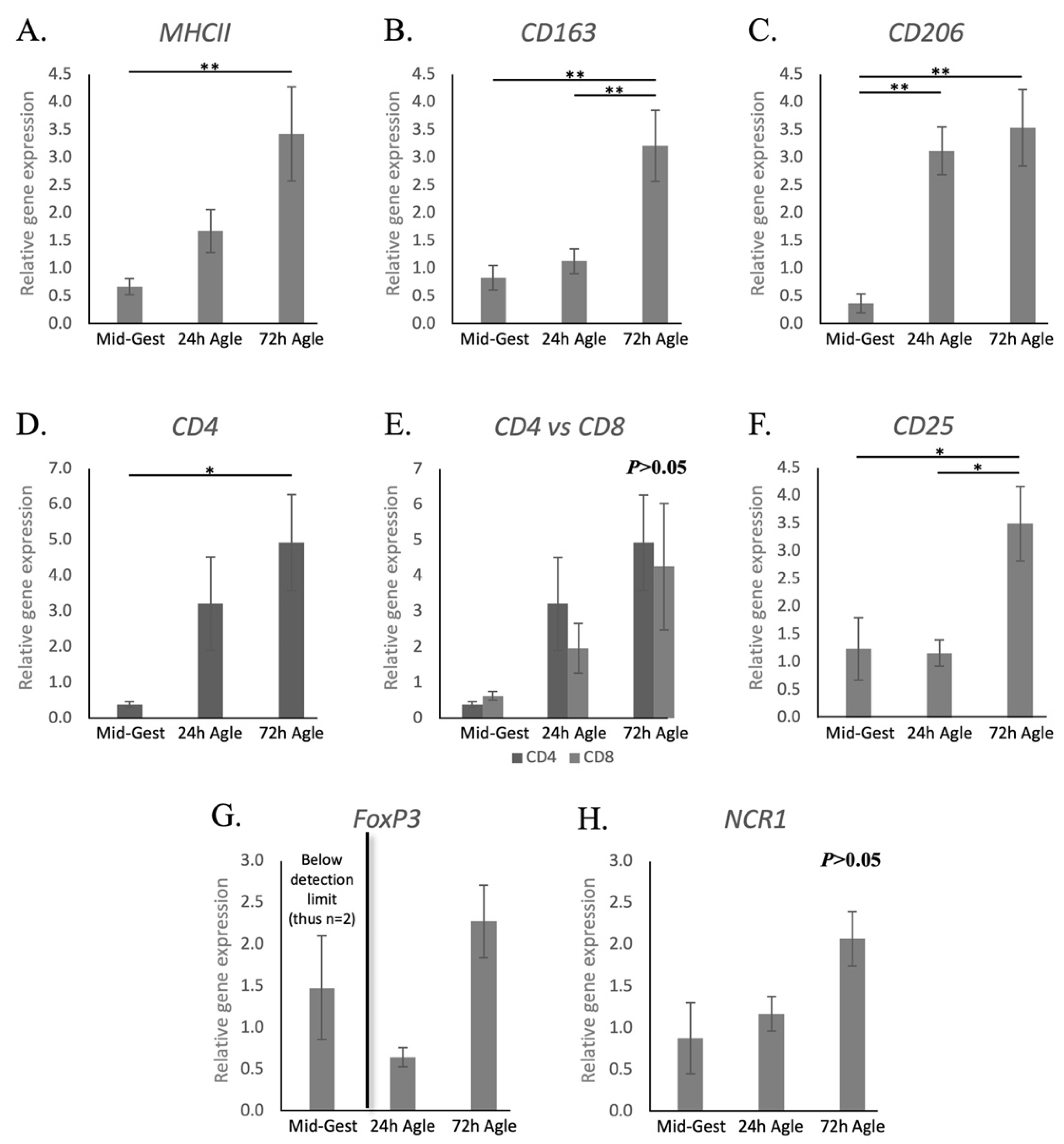
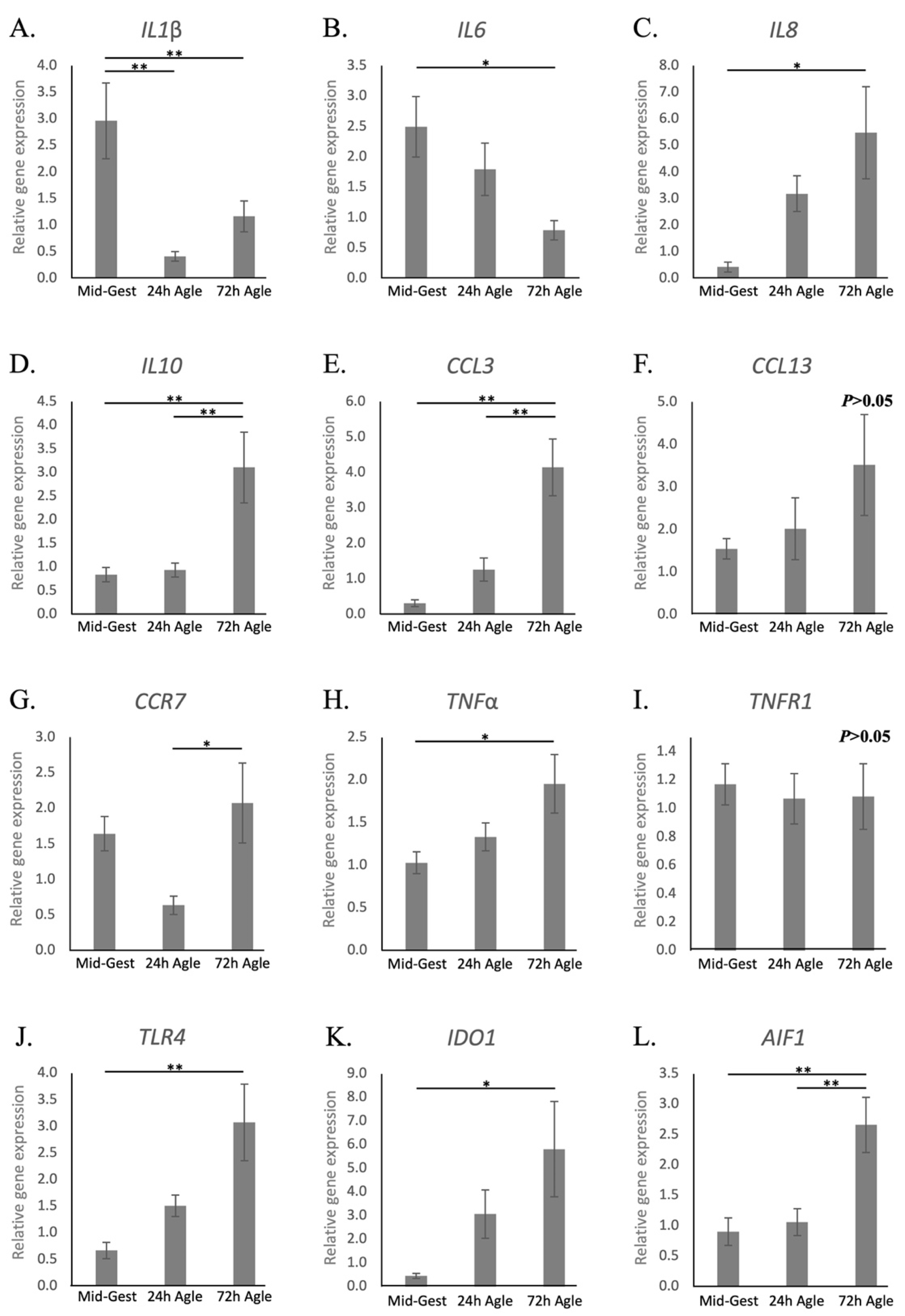
3.2. Aglepristone-Induced Luteolysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romero, R.; Dey, S.K.; Fisher, S.J. Preterm labor: One syndrome, many causes. Science 2014, 345, 760–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusterholz, C.; Hahn, S.; Holzgreve, W. Role of placentally produced inflammatory and regulatory cytokines in pregnancy and the etiology of preeclampsia. Semin. Immunopathol. 2007, 29, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Romero, R.; Gotsch, F.; Pineles, B.; Kusanovic, J.P. Inflammation in Pregnancy: Its Roles in Reproductive Physiology, Obstetrical Complications, and Fetal Injury. Nutr. Rev. 2007, 65, 194–202. [Google Scholar] [CrossRef]
- Szekeres-Bartho, J.; Polgar, B. PIBF: The double edged sword. Pregnancy and tumor. Am. J. Reprod. Immunol. 2010, 64, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Hebisch, G.; Neumaier-Wagner, P.M.; Huch, R.; von Mandach, U. Maternal serum interleukin-1 beta, -6 and -8 levels and potential determinants in pregnancy and peripartum. J. Perinat. Med. 2004, 32, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, C.; Trenk, L.; Aurich, J.; Wulf, M.; Aurich, C. Changes in blood pressure, heart rate, and blood profile in mares during the last 3 months of gestation and the peripartum period. Theriogenology 2016, 86, 1856–1864. [Google Scholar] [CrossRef]
- Magnusson, U.; Fossum, C. Numerical variations among blood mononuclear cells during the peripartal period in the gilt. J. Veter. Med. Ser. B 1990, 37, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.M.; Lai, P.F.; Imami, N.; Johnson, M.R. Progesterone-Related Immune Modulation of Pregnancy and Labor. Front. Endocrinol. (Lausanne) 2019, 10, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenavai, S.; Preissing, S.; Hoffmann, B.; Dilly, M.; Pfarrer, C.; Özalp, G.R.; Caliskan, C.; Seyrek-Intas, K.; Schuler, G. Investigations into the mechanisms controlling parturition in cattle. Reproduction 2012, 144, 279–292. [Google Scholar] [CrossRef] [Green Version]
- De Rensis, F.; Saleri, R.; Tummaruk, P.; Techakumphu, M.; Kirkwood, R.N. Prostaglandin F2alpha and control of reproduction in female swine: A review. Theriogenology 2012, 77, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ibuki, R.; Haga, N.; Muramatsu, S.; Mizumoto, A.; Itoh, Z. Long-Term Observations of Uterine Contractions in Nonpregnant Dogs. Biol. Reprod. 1997, 56, 632–639. [Google Scholar] [CrossRef]
- Gram, A.; Büchler, U.; Boos, A.; Hoffmann, B.; Kowalewski, M.P. Biosynthesis and degradation of canine placental prostaglandins: Prepartum changes in expression and function of prostaglandin F2alpha-synthase (PGFS, AKR1C3) and 15-hydroxyprostaglandin dehydrogenase (HPGD). Biol. Reprod. 2013, 89, 2. [Google Scholar] [CrossRef] [PubMed]
