
Dietary Supplementation with Microalgae (Schizochytrium sp.) Improves the Antioxidant Status, Fatty Acids Profiles and Volatile Compounds of Beef

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design and Diets
2.2. Sample Collection
2.3. Analysis of Antioxidant Enzymes Activity and Lipid Oxidation
2.4. Meat Physicochemical Quality Characteristics Analysis
2.4.1. pH and Color Measurement
2.4.2. Drip Loss and Cooking Loss Determination
2.4.3. Proximate Composition Analysis
2.5. Fatty Acids Analysis
2.6. Volatile Compounds Analysis
2.7. Sensory Analysis
2.8. Statistical Analysis
3. Results
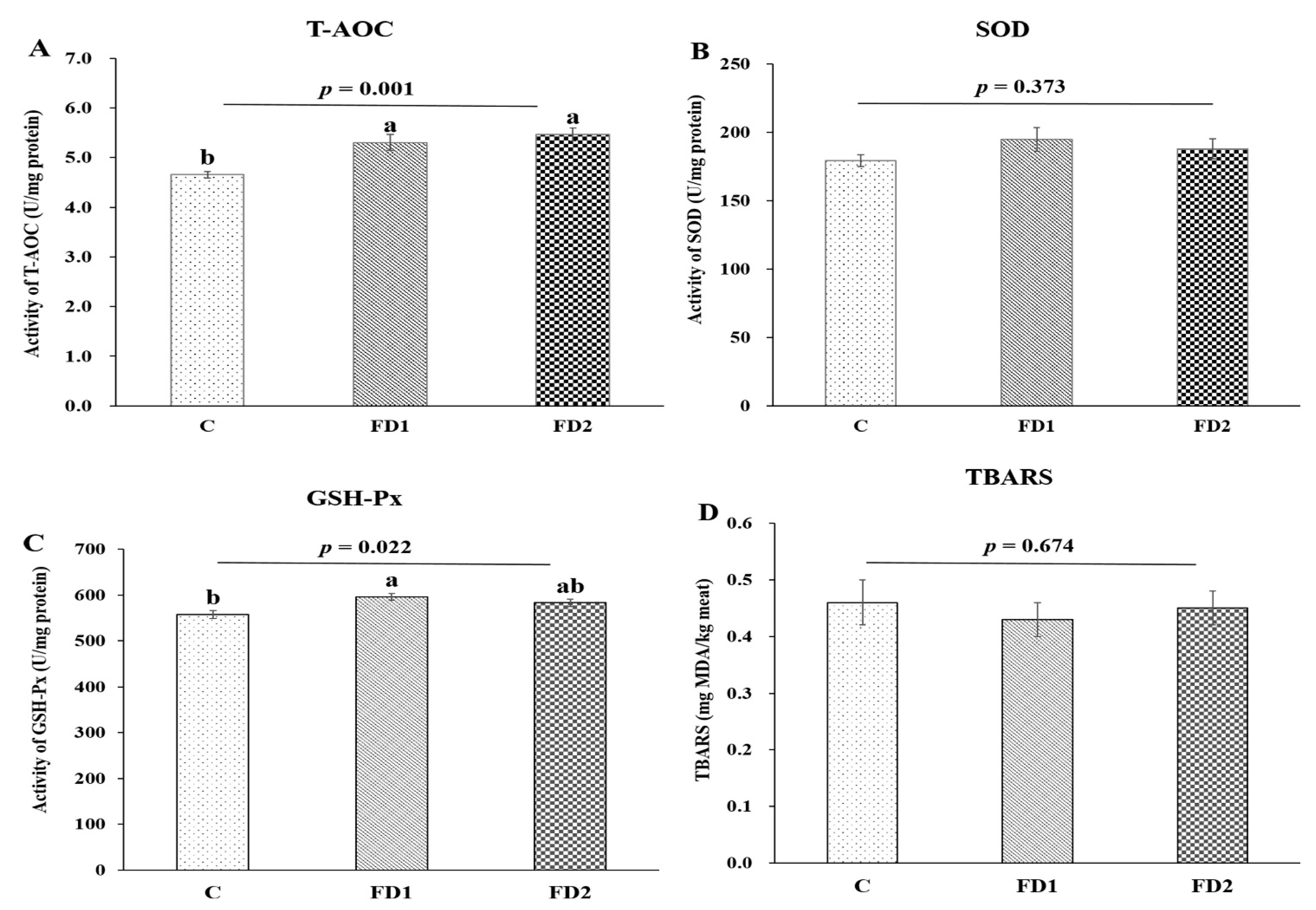
3.1. Antioxidant Enzymes Activity and Lipid Oxidation
3.2. Meat Physicochemical Quality
3.3. Fatty Acid Composition
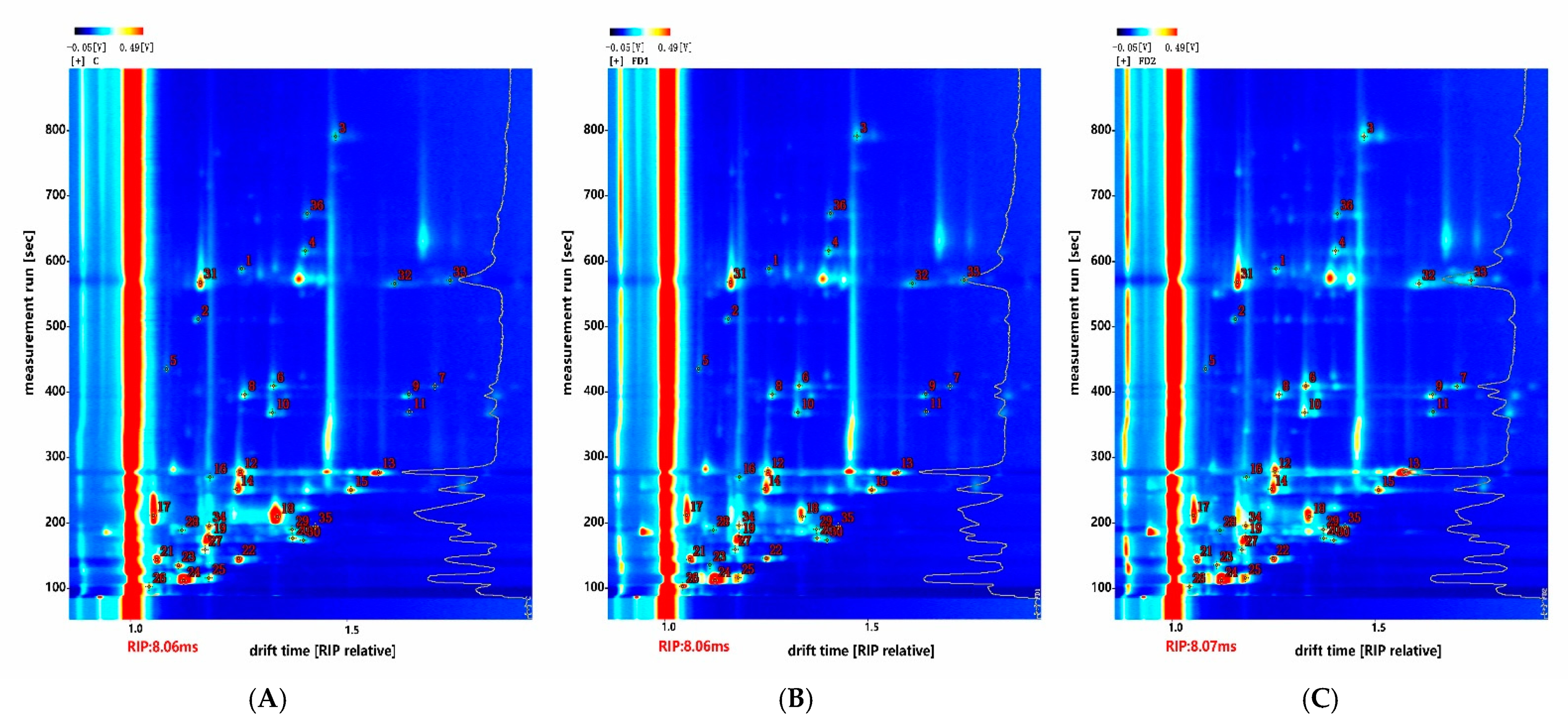
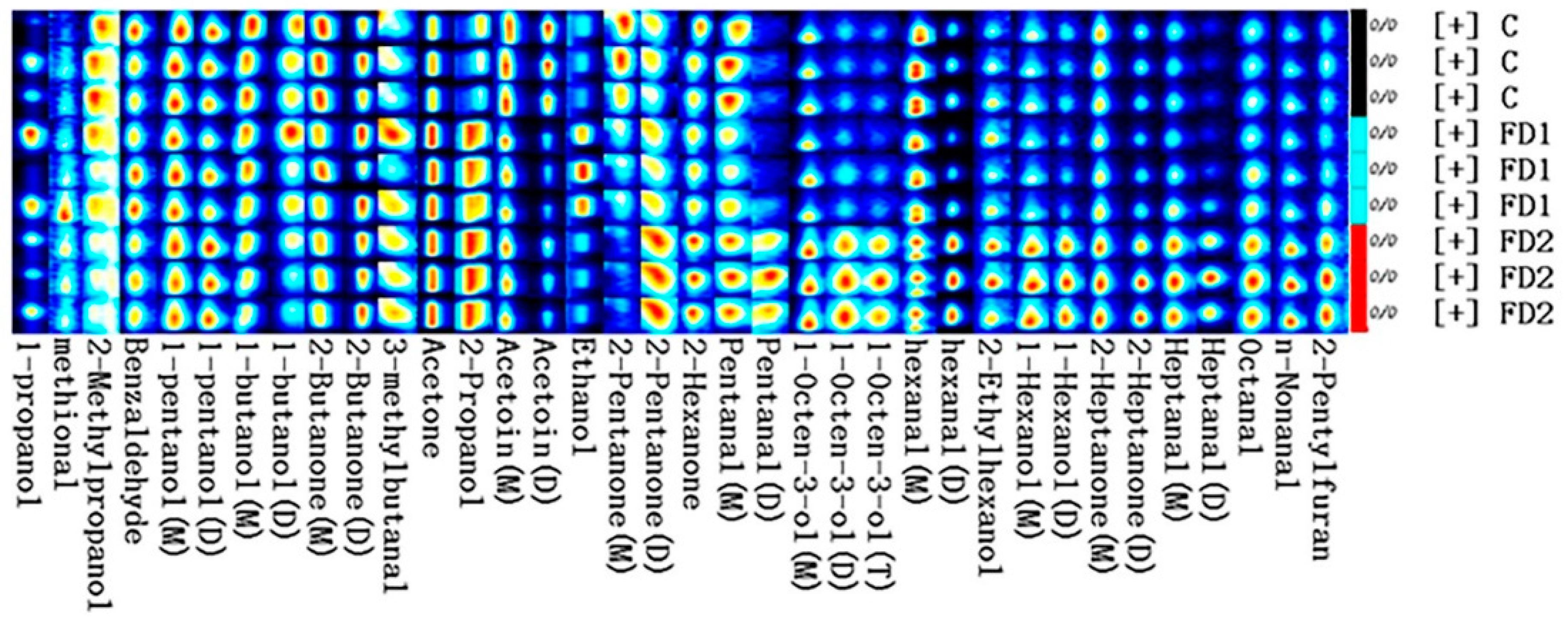
3.4. Volatile Compounds
3.5. Sensory Evaluation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, X.Z.; Yan, C.G.; Zan, L.S. Current situation and future prospects for beef production in China—A review. Asian-Australas J. Anim. Sci. 2018, 31, 984–991. [Google Scholar] [CrossRef]
- Toral, P.G.; Hervás, G.; Carreño, D.; Leskinen, H.; Belenguer, A.; Shingfield, K.J.; Frutos, P. In vitro response to EPA, DPA, and DHA: Comparison of effects on ruminal fermentation and biohydrogenation of 18-carbon fatty acids in cows and ewes. J. Dairy Sci. 2017, 100, 6187–6198. [Google Scholar] [CrossRef]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Colussi, G.; Catena, C.; Novello, M.; Bertin, N.; Sechi, L.A. Impact of omega-3 polyunsaturated fatty acids on vascular function and blood pressure: Relevance for cardiovascular outcomes. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 191–200. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Gavala, E.; Christea, M.; Andrikopoulos, N.K. Nutritional evaluation and bioactive microconstituents (carotenoids, tocopherols, sterols and squalene) of raw and roasted chicken fed on DHA-rich microalgae. Food Res. Int. 2010, 43, 2006–2013. [Google Scholar] [CrossRef]
- Lagarde, M. Docosahexaenoic acid: Nutrient and precursor of bioactive lipids. Eur. J. Lipid. Sci. Technol. 2008, 110, 673–678. [Google Scholar] [CrossRef]
- Plourde, M.; Cunnane, S.C. Extremely limited synthesis of long chain polyunsaturates in adults: Implications for their dietary essentiality and use as supplements. Appl. Physiol. Nutr. Metab. 2007, 32, 619–634. [Google Scholar] [CrossRef] [Green Version]
- Bryhni, E.A.; Kjos, N.P.; Ofstad, R.; Hunt, M. Polyunsaturated fat and fish oil in diets for growing-finishing pigs: Effects on fatty acid composition and meat, fat, and sausage quality. Meat Sci. 2002, 62, 1–8. [Google Scholar] [CrossRef]
- Kahraman, R.; Özpınar, H.; Abas, İ.; Kutay, H.C.; Eseceli, H.; Grashorn, M. Effects of different dietary oil sources on fatty acid composition and malondialdehyde levels of thigh meat in broiler chickens. Arch. Geflugelkd. 2004, 68, 77–86. [Google Scholar]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Altomonte, I.; Salari, F.; Licitra, R.; Martini, M. Use of microalgae in ruminant nutrition and implications on milk quality—A review. Livest. Sci. 2018, 214, 25–35. [Google Scholar] [CrossRef]
