
Fruit Size and Structure of Zoochorous Trees: Identifying Drivers for the Foraging Preferences of Fruit-Eating Birds in a Mexican Successional Dry Forest

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
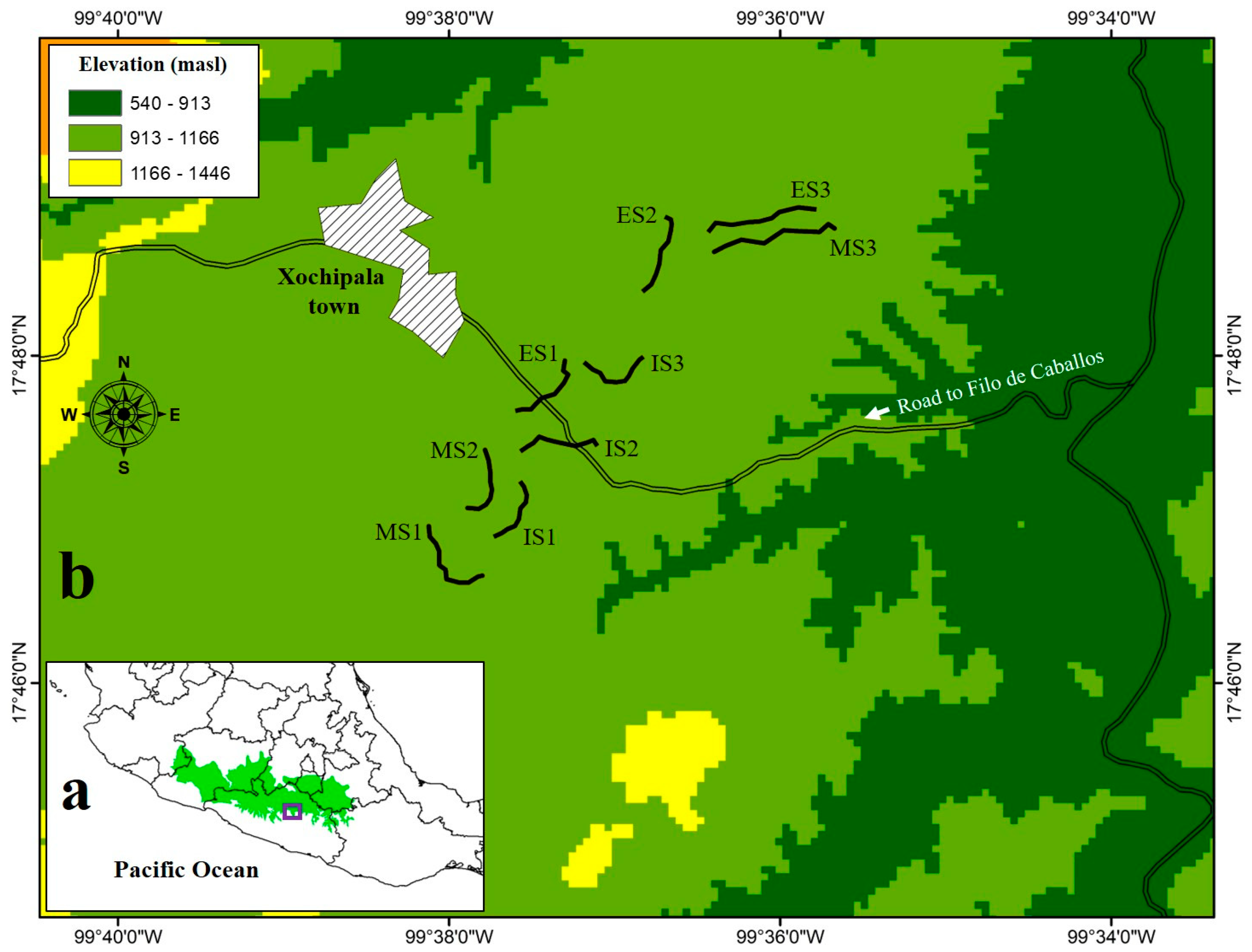
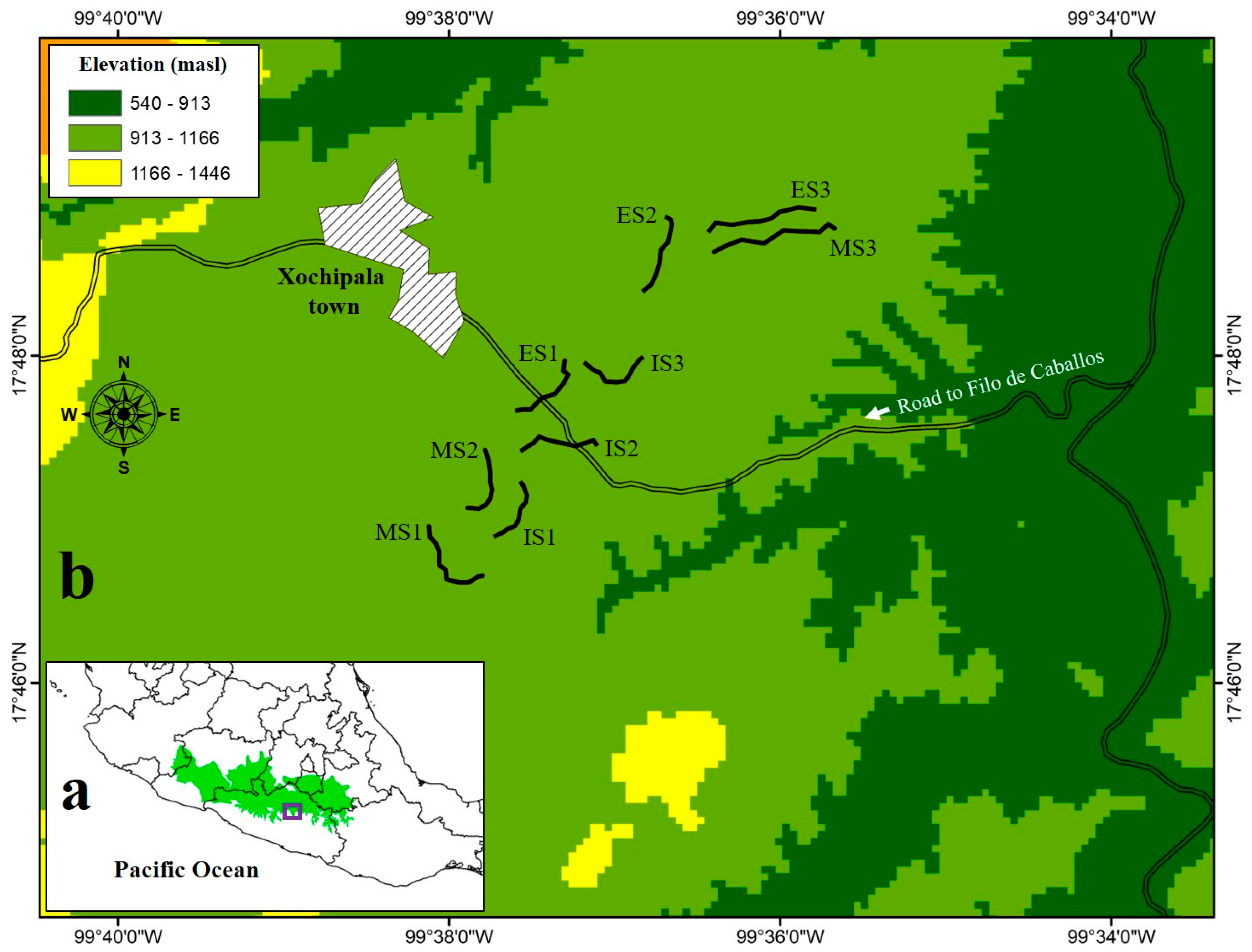
2.1. Description of the Study Area and Sampling Sites
- (1)
- Early stages (~15 years, ES) are composed of vegetation that arose spontaneously in areas used for livestock and temporal agriculture. Currently, these sites continue to be subjected to species-selective logging; hence, there are scattered trees in rocky soils unsuitable for large-scale agriculture [47]. Some representative species from these sites are Gliricidia sepium, Acacia cochliacantha, Ipomoea pauciflora, Ceiba aesculifolia, and Bursera longipes.
- (2)
- Intermediate stages (~30–35 years, IS) represent a transition zone between the early and mature forests. These sites were also used for livestock and temporal agriculture for corn and bean production. Nonetheless, to a large extent, they have developed structural and floristic elements from the original vegetation. The dominant species at this stage are Lysiloma tergemina, Exostema caribaeum, Bursera longipes, B. morelensis, and Pterocarpus acapulcensis. The orography of these sites is not very rugged, with slopes of 15° to 30°.
- (3)
- Mature stages (>50 years, MS) are sites characterized by the presence of typical plant species of mature forests, such as Bursera spp., Desmanthus balsensis, Mimosa polyantha, M. goldmanii, and Pseudosmodingium perniciosum. These sites were subjected to a clear-cut-and-burn system for agricultural and/or ranching activities, but their abandonment permitted the regeneration of the vegetation. The orography of these sites is rugged, with slopes >30° [47].