- Luz, M.R.; Bertan, C.M.; Binelli, M.; Lopes, M.D. Plasma concentrations of 13,14-dihydro-15-keto prostaglandin F2-alpha (PGFM), progesterone and estradiol in pregnant and nonpregnant diestrus cross-bred bitches. Theriogenology 2006, 66, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Kowalewski, M.P. Luteal regression vs. prepartum luteolysis: Regulatory mechanisms governing canine corpus luteum function. Reprod. Biol. 2014, 14, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Concannon, P.W.; Hansel, W. Prostaglandin F2α induced luteolysis, hypothermia, and abortions in beagle bitches. Prostaglandins 1977, 13, 533–542. [Google Scholar] [CrossRef]
- Nishiyama, T.; Tsumagari, S.; Ito, M.; Kimura, J.; Watanabe, G.; Taya, K.; Takeishi, M. Immunohistochemical study of steroidogenic enzymes in the ovary and placenta during pregnancy in the dog. Anat. Hist. Embryol. 1999, 28, 125–129. [Google Scholar] [CrossRef]
- Hoffmann, B.; Höveler, R.; Nohr, B.; Hasan, S.H. Investigations on hormonal changes around parturition in the dog and the occurrence of pregnancy-specific non conjugated oestrogens. Exp. Clin. Endocrinol. 1994, 102, 185–189. [Google Scholar] [CrossRef]
- Concannon, P.W.; McCann, J.P.; Temple, M. Biology and endocrinology of ovulation, pregnancy and parturition in the dog. J. Reprod. Fertil. Suppl. 1989, 39, 3–25. [Google Scholar]
- Kowalewski, M.P.; Pereira, M.T.; Papa, P.; Gram, A. Progesterone receptor blockers: Historical perspective, mode of function and insights into clinical and scientific applications. Tierarztl. Prax. Ausg. K Kleintiere Heimtiere 2020, 48, 433–440. [Google Scholar] [CrossRef]
- van Engelen, E.; de Groot, M.W.; Breeveld-Dwarkasing, V.; Everts, M.; van der Weyden, G.C.; Taverne, M.; Rutten, V. Cervical ripening and parturition in cows are driven by a cascade of pro-inflammatory cytokines. Reprod. Domest. Anim. 2009, 44, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Thomson, A.J.; Ledingham, M.; Jordan, F.; Greer, I.A.; Norman, J.E. Immunolocalization of Proinflammatory Cytokines in Myometrium, Cervix, and Fetal Membranes During Human Parturition at Term. Biol. Reprod. 2002, 66, 445–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiaens, I.; Zaragoza, D.B.; Guilbert, L.; Robertson, S.A.; Mitchell, B.F.; Olson, D.M. Inflammatory processes in preterm and term parturition. J. Reprod. Immunol. 2008, 79, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.M.; Chamley, L.; Keelan, J.A.; Mitchell, M.D. Cytokines of the placenta and extra-placental membranes: Roles and regulation during human pregnancy and parturition. Placenta 2002, 23, 257–273. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; StLouis, D.; Lehr, M.A.; Sanchez-Rodriguez, E.N.; Arenas-Hernandez, M. Immune cells in term and preterm labor. Cell Mol. Immunol. 2014, 11, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol. Hum. Reprod. 2003, 9, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Dudley, D.J.; Collmer, D.; Mitchell, M.D.; Trautman, M.S. Inflammatory cytokine mRNA in human gestational tissues: Implications for term and preterm labor. J. Soc. Gynecol. Investig. 1996, 3, 328–335. [Google Scholar] [CrossRef]
- Hamilton, S.; Oomomian, Y.; Stephen, G.; Shynlova, O.; Tower, C.L.; Garrod, A.; Lye, S.J.; Jones, R.L. Macrophages infiltrate the human and rat decidua during term and preterm labor: Evidence that decidual inflammation precedes labor. Biol. Reprod. 2012, 86, 39. [Google Scholar] [CrossRef]
- Brown, M.B.; von Chamier, M.; Allam, A.B.; Reyes, L. M1/M2 macrophage polarity in normal and complicated pregnancy. Front. Immunol. 2014, 5, 606. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Romero, R.; Miller, D.; Kadam, L.; Mial, T.N.; Plazyo, O.; Garcia-Flores, V.; Hassan, S.S.; Xu, Z.; Tarca, A.L.; et al. An M1-like Macrophage Polarization in Decidual Tissue during Spontaneous Preterm Labor That Is Attenuated by Rosiglitazone Treatment. J. Immunol. 2016, 196, 2476–2491. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Hernandez, M.; Gomez-Lopez, N.; Garcia-Flores, V.; Rangel-Escareño, C.; Alvarez-Salas, L.M.; Martinez-Acuña, N.; Vazquez-Perez, J.A.; Vega-Sanchez, R. Choriodecidual leukocytes display a unique gene expression signature in spontaneous labor at term. Genes. Immun. 2019, 20, 56–68. [Google Scholar] [CrossRef]
- Hooshmandabbasi, R.; Kazemian, A.; Zerbe, H.; Kowalewski, M.P.; Klisch, K. Macrophages in bovine term placenta: An ultrastructural and molecular study. Reprod. Domest. Anim. 2021, 56, 1243–1253. [Google Scholar] [CrossRef]
- Kimura, A.; Kishimoto, T. IL-6: Regulator of Treg/Th17 balance. Eur. J. Immunol. 2010, 40, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Physiology and Endocrinology Symposium: Maternal immunological adjustments to pregnancy and parturition in ruminants and possible implications for postpartum uterine health: Is there a prepartum–postpartum nexus? J. Anim. Sci. 2013, 91, 1639–1649. [Google Scholar] [CrossRef]
- Meeusen, E.N.; Bischof, R.J.; Lee, C.-S. Comparative T-cell responses during pregnancy in large animals and humans. Am. J. Reprod. Immunol. 2001, 46, 169–179. [Google Scholar] [CrossRef]
- Nelli, R.K.; De Koster, J.; Roberts, J.N.; de Souza, J.; Lock, A.L.; Raphael, W.; Agnew, D.; Contreras, G.A. Impact of uterine macrophage phenotype on placental retention in dairy cows. Theriogenology 2019, 127, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Goff, J.P.; Kehrli, M.E., Jr.; Reinhardt, T.A. Decreased neutrophil function as a cause of retained placenta in dairy cattle. J. Dairy Sci. 2002, 85, 544–550. [Google Scholar] [CrossRef]