- Mastellone, V.; Morittu, V.M.; Musco, N.; Spina, A.A.; Malgeri, A.; Molinari, M.L.; D’Aniello, B.; Infascelli, F.; Tudisco, R.; Lombardi, P. Dietary supplementation with a phytocomplex affects blood parameters and milk yield and quality in grazing goats. Small Ruminant Res. 2021, 201, 106421. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Mathys, A. Trends in microalgae incorporation into innovative food products with potential health benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef]
- Long, S.F.; Kang, S.; Wang, Q.Q.; Xu, Y.T.; Pan, L.; Hu, J.X.; Li, M.; Piao, X.S. Dietary supplementation with DHA-rich microalgae improves performance, serum composition, carcass trait, antioxidant status, and fatty acid profile of broilers. Poult. Sci. 2018, 97, 1881–1890. [Google Scholar] [CrossRef] [PubMed]
- Abdelnour, S.A.; Sheiha, A.M.; Taha, A.E.; Swelum, A.A.; Alarifi, S.; Alkahtani, S.; Ali, D.; AlBasher, G.; Almeer, R.; Falodah, F.; et al. Impacts of enriching growing rabbit diets with chlorella vulgaris microalgae on growth, blood variables, carcass traits, immunological and antioxidant indices. Animals 2019, 9, 788. [Google Scholar] [CrossRef] [Green Version]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of dietary microalgae on growth performance, profiles of amino and fatty acids, antioxidant status, and meat quality of broiler chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Malau-Aduli, A.E.O. Spirulina as a livestock supplement and animal feed. J. Anim. Physiol. Anim. Nutr. 2013, 97, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Abril, R.; Garrett, J.; Zeller, S.G.; Sander, W.J.; Mast, R.W. Safety assessment of DHA-rich microalgae from Schizochytrium sp. Part V: Target animal safety/toxicity study in growing swine. Regul. Toxicol. Pharmacol. 2003, 37, 73–82. [Google Scholar] [CrossRef]
- Li, M.H.; Robinson, E.H.; Tucker, C.S.; Manning, B.B.; Khoo, L. Effects of dried algae Schizochytrium sp., a rich source of docosahexaenoic acid, on growth, fatty acid composition, and sensory quality of channel catfish Ictalurus punctatus. Aquaculture 2009, 292, 232–236. [Google Scholar] [CrossRef]
- Demeda, M.A.; Tomaluski, C.R.; Baggio, C.; Mateus, K.A.; Petrolli, T.G.; Mueller, L.F.; Pereira, A.S.C.; Griebler, L.; Zotti, C.A. Feeding microalgae (Schizochytrium limacinum) to beef steers increases meat omega-3 content. Res. Soc. Dev. 2020, 9, e675974568. [Google Scholar] [CrossRef]
- de Lima Valença, R.; da Silva Sobrinho, A.G.; Borghi, T.H.; Meza, D.A.R.; de Andrade, N.; Silva, L.G.; Bezerra, L.R. Performance, carcass traits, physicochemical properties and fatty acids composition of lamb’s meat fed diets with marine microalgae meal (Schizochytrium sp.). Livest. Sci. 2021, 243, 104387. [Google Scholar] [CrossRef]