2.2. Fruiting Phenology
2.3. Foliar Cover, Foliage Height Diversity, and Fruit Size of Zoochorous Trees
2.4. Foraging Observations in Zoochorous Trees
2.5. Data Analysis
3. Results
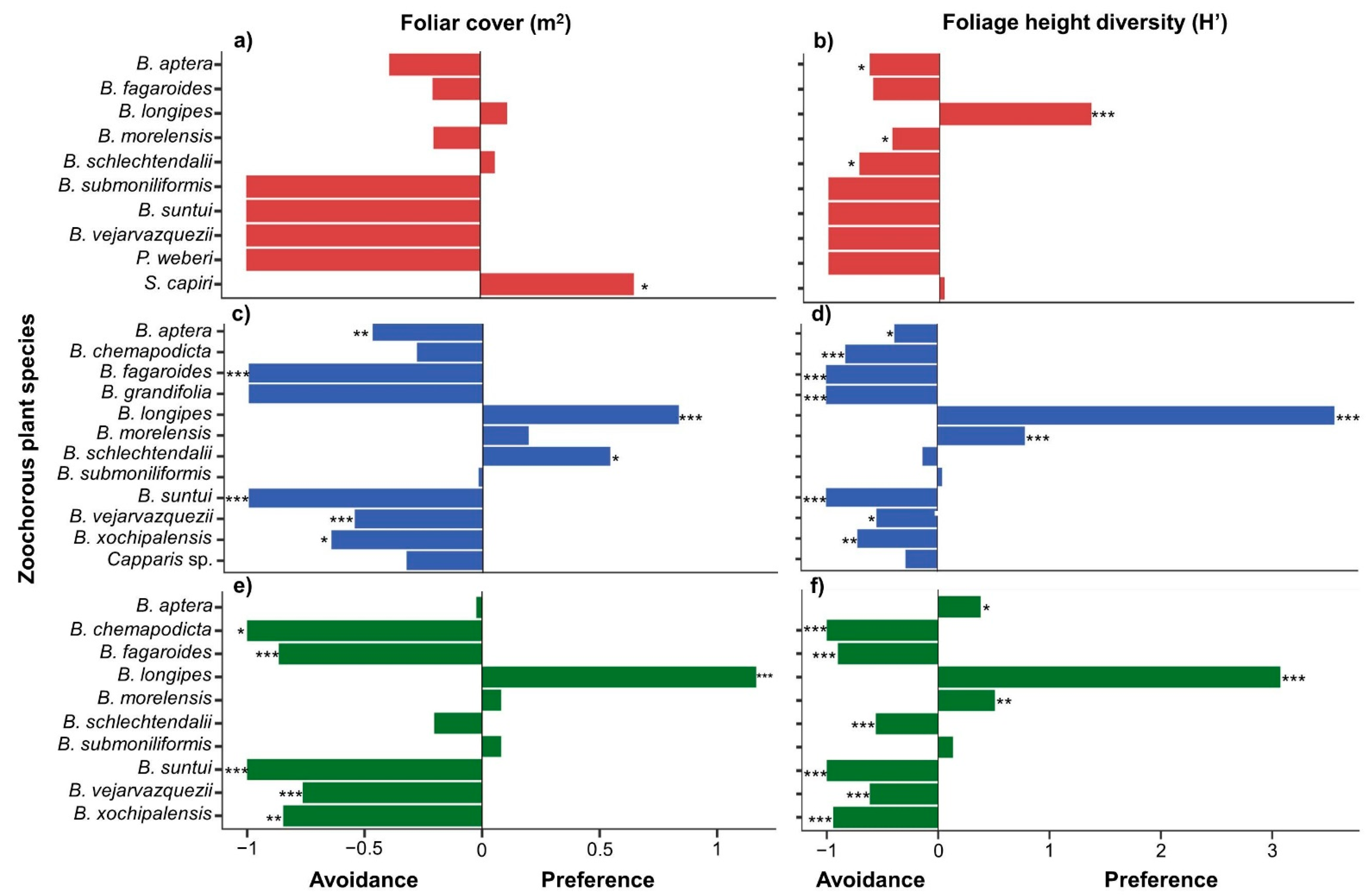
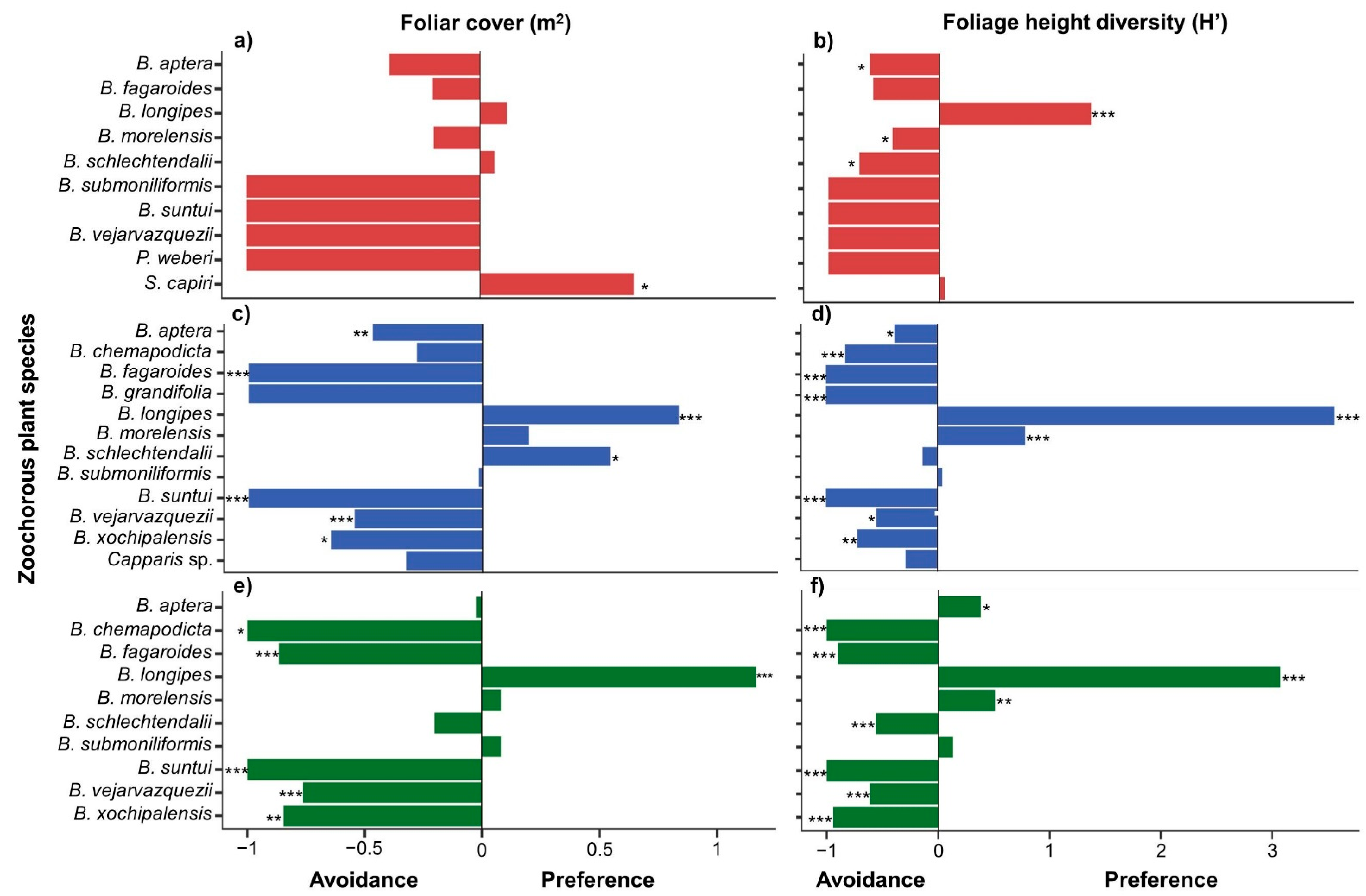
3.1. Foliar Cover, Foliage Height Diversity, and Fruit Size of Zoochorous Trees
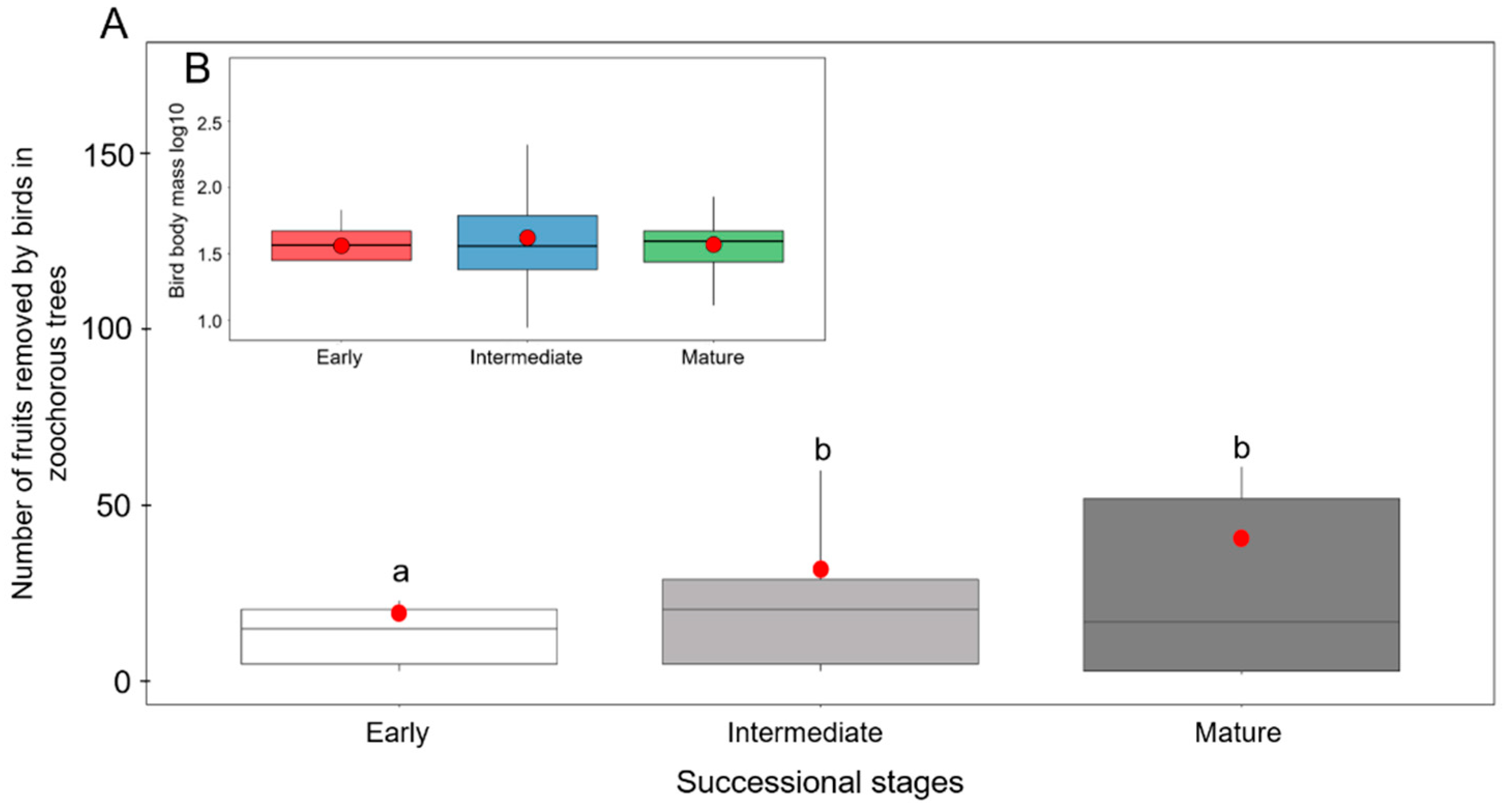
3.2. Fruit Removal in Zoochorous Trees by Successional Stage
3.3. Foraging Preferences in Zoochorous Trees by Successional Stage
3.4. Effects of Fruit Size and Structural Variables of Zoochorous Trees on Fruit Removal
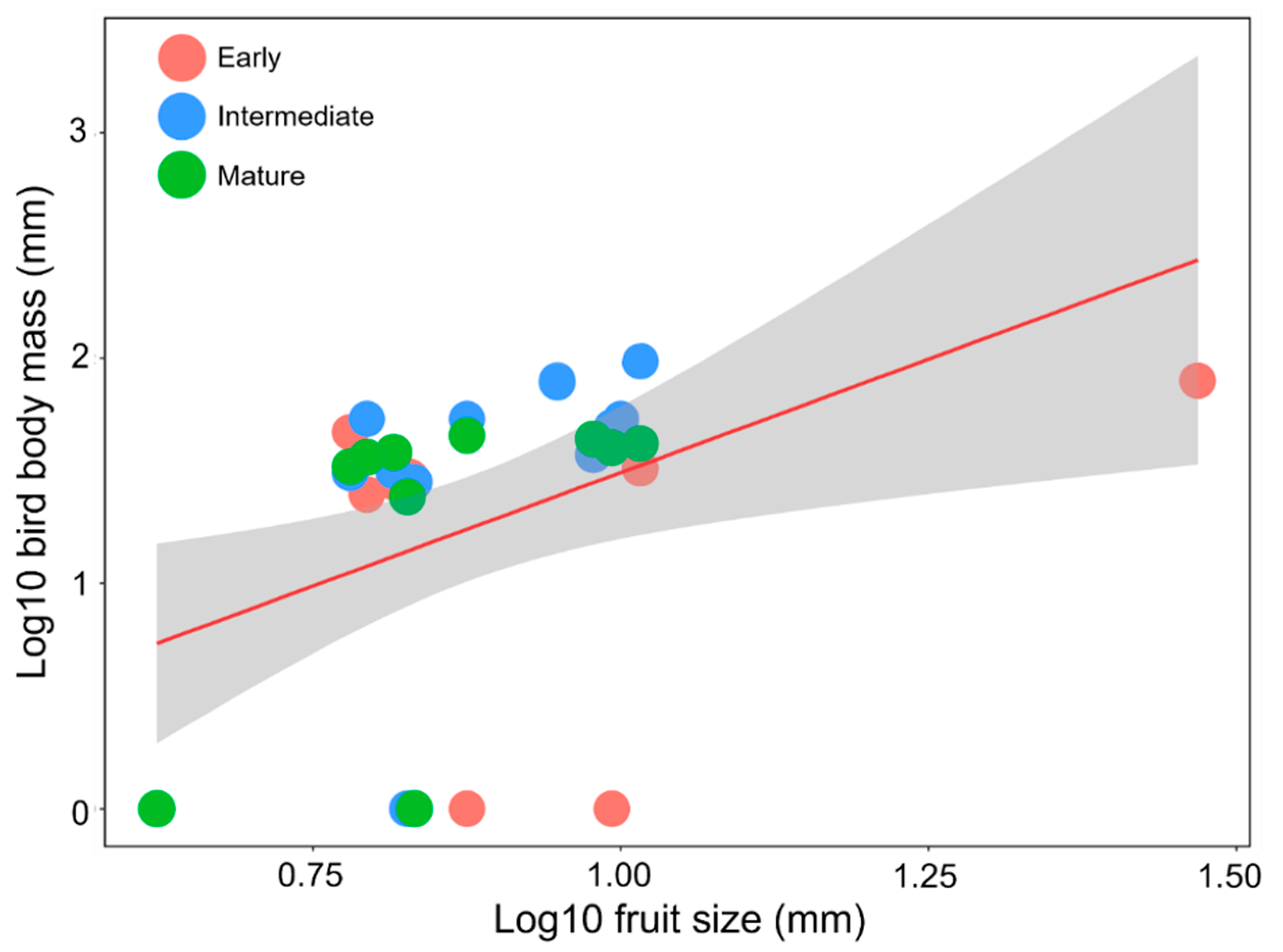
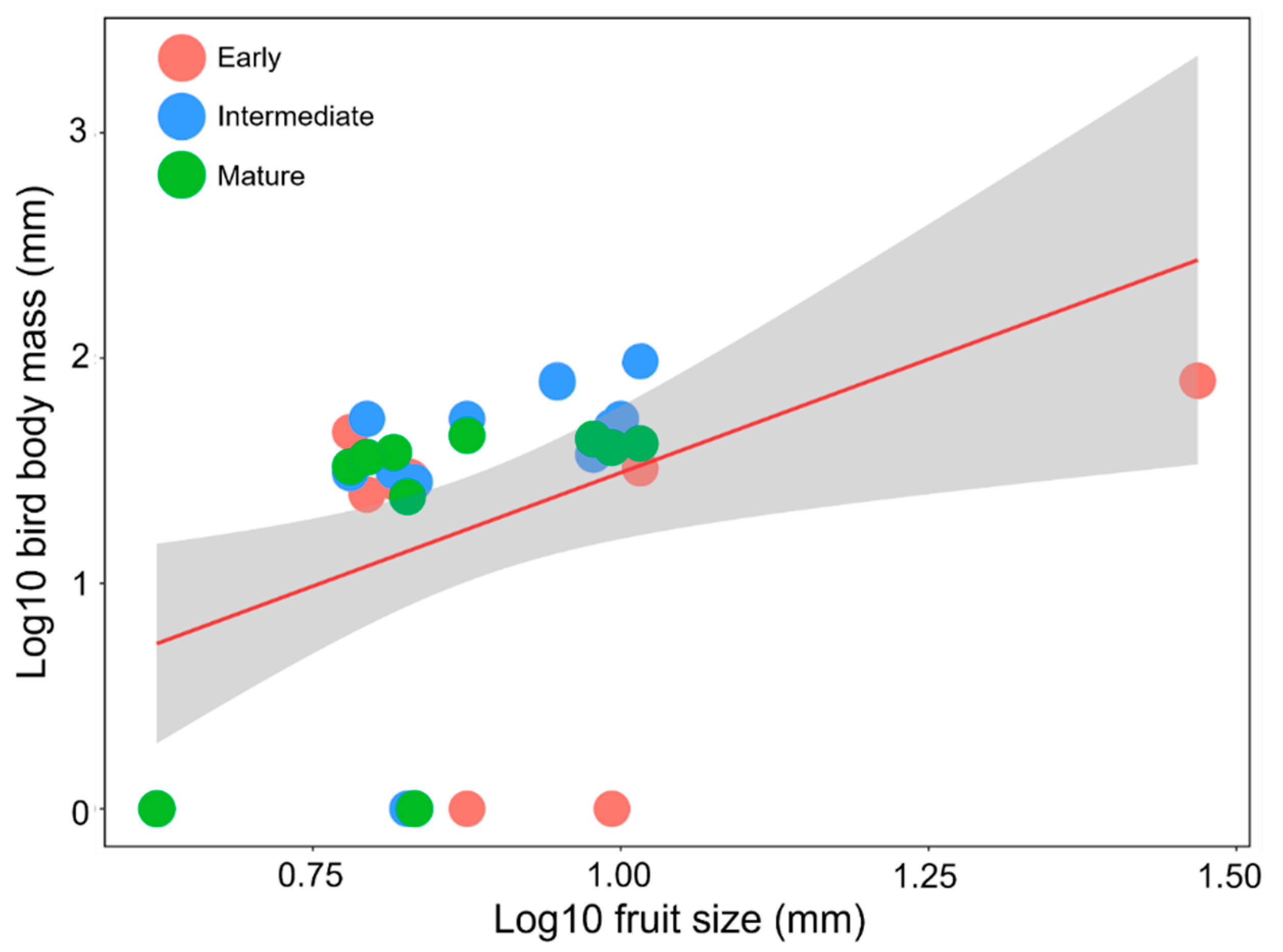
3.5. Relationship between Bird Body Mass and Fruit Size of Zoochorous Trees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portillo-Quintero, C.A.; Sánchez-Azofeifa, G.A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Quesada, M.; Rosas, F.; Aguilar, R.; Ashworth, L.; Rosas-Guerrero, V.M.; Sayago, R.; Lobo, J.A.; Herrerías-Diego, Y.; Sánchez-Montoya, G. Human impacts on pollination, reproduction, and breeding systems in tropical forest plants. In Seasonally Dry Tropical Forests. Ecology and Conservation; Dirzo, R., Young, H., Mooney, H.A., Ceballos, G., Eds.; Island Press: Washington, DC, USA, 2011; pp. 173–194. [Google Scholar] [CrossRef]
- Balvanera, P.; Islas, Á.; Aguirre, E.; Quijas, S. Las selvas secas. Ciencias 2000, 37, 19–24. [Google Scholar]
- Stoner, K.E.; Sanchez-Azofeita, G.A. Ecology and regeneration of tropical dry forests in the Americas: Implications for management. For. Ecol. Manag. 2009, 258, 903–906. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Schondube, J.E. Gray vs. green urbanization: Relative importance of urban features for urban bird communities. Basic Appl. Ecol. 2011, 12, 372–381. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Arizmendi, M.C.; Eguiarte, L.E.; Corcuera, P. Distribution of the community of frugivorous birds along a successional gradient in dry tropical forest in southwestern Mexico. J. Trop. Ecol. 2015, 31, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Almazán-Núñez, R.C.; Alvarez-Alvarez, E.A.; Pineda-López, R.; Corcuera, P. Seasonal variation in bird assemblage composition in a dry forest of Southwestern Mexico. Ornitol. Neotrop. 2018, 29, 215–224. [Google Scholar]