- Tavares Pereira, M.; Nowaczyk, R.; Payan-Carreira, R.; Miranda, S.; Aslan, S.; Kaya, D.; Kowalewski, M.P. Selected uterine immune events associated with the establishment of pregnancy in the dog. Front. Vet. Sci. 2021, 7, 625921. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Rehrauer, H.; Ay, S.S.; Findik, M.; Boos, A.; Kautz, E.; Kowalewski, M.P. Gene expression profiling of the canine placenta during normal and antigestagen-induced luteolysis. Gen. Comp. Endocrinol. 2019, 282, 113194. [Google Scholar] [CrossRef] [PubMed]
- Pires, M.A.; Payan-Carreira, R. Resident Macrophages and Lymphocytes in the Canine Endometrium. Reprod. Domest. Anim. 2015, 50, 740–749. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Beceriklisoy, H.B.; Pfarrer, C.; Aslan, S.; Kindahl, H.; Kücükaslan, I.; Hoffmann, B. Canine placenta: A source of prepartal prostaglandins during normal and antiprogestin-induced parturition. Reproduction 2010, 139, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Kowalewski, M.P.; Beceriklisoy, H.B.; Aslan, S.; Agaoglu, A.R.; Hoffmann, B. Time related changes in luteal prostaglandin synthesis and steroidogenic capacity during pregnancy, normal and antiprogestin induced luteolysis in the bitch. Anim. Reprod. Sci. 2009, 116, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, E.C. Placentation. In Marshall’s Physiology of Reproduction; Parkes, A.S., Ed.; Longmans, Greens and Co.: London, UK, 1952; Volume II. [Google Scholar]
- Kehrer, A. Zur Entwicklung und Ausbildung des Chorions der Placenta zonaria bei Katze, Hund und Fuchs. Z. Anat. Entwickl. 1973, 143, 25–42. [Google Scholar] [CrossRef]
- Pereira, M.T.; Gram, A.; Nowaczyk, R.; Boos, A.; Hoffmann, B.; Janowski, T.; Kowalewski, M.P. Prostaglandin-mediated effects in early canine corpus luteum: In vivo effects on vascular and immune factors. Reprod. Biol. 2019, 10, 100–111. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Meyer, A.; Hoffmann, B.; Aslan, S.; Boos, A. Expression and functional implications of peroxisome proliferator—Activated receptor gamma (PPARgamma) in canine reproductive tissues during normal pregnancy and parturition and at antiprogestin induced abortion. Theriogenology 2011, 75, 877–886. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Schuler, G.; Taubert, A.; Engel, E.; Hoffmann, B. Expression of cyclooxygenase 1 and 2 in the canine corpus luteum during diestrus. Theriogenology 2006, 66, 1423–1430. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Gram, A.; Fox, B.; Büchler, U.; Boos, A.; Hoffmann, B.; Kowalewski, M.P. Canine placental prostaglandin E2 synthase: Expression, localization, and biological functions in providing substrates for prepartum PGF2alpha synthesis. Biol. Reprod. 2014, 91, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Montaldo, E.; Del Zotto, G.; Della Chiesa, M.; Mingari, M.C.; Moretta, A.; De Maria, A.; Moretta, L. Human NK cell receptors/markers: A tool to analyze NK cell development, subsets and function. Cytom. Part A 2013, 83, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Nancy, P.; Erlebacher, A. T cell behavior at the maternal-fetal interface. Int. J. Dev. Biol. 2014, 58, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruocco, M.G.; Chaouat, G.; Florez, L.; Bensussan, A.; Klatzmann, D. Regulatory T-cells in pregnancy: Historical perspective, state of the art, and burning questions. Front. Immunol. 2014, 5, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aluvihare, V.R.; Kallikourdis, M.; Betz, A.G. Regulatory T cells mediate maternal tolerance to the fetus. Nat. Immunol. 2004, 5, 266–271. [Google Scholar] [CrossRef]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of Regulatory T Cell Development by the Transcription Factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef]
- Hansen, V.L.; Faber, L.S.; Salehpoor, A.A.; Miller, R.D. A pronounced uterine pro-inflammatory response at parturition is an ancient feature in mammals. Proc. Biol. Sci. 2017, 284, 20171694. [Google Scholar] [CrossRef] [Green Version]
- de Mestre, A.; Noronha, L.; Wagner, B.; Antczak, D.F. Split immunological tolerance to trophoblast. Int. J. Dev. Biol. 2010, 54, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Mold, J.E.; McCune, J.M. Immunological tolerance during fetal development: From mouse to man. Adv. Immunol. 2012, 115, 73–111. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Hanna, N.; Bonifacio, L.; Weinberger, B.; Reddy, P.; Murphy, S.; Romero, R.; Sharma, S. Evidence for interleukin-10-mediated inhibition of cyclo-oxygenase-2 expression and prostaglandin production in preterm human placenta. Am. J. Reprod. Immunol. 2006, 55, 19–27. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Vega-Sanchez, R.; Castillo-Castrejon, M.; Romero, R.; Cubeiro-Arreola, K.; Vadillo-Ortega, F. Evidence for a role for the adaptive immune response in human term parturition. Am. J. Reprod. Immunol. 2013, 69, 212–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeaman, G.R.; Howell, A.L.; Weldon, S.; Demian, D.J.; Collins, J.E.; O’Connell, D.M.; Asin, S.N.; Wira, C.R.; Fanger, M.W. Human immunodeficiency virus receptor and coreceptor Expression on human uterine epithelial cells: Regulation of expression during the menstrual cycle and implications for human immunodeficiency virus infection. Immunology 2003, 109, 137–146. [Google Scholar] [CrossRef]