- Do Nascimento, T.C.; Cazarin, C.B.; Marostica, M.R., Jr.; Risso, É.M.; Amaya-Farfan, J.; Grimaldi, R.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Microalgae biomass intake positively modulates serum lipid profile and antioxidant status. J. Funct. Foods 2019, 58, 11–20. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2005; pp. 12–13. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Tian, J.; Zhou, J.; Yu, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Fabre, R.; Dalzotto, G.; Perlo, F.; Bonato, P.; Teira, G.; Tisocco, O. Cooking method effect on Warner-Bratzler shear force of different beef muscles. Meat Sci. 2018, 138, 10–14. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; Pearce, K.M.; Mortimer, S.I.; Pethick, D.W.; Ball, A.J.; Hopkins, D.L. Sources of variation of health claimable long chain omega-3 fatty acids in meat from Australian lamb slaughtered at similar weights. Meat Sci. 2014, 96, 1095–1103. [Google Scholar] [CrossRef]
- Aldai, N.; Lavín, P.; Kramer, J.K.; Jaroso, R.; Mantecón, A.R. Breed effect on quality veal production in mountain areas: Emphasis on meat fatty acid composition. Meat Sci. 2012, 92, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.C.; Yu, H.; Xie, P.; Sun, B.Z.; Wang, X.Y.; Zhang, S.S. Influence of electrostatic field on the quality attributes and volatile flavor compounds of dry-cured beef during chill storage. Foods 2020, 9, 478. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Lin, W.C.; Yu, B.; Lee, T.T. Antioxidant capacity of phytochemicals and their potential effects on oxidative status in animals—A review. Asian-Australas J. Anim. Sci. 2017, 30, 299. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. J. Clin. Investig. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Delles, R.M.; Xiong, Y.L.; True, A.D.; Ao, T.; Dawson, K.A. Dietary antioxidant supplementation enhances lipid and protein oxidative stability of chicken broiler meat through promotion of antioxidant enzyme activity. Poult Sci. 2014, 93, 1561–1570. [Google Scholar] [CrossRef]
- Mirzaie, S.; Zirak-Khattab, F.; Hosseini, S.A.; Donyaei-Darian, H. Effects of dietary Spirulina on antioxidant status, lipid profile, immune response and performance characteristics of broiler chickens reared under high ambient temperature. Asian-Australas J. Anim. Sci. 2018, 31, 556. [Google Scholar] [CrossRef] [PubMed]
- La, A.L.T.Z.; Pierce, K.M.; Liu, W.H.; Gao, S.T.; Bu, D.P.; Ma, L. Supplementation with Schizochytrium sp. enhances growth performance and antioxidant capability of dairy calves before weaning. Anim. Feed Sci. Technol. 2021, 271, 114779. [Google Scholar]
- Wang, B.; Xu, C.; Liu, C.; Qu, Y.; Zhang, H.; Luo, H. The effect of dietary lycopene supplementation on drip loss during storage of lamb meat by iTRAQ analysis. Antioxidants 2021, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Xing, J.; Li, H.; Xu, X.; Hu, Z.; Ji, H. Effects of the defatted Schizochytrium sp. on growth performance, fatty acid composition, histomorphology and antioxidant status of juvenile mirror carp (Cyprinus carpio var. specularis). Aquac. Res. 2021, 3, 3062–3076. [Google Scholar] [CrossRef]