- Battistelli, G.; Chisci, L.; Selvi, D.A. Distributed Kalman filter with event-triggered communication and guaranteed stability. Automatica 2018, 93, 75–82. [Google Scholar] [CrossRef]
- Şekercioğlu, Ç.H.; Daily, G.C.; Ehrlich, P.R. Ecosystem consequences of bird declines. Proc. Natl. Acad. Sci. USA 2004, 101, 18042–18047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlo, T.A.; Aukema, J.E.; Morales, J.M. Plant-frugivore interactions as spatially explicit networks: Integrating frugivore foraging with fruiting plant spatial patterns. In Seed Dispersal: Theory and its Application in a Changing World; Dennis, A., Schupp, E., Wescott, D., Eds.; CABI: Oxon, UK, 2007; pp. 369–390. [Google Scholar]
- Godínez-Álvarez, H.O.; Valiente-Banuet, A.; Rojas-Martínez, A. The role of seed dispersers in the population dynamics of the columnar cactus Neobuxbaumia tetetzo. Ecology 2002, 83, 2617–2629. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Eguiarte, L.E.; Arizmendi, M.C.; Corcuera, P. Myiarchus flycatchers are the primary seed dispersers of Bursera longipes in a Mexican dry forest. PeerJ 2016, 4, e2126. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Pulido, R.; Rico-Gray, V. Seed dispersal of Bursera fagaroides (Burseraceae): The effect of linking environmental factors. Southwest. Nat. 2006, 51, 11–21. [Google Scholar] [CrossRef]
- Ramos-Ordoñez, M.F.; Arizmendi, M.C. Parthenocarpy, attractiveness and seed predation by birds in Bursera morelensis. J. Arid. Environ. 2011, 75, 757–762. [Google Scholar] [CrossRef]
- Contreras-González, A.M.; Arizmendi, M.C. Pre-dispersal seed predation of the columnar cactus (Neobuxbaumia tetetzo, Cactaceae) by birds in central Mexico. Ornitol. Neotrop. 2014, 25, 373–387. [Google Scholar]
- Van der Pijl, L. Principles of Dispersal of Higher Plants; Springer: Berlin, Germany, 1982. [Google Scholar]