- Gaynor, L.M.; Colucci, F. Uterine Natural Killer Cells: Functional Distinctions and Influence on Pregnancy in Humans and Mice. Front. Immunol. 2017, 8, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, K.; Mor, G. Toll-like receptors at the maternal-fetal interface in normal pregnancy and pregnancy disorders. Am. J. Reprod. Immunol. 2010, 63, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Schatz, F.; Moore, R.M.; Mercer, B.M.; Rangaswamy, N.; Mansour, J.M.; Lockwood, C.J.; Moore, J.J. The effects of thrombin and cytokines upon the biomechanics and remodeling of isolated amnion membrane, in vitro. Placenta 2011, 32, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Nadeau-Vallée, M.; Obari, D.; Quiniou, C.; Lubell, W.D.; Olson, D.M.; Girard, S.; Chemtob, S. A critical role of interleukin-1 in preterm labor. Cytokine Growth Factor Rev. 2016, 28, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Gogny A, Fieni F: Aglepristone: A review on its clinical use in animals. Theriogenology 2016, 85, 555–566. [CrossRef] [Green Version]
- Jurka, P.; Max, A.; Hawrynska, K.; Snochowski, M. Age-related pregnancy results and further examination of bitches after aglepristone treatment of pyometra. Reprod. Domest. Anim. 2010, 45, 525–529. [Google Scholar] [CrossRef]
- Fieni, F.; Tainturier, D.; Bruyas, J.F.; Badinand, F.; Berthelot, X.; Ronsin, P.; Rachail, M.; Lefay, M.P. Etude clinique d’une anti-hormone pour provoquer l’avortement chez la chienne. Rec. Med. Vet. 1996, 172, 359–367. [Google Scholar]
- Martinez, F.O.; Gordon, S.; Locati, M.; Mantovani, A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: New molecules and patterns of gene expression. J. Immunol. 2006, 177, 7303–7311. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavares Pereira, M.; Nowaczyk, R.; Aslan, S.; Ay, S.S.; Kowalewski, M.P. Utero-Placental Immune Milieu during Normal and Aglepristone-Induced Parturition in the Dog. Animals 2021, 11, 3598. https://doi.org/10.3390/ani11123598
Tavares Pereira M, Nowaczyk R, Aslan S, Ay SS, Kowalewski MP. Utero-Placental Immune Milieu during Normal and Aglepristone-Induced Parturition in the Dog. Animals. 2021; 11(12):3598. https://doi.org/10.3390/ani11123598
Chicago/Turabian StyleTavares Pereira, Miguel, Renata Nowaczyk, Selim Aslan, Serhan S. Ay, and Mariusz P. Kowalewski. 2021. "Utero-Placental Immune Milieu during Normal and Aglepristone-Induced Parturition in the Dog" Animals 11, no. 12: 3598. https://doi.org/10.3390/ani11123598
APA StyleTavares Pereira, M., Nowaczyk, R., Aslan, S., Ay, S. S., & Kowalewski, M. P. (2021). Utero-Placental Immune Milieu during Normal and Aglepristone-Induced Parturition in the Dog. Animals, 11(12), 3598. https://doi.org/10.3390/ani11123598