- Zhang, Y.; Holman, B.W.; Mao, Y.; Chen, X.; Luo, X.; Hopkins, D.L.; Zhang, Y. Determination of a pH threshold for dark cutting beef based on visual evaluation by Asian consumers. Meat Sci. 2021, 172, 108347. [Google Scholar] [CrossRef]
- Ribeiro, T.; Lordelo, M.M.; Alves, S.P.; Bessa, R.J.B.; Costa, P.; Lemos, J.P.C.; Ferreira, L.M.A.; Fontes, C.M.G.A.; Prates, J.A.M. Direct supplementation of diet is the most efficient way of enriching broiler meat with n-3 long-chain polyunsaturated fatty acids. Br. Poult. Sci. 2013, 54, 753–765. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Clayton, E.H.; Lamb, T.A.; Van de Ven, R.J.; Refshauge, G.; Kerr, M.J.; Bailes, K.; Lewandowski, P.; Ponnampalam, E.N. The impact of supplementing lambs with algae on growth, meat traits and oxidative status. Meat Sci. 2014, 98, 135–141. [Google Scholar] [CrossRef]
- Sardi, L.; Martelli, G.; Lambertini, L.; Parisini, P.; Mordenti, A. Effects of a dietary supplement of DHA-rich marine algae on Italian heavy pig production parameters. Livest. Sci. 2006, 103, 95–103. [Google Scholar] [CrossRef]
- Pannier, L.; Gardner, G.E.; Pearce, K.L.; McDonagh, M.; Ball, A.J.; Jacob, R.H.; Pethick, D.W. Associations of sire estimated breeding values and objective meat quality measurements with sensory scores in Australian lamb. Meat Sci. 2014, 96, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, O.; Mendizabal, J.A.; Insausti, K.; Soret, B.; Purroy, A.; Arana, A. Effects of addition of linseed and marine algae to the diet on adipose tissue development, fatty acid profile, lipogenic gene expression, and meat quality in lambs. PLoS ONE 2016, 11, e0156765. [Google Scholar] [CrossRef] [Green Version]
- Kalbe, C.; Priepke, A.; Nürnberg, G.; Dannenberger, D. Effects of long-term microalgae supplementation on muscle microstructure, meat quality and fatty acid composition in growing pigs. J. Anim. Phys. Anim. Nutri. 2019, 103, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Givens, D.I.; Kliem, K.E.; Gibbs, R.A. The role of meat as a source of n−3 polyunsaturated fatty acids in the human diet. Meat Sci. 2006, 74, 209–218. [Google Scholar] [CrossRef]
- Nutrition Information Requirements. Available online: http://www.foodstandards.gov.au/foodstandards/userguides/nutritioninformation1406.cfm (accessed on 20 July 2016).
- Commission Regulation of European Union. Amending Regulation (EC) No 1924/2006 of the European Parliament and of the Council with regard to the list of nutrition claims. Off. J. Eur. Union. 2010, 37, 16–18. [Google Scholar]
- Calder, P.C. Very long-chain n-3 fatty acids and human health: Fact, fiction and the future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [Green Version]