- Rzedowski, J.; Medina, R.; Calderón de Rzedowski, G. Inventario del conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Bot. Mex. 2005, 70, 85–111. [Google Scholar] [CrossRef]
- Mokotjomela, T.M.; Musil, C.F.; Esler, K.J. Do frugivorous birds concentrate their foraging activities on those alien plants with the most abundant and nutritious fruits in the South African Mediterranean-climate region? Plant Ecol. 2013, 214, 49–59. [Google Scholar] [CrossRef]
- Traveset, A.; Richardson, D.M. Mutualistic interactions and biological invasions. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Foster, M.S. Factors influencing bird foraging preferences among conspecific fruit trees. Condor 1990, 92, 844–854. [Google Scholar] [CrossRef]
- Ortiz-Pulido, R.; Albores-Barajas, Y.V.; Díaz, S.A. Fruit removal efficiency and success: Influence of crop size in a neotropical treelet. Plant Ecol. 2007, 189, 147–154. [Google Scholar] [CrossRef]
- Palacio, F.X.; Ordano, M. The strength and drivers of bird-mediated selection on fruit crop size: A meta-analysis. Front. Ecol. Evol. 2018, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Fuzessy, L.F.; Janson, C.; Silveira, F.A. Effects of seed size and frugivory degree on dispersal by Neotropical frugivores. Acta Oecol. 2018, 93, 41–47. [Google Scholar] [CrossRef]
- Jordano, P. Fruits and frugivory. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CABI: Wallingford, UK, 2000; p. 410. [Google Scholar]
- Lim, J.Y.; Svenning, J.S.; Göldel, B.; Faurby, S.; Kissling, D. Frugivore-fruit size relationships between palms and mammals reveal past and future defaunation impacts. Nat. Commun. 2020, 11, 4904. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, P.R.; Galetti, M.; Jordano, P. Seed dispersal anachronisms: Rethinking the fruits extinct megafauna ate. PLoS ONE 2008, 3, e1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, L.; Oostra, V.; Nijman, V.; Cleef, A.M.; Kappelle, M. Tolerance of frugivorous birds to habitat disturbance in a tropical cloud forest. Biol. Conserv. 2008, 141, 860–871. [Google Scholar] [CrossRef]
- Mahoney, M.; Browne, L.; Díaz-Martin, Z.; Olivo, J.; Cabrera, J.; González, M.; Hazlehurst, J.; Karubian, J. Fruit removal by large avian frugivores varies in relation to habitat quality in continuous neotropical rainforest. Ornitol. Neotrop. 2018, 29, 247–254. [Google Scholar]
- Galetti, M.; Guevara, R.; Côrtes, M.; Fadini, R.; Von Matter, S.; Leite, A.; Labecca, F.; Ribeiro, T.; Carvalho, C.; Collevatti, R.; et al. Functional extinction of birds drives rapid evolutionary changes in seed size. Science 2013, 340, 1086–1090. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Chapman, C.A.; Chapman, L.J.; Wangham, R.; Hunt, K.; Gebo, D.; Gardner, L. Estimators of fruit abundance of tropical trees. Biotropica 1992, 24, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.A.; Wrangham, R.; Chapman, L.J. Indices of habitat-wide fruit abundance in tropical forest. Biotropica 1994, 26, 160–171. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Maliakal, S.K.; Denslow, J.S. Changes in vegetation structure and composition along a tropical forest chronosequence: Implications for wildlife. For. Ecol. Manag. 2003, 182, 139–151. [Google Scholar] [CrossRef]
- Gomes, V.S.M.; Loiselle, B.A.; Alves, M.A. Birds foraging for fruits and insects in shrubby resting vegetation, southeastern Brazil. Biota Neotrop. 2008, 8, 21–31. [Google Scholar] [CrossRef]
- Naniwadekar, R.; Mishra, C.; Datta, A. Fruit resource tracking by hornbill species at multiple scales in a tropical forest in India. J. Trop. Ecol. 2015, 31, 477–490. [Google Scholar] [CrossRef]
- Gopal, A.; Mudappa, D.; Raman, T.S.; Naniwadekar, R. Forest cover and fruit crop size differentially influence frugivory of select rainforest tree species in Western Ghats, India. Biotropica 2020, 52, 871–883. [Google Scholar] [CrossRef]
- Dinerstein, E. The foliage-as-fruit hypothesis and the feeding behavior of south Asian ungulates. Biotropica 1989, 21, 214–218. [Google Scholar] [CrossRef]
- Minor, D.; Kobe, R. Fruit production is influenced by tree size and size-asymmetric crowding in a wet tropical forest. Ecol. Evol. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R. The relation of foliage complexity to ecological diversity of three Amazonian bird communities. Condor 1975, 77, 453–466. [Google Scholar] [CrossRef]