- Burghardt, P.R.; Kemmerer, E.S.; Buck, B.J.; Osetek, A.J.; Yan, C.; Koch, L.G.; Britton, S.L.; Evans, S.J. Dietary n-3: n-6 fatty acid ratios differentially influence hormonal signature in a rodent model of metabolic syndrome relative to healthy controls. Nutr. Metab. 2010, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.Y.; Xu, C.Y.; Chang, X.X.; Li, W.W.; Sun, L.Y.; Yang, X.B.; Yu, L.F. Fatty liver index correlates with non-alcoholic fatty liver disease, but not with newly diagnosed coronary artery atherosclerotic disease in Chinese patients. BMC Gastroenterol. 2013, 13, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kumar, F.; Tyagi, P.K.; Mir, N.A.; Dev, K.; Begum, J.; Biswas, A.; Sheikh, H.; Tyagi, P.; Sharma, D.; Sahu, B.; et al. Dietary flaxseed and turmeric is a novel strategy to enrich chicken meat with long chain ω-3 polyunsaturated fatty acids with better oxidative stability and functional properties. Food Chem. 2020, 305, 125458. [Google Scholar] [CrossRef]
- Rymer, C.; Gibbs, R.A.; Givens, D.I. Comparison of algal and fish sources on the oxidative stability of poultry meat and its enrichment with omega-3 polyunsaturated fatty acids. Poult. Sci. 2010, 89, 150–159. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, M.; Xie, J.; Zhao, M.; Hou, L.; Liang, J.; Wang, S.; Cheng, J. Volatile flavor constituents in the pork broth of black-pig. Food Chem. 2017, 226, 51–60. [Google Scholar] [CrossRef]
- Kerth, C.R.; Miller, R.K. Beef flavor: A review from chemistry to consumer. J. Sci. Food Agric. 2015, 95, 2783–2798. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Busboom, J.R.; Nelson, M.L.; Mengarelli, R. Omega-3 fatty acids affected human perception of ground beef negatively. Meat Sci. 2011, 89, 390–399. [Google Scholar] [CrossRef]
- Piao, M.Y.; Yong, H.I.; Lee, H.J.; Fassah, D.M.; Kim, H.J.; Jo, C.; Baik, M. Comparison of fatty acid profiles and volatile compounds among quality grades and their association with carcass characteristics in longissimus dorsi and semimembranosus muscles of Korean cattle steer. Livest. Sci. 2017, 198, 147–156. [Google Scholar] [CrossRef]
- Utama, D.T.; Lee, C.W.; Park, Y.S.; Jang, A.; Lee, S.K. Comparison of meat quality, fatty acid composition and aroma volatiles of Chikso and Hanwoo beef. Asian-Australas J. Anim. Sci. 2018, 31, 1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasta, V.; Priolo, A. Ruminant fat volatiles as affected by diet. A review. Meat Sci. 2006, 73, 218–228. [Google Scholar] [CrossRef]
- Morán, L.; Giráldez, F.J.; Panseri, S.; Aldai, N.; Jordán, M.J.; Chiesa, L.M.; Andrés, S. Effect of dietary carnosic acid on the fatty acid profile and flavour stability of meat from fattening lambs. Food Chem. 2013, 138, 2407–2414. [Google Scholar] [CrossRef]
- Brewer, M.S. Irradiation effects on meat flavor: A review. Meat Sci. 2009, 81, 1–14. [Google Scholar] [CrossRef]
- Song, S.; Tang, Q.; Fan, L.; Xu, X.; Song, Z.; Hayat, K.; Feng, T.; Wang, Y. Identification of pork flavour precursors from enzyme-treated lard using Maillard model system assessed by GC–MS and partial least squares regression. Meat Sci. 2017, 124, 15–24. [Google Scholar] [CrossRef]