- Jayson, E.A.; Mathew, D.N. Vertical stratification and its relation to foliage in tropical forest birds in Western Ghats (India). Acta Ornitholog. 2003, 38, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Basu, P. Avian frugivory and seed dispersal of a large fruited tree in an Indian moist deciduous forest. Acta Oecol. 2015, 65, 32–40. [Google Scholar] [CrossRef]
- Loiselle, B.A.; Blake, J.G. Population variation in a tropical bird community. BioScience 1992, 42, 838–845. [Google Scholar] [CrossRef]
- Tinoco, B. Estacionalidad de la comunidad de aves en un bosque deciduo tumbesino en el sur occidente de Ecuador. Ornitol. Neotrop. 2009, 20, 157–170. [Google Scholar]
- Pérez-Villafaña, M.G.; Valiente-Banuet, A. Effectiveness of dispersal of an ornithocorous cactus Myrtillocactus geometrizans (Cactaceae) in a patchy environment. Open Biol. J. 2009, 2, 101–113. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Mariano-Rendón, A.; Rodríguez-Godínez, R.; Méndez-Bahena, A.; Pineda-López, R. Las Aves Frugívoras y su Papel en la Restauración Pasiva del Bosque Tropical Caducifolio del sur de México: Un Caso de Estudio con la Cactácea Pachycereus weberi. In Manejo y Conservación de Fauna en Ambientes Antropizados; Mercado Silva, N., Ek del Val, G., Eds.; Fondo Editorial Universidad Autónoma de Querétaro: Querétaro, Mexico, 2021; pp. 61–83. [Google Scholar]
- Quesada, M.; Sánchez-Azofeita, G.A.; Álvarez-Añorve, M.; Stoner, K.E.; Avila-Cabadilla, L.; Calvo-Alvarado, J.; Castillo, A.; Espírito-Santo, M.M.; Fagundes, M.; Fernandes, G.W.; et al. Succession and management of tropical dry forests in the Americas: Review and new perspectives. For. Ecol. Manag. 2009, 258, 1014–1024. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Arizmendi, M.C.; Eguiarte, L.E.; Corcuera, P. Changes in composition, diversity and structure of woody plants in successional stages of tropical dry forest in southwest Mexico. Rev. Mex. Biodivers. 2012, 83, 1096–1109. [Google Scholar] [CrossRef] [Green Version]