- Legako, J.F.; Dinh, T.T.N.; Miller, M.F.; Adhikari, K.; Brooks, J.C. Consumer palatability scores, sensory descriptive attributes, and volatile compounds of grilled beef steaks from three USDA Quality Grades. Meat Sci. 2016, 112, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, B.; Kim, H.K.; Yun, Y.K.; Kang, S.J.; Kim, K.T.; Kim, B.D.; Kim, E.J.; Choi, Y.M. Sensory quality characteristics with different beef quality grades and surface texture features assessed by dented area and firmness, and the relation to muscle fiber and bundle characteristics. Meat Sci. 2018, 145, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Chail, A.; Legako, J.F.; Pitcher, L.R.; Ward, R.E.; Martini, S.; MacAdam, J.W. Consumer sensory evaluation and chemical composition of beef gluteus medius and triceps brachii steaks from cattle finished on forage or concentrate diets. J. Anim. Sci. 2017, 95, 1553–1564. [Google Scholar] [PubMed]
- Coelho, D.; Pestana, J.; Almeida, J.M.; Alfaia, C.M.; Fontes, C.M.; Moreira, O.; Prates, J.A. A high dietary incorporation level of Chlorella vulgaris improves the nutritional value of pork fat without impairing the performance of finishing pigs. Animals 2020, 10, 2384. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg Fed Basis) | Chemical Composition (g/kg DM Basis) 2 | ||
|---|---|---|---|
| Oat Hay | 200 | Crude protein | 122.0 |
| Alfalfa hay | 400 | Neutral detergent fiber | 315.5 |
| Corn | 216 | Acid detergent fiber | 169.0 |
| Wheat bran | 24 | Calcium | 5.2 |
| Wheat distillers dried grains with solubles | 20 | Phosphorus | 4.0 |
| Soybean meal | 48 | Net energy/(MJ/kg) 3 | 5.3 |
| Rapeseed meal | 40 | ||
| Cottonseed meal | 16 | ||
| Jujube powder | 12 | ||
| Fatty Acid Calcium | 4 | ||
| Sodium Chloride | 4 | ||
| Mineral and vitamin premix 1 | 16 | ||
| Fatty Acids, mg/g Diet | |||
| C14:0 | 0.13 | C16:1 | 0.10 |
| C15:0 | 0.02 | C18:1 n-9 | 12.08 |
| C16:0 | 9.19 | C18:2 n-6 | 16.44 |
| C17:0 | 0.05 | C18:3 n-3 (ALA) | 0.94 |
| C18:0 | 1.03 | C20:1 | 0.16 |
| C20:0 | 0.17 | C20:2 n-6 | 0.01 |
| C21:0 | 0.02 | C20:5 n-3 (EPA) | 0.02 |
| C22:0 | 0.10 | ||
| Fatty Acid, mg/g Dried Powder | |
|---|---|
| C14:0 | 1.96 |
| C15:0 | 5.07 |
| C16:0 | 73.04 |
| C17:0 | 4.33 |
| C18:0 | 3.26 |
| C20:0 | 0.84 |
| C21:0 | 0.02 |
| C22:0 | 0.53 |
| C23:0 | 0.11 |
| C16:1 | 0.56 |
| C18:1 n-9 | 0.03 |
| C20:1 | 0.06 |
| C18:2 n-6 | 0.15 |
| C20:2 n-6 | 0.24 |
| C18:3 n-3 (ALA) | 0.97 |
| C20:3 n-6 | 0.82 |
| C20:4 n-6 | 0.59 |
| C20:5 n-3 (EPA) | 1.90 |
| C22:6 n-3 (DHA) | 202.62 |
| Item | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| C | FD1 | FD2 | |||
| L* | 30.07 | 29.52 | 29.73 | 1.72 | 0.66 |
| a* | 15.17 | 14.34 | 15.73 | 2.51 | 0.64 |
| b* | 6.37 | 6.72 | 6.94 | 1.67 | 0.84 |
| C* | 16.47 | 15.93 | 17.23 | 2.66 | 0.70 |
| H* | 22.51 | 25.60 | 23.61 | 5.17 | 0.59 |
| pH | 6.1 | 6.04 | 6.19 | 0.522 | 0.885 |
| Drip loss (%) | 1.97 | 1.78 | 1.56 | 0.44 | 0.294 |
| Cooking loss (%) | 0.29 | 0.31 | 0.26 | 0.04 | 0.126 |
| Protein (g/100 g) | 23.84 | 23.59 | 23.91 | 1.11 | 0.875 |
| Fat (g/100 g) | 1.35 b | 1.76 a | 1.82 a | 0.27 | 0.047 |
| Item (mg/100 g Fresh Meat) | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| C | FD1 | FD2 | |||
| C14:0 | 17.64 | 23.00 | 24.99 | 8.11 | 0.297 |
| C15:0 | 6.85 | 7.34 | 8.86 | 1.66 | 0.111 |
| C16:0 | 248.47 b | 327.77 a | 336.94 a | 59.86 | 0.042 |
| C17:0 | 14.39 | 19.11 | 19.06 | 4.64 | 0.164 |
| C18:0 | 236.01 | 331.83 | 357.42 | 86.44 | 0.065 |
| C20:0 | 2.86 b | 3.35 ab | 4.41 a | 0.97 | 0.041 |
| C21:0 | 1.71 | 1.89 | 1.39 | 0.53 | 0.285 |
| C22:0 | 0.08 | 0.07 | 0.07 | 0.01 | 0.752 |
| C23:0 | 1.01 | 1.11 | 1.16 | 0.19 | 0.386 |
| SFA | 529.03 b | 715.48 a | 754.29 a | 111.74 | 0.007 |
| C15:1 | 82.88 | 90.44 | 86.85 | 17.87 | 0.768 |
| C16:1 | 20.08 b | 27.86 a,b | 36.46 a | 7.12 | 0.004 |
| C18:1 n-9 | 213.31 | 222.08 | 286.45 | 91.73 | 0.347 |
| C20:1 | 1.88 | 1.90 | 1.98 | 0.64 | 0.962 |
| C22:1 | 1.60 | 1.63 | 2.02 | 0.45 | 0.230 |
| MUFA | 319.75 | 343.92 | 413.76 | 104.80 | 0.301 |
| C18:2 n-6 | 165.56 | 176.09 | 160.43 | 35.64 | 0.744 |
| C18:3 n-6 | 1.29 | 1.54 | 1.20 | 0.28 | 0.128 |
| C18:3 n-3 (ALA) | 5.31 b | 6.33 a | 6.82 a | 0.74 | 0.009 |
| C20:2 n-6 | 5.09 | 5.02 | 5.68 | 1.26 | 0.614 |
| C20:3 n-6 | 13.39 | 11.86 | 11.16 | 2.31 | 0.261 |
| C20:4 n-6 | 73.76 | 74.48 | 85.72 | 9.91 | 0.096 |
| C20:5 n-3 (EPA) | 3.46 c | 12.34 b | 18.87 a | 2.44 | <0.001 |
| C22:6 n-3 (DHA) | 7.33 c | 60.30 b | 80.19 a | 5.24 | <0.001 |
| PUFA | 275.19 b | 351.51 a | 370.09 a | 36.86 | 0.001 |
| n-6 PUFA | 259.10 | 268.98 | 264.20 | 36.54 | 0.897 |
| n-3 PUFA | 16.10 c | 78.97 b | 105.89 a | 6.19 | <0.001 |
| EPA + DHA | 10.79 c | 72.64 b | 99.06 a | 6.28 | <0.001 |
| n-6/n-3 | 16.15 a | 3.43 b | 2.52 b | 1.13 | <0.001 |
| Number | Compound | CAS# | Formula | MW | RI | Rt [s] | Dt [ms] |
|---|---|---|---|---|---|---|---|
| 1 | 2-Pentylfuran | C3777693 | C9H14O | 138.2 | 995.0 | 588.195 | 1.25804 |
| 2 | Benzaldehyde | C100527 | C7H6O | 106.1 | 958.7 | 511.489 | 1.15827 |
| 3 | n-Nonanal | C124196 | C9H18O | 142.2 | 1103.3 | 790.923 | 1.47531 |
| 4 | Octanal | C124130 | C8H16O | 128.2 | 1011.5 | 616.193 | 1.40526 |
| 5 | Methional | C3268493 | C4H8OS | 104.2 | 916.8 | 435.272 | 1.08542 |
| 6 | Heptanal (Monomer) | C111717 | C7H14O | 114.2 | 900.6 | 409.009 | 1.33262 |
| 7 | Heptanal (Dimer) | C111717 | C7H14O | 114.2 | 900.0 | 408.054 | 1.7041 |
| 8 | 2-heptanone (Monomer) | C110430 | C7H14O | 114.2 | 892.3 | 396.116 | 1.26633 |
| 9 | 2-Heptanone (Dimer) | C110430 | C7H14O | 114.2 | 891.9 | 395.639 | 1.64334 |
| 10 | 1-Hexanol (Monomer) | C111273 | C6H14O | 102.2 | 871.8 | 368.421 | 1.32986 |
| 11 | 1-Hexanol (Dimer) | C111273 | C6H14O | 102.2 | 872.1 | 368.898 | 1.64472 |
| 12 | Hexanal (Monomer) | C66251 | C6H12O | 100.2 | 792.4 | 278.226 | 1.25507 |
| 13 | Hexanal (Dimer) | C66251 | C6H12O | 100.2 | 791.8 | 277.602 | 1.57443 |
| 14 | 1-Pentanol (Monomer) | C71410 | C5H12O | 88.1 | 764.5 | 251.354 | 1.24767 |
| 15 | 1-Pentanol (Dimer) | C71410 | C5H12O | 88.1 | 762.4 | 249.479 | 1.51031 |
| 16 | 2-Hexanone | C591786 | C6H12O | 100.2 | 783.7 | 269.79 | 1.18602 |
| 17 | Acetoin (Monomer) | C513860 | C4H8O2 | 88.1 | 716.1 | 210.419 | 1.05532 |
| 18 | Acetoin (Dimer) | C513860 | C4H8O2 | 88.1 | 713.6 | 208.544 | 1.34015 |
| 19 | 1-Butanol (Monomer) | C71363 | C4H10O | 74.1 | 663.8 | 177.922 | 1.18356 |
| 20 | 1-Butanol (Dimer) | C71363 | C4H10O | 74.1 | 659.9 | 176.047 | 1.37591 |
| 21 | 2-Butanone (Monomer) | C78933 | C4H8O | 72.1 | 581.6 | 142.612 | 1.06395 |
| 22 | 2-Butanone (Dimer) | C78933 | C4H8O | 72.1 | 588.9 | 145.424 | 1.25137 |
| 23 | 1-Propanol | C71238 | C3H8O | 60.1 | 560.7 | 134.8 | 1.11204 |
| 24 | Acetone | C67641 | C3H6O | 58.1 | 489.8 | 111.364 | 1.1256 |
| 25 | 2-Propanol | C67630 | C3H8O | 60.1 | 501.1 | 114.802 | 1.18232 |
| 26 | Ethanol | C64175 | C2H6O | 46.1 | 458.3 | 102.303 | 1.04546 |
| 27 | 2-Methylpropanol | C78831 | C4H10O | 74.1 | 620.3 | 158.236 | 1.17492 |
| 28 | 2-Pentanone (Monomer) | C107879 | C5H10O | 86.1 | 685.3 | 188.546 | 1.12067 |
| 29 | 2-Pentanone (Dimer) | C107879 | C5H10O | 86.1 | 685.8 | 188.78 | 1.37521 |
| 30 | 3-Methylbutanal | C590863 | C5H10O | 86.1 | 652.1 | 172.421 | 1.40027 |
| 31 | 1-Octen-3-ol (Monomer) | C3391864 | C8H16O | 128.2 | 985.9 | 567.857 | 1.16379 |
| 32 | 1-Octen-3-ol (Dimer) | C3391864 | C8H16O | 128.2 | 984.8 | 565.584 | 1.61059 |
| 33 | 1-Octen-3-ol (Trimer) | C3391864 | C8H16O | 128.2 | 987.3 | 571.04 | 1.73824 |
| 34 | Pentanal (Monomer) | C110623 | C5H10O | 86.1 | 695.9 | 195.421 | 1.18443 |
| 35 | Pentanal (Dimer) | C110623 | C5H10O | 86.1 | 694.3 | 194.228 | 1.42808 |
| 36 | 2-Ethylhexanol | C104767 | C8H18O | 130.2 | 1043.5 | 672.122 | 1.40894 |
| Item | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| C | FD1 | FD2 | |||
| Initial juiciness | 5.13 b | 6.19 a | 6.21 a | 0.56 | 0.006 |
| Sustained juiciness | 4.94 b | 5.94 a | 6.10 a | 0.63 | 0.012 |
| Flavor intensity | 5.48 b | 6.33 a | 6.44 a | 0.45 | 0.004 |
| Off-flavor intensity | 2.17 | 2.00 | 2.31 | 0.28 | 0.187 |
| Initial tenderness | 5.52 b | 6.33 a | 6.50 a | 0.60 | 0.029 |
| Sustained tenderness | 5.63 b | 6.35 a | 6.42 a | 0.56 | 0.049 |
| Residue | 3.68 a | 3.21 b | 3.10 b | 0.34 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Zhang, S.; Sun, B.; Xie, P.; Liu, X.; Chang, L.; Lu, F.; Zhang, S. Dietary Supplementation with Microalgae (Schizochytrium sp.) Improves the Antioxidant Status, Fatty Acids Profiles and Volatile Compounds of Beef. Animals 2021, 11, 3517. https://doi.org/10.3390/ani11123517
Xu C, Zhang S, Sun B, Xie P, Liu X, Chang L, Lu F, Zhang S. Dietary Supplementation with Microalgae (Schizochytrium sp.) Improves the Antioxidant Status, Fatty Acids Profiles and Volatile Compounds of Beef. Animals. 2021; 11(12):3517. https://doi.org/10.3390/ani11123517
Chicago/Turabian StyleXu, Chenchen, Shou Zhang, Baozhong Sun, Peng Xie, Xiaochang Liu, Lan Chang, Fushan Lu, and Songshan Zhang. 2021. "Dietary Supplementation with Microalgae (Schizochytrium sp.) Improves the Antioxidant Status, Fatty Acids Profiles and Volatile Compounds of Beef" Animals 11, no. 12: 3517. https://doi.org/10.3390/ani11123517
APA StyleXu, C., Zhang, S., Sun, B., Xie, P., Liu, X., Chang, L., Lu, F., & Zhang, S. (2021). Dietary Supplementation with Microalgae (Schizochytrium sp.) Improves the Antioxidant Status, Fatty Acids Profiles and Volatile Compounds of Beef. Animals, 11(12), 3517. https://doi.org/10.3390/ani11123517