- Meza, L.; López, J.L. Vegetación y Mesoclima de Guerrero. In Estudios Florísticos en Guerrero; Pérez, N.D., Fonseca, R.M., Eds.; Facultad de Ciencias, Universidad Autonoma de Mexico: Mexico City, Mexico, 1997; pp. 1–53. [Google Scholar]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen, 4th ed.; Universidad Autonoma de Mexico: Mexico City, Mexico, 1988. [Google Scholar]
- Muller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Montaña, C.; Ezcurra, E. Simple instrument for quick measurement of crown projections. J. For. 1980, 78, 699. [Google Scholar] [CrossRef]
- Hutto, R.L.; Pletschet, S.M.; Hendricks, P. A fixed-radius point count method for nonbreeding and breeding season use. Auk 1986, 103, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, S.T.; Scott, J.M.; Nussbaum, R.A. A variable circular-plot method for estimating bird numbers. Condor 1980, 82, 309–313. [Google Scholar] [CrossRef]
- Traveset, A. Influence of type of avian frugivory on the fitness of Pistacia terebinthus. Evol. Ecol. 1994, 8, 618–627. [Google Scholar] [CrossRef]
- Jordano, P.; Schupp, E. Seed disperser effectiveness: The quantity component and patterns of seed rain for Prunus mahaleb. Ecol. Monogr. 2000, 70, 591–615. [Google Scholar] [CrossRef]
- Peck, K.M. Tree species preferences shown by foraging birds in forest plantation in northern England. Biol. Conserv. 1989, 48, 41–57. [Google Scholar] [CrossRef]
- Cody, M.L. An Introduction to Habitat Selection in Birds. In Habitat Selection in Birds; Cody, M.L., Ed.; Academic Press: Orlando, FL, USA, 1981; pp. 4–56. [Google Scholar]
- Hasui, É.; Gomes, V.S.D.M.; Silva, W.R. Effects of vegetation traits on habitat preferences of frugivorous birds in Atlantic rain forest. Biotropica 2007, 39, 502–509. [Google Scholar] [CrossRef]
- Wilman, H.; Belmaker, J.; Simpson, J.; De la Rosa, C.; Rivadeneira, M.; Jetz, W. Eltontraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org (accessed on 15 August 2021).
- Kissling, W.D.; Rahbek, C.; Böhning-Gaese, K. Food plant diversity as broad-scale determinant of avian frugivore richness. Proc. R. Soc. B Biol. Sci. 2007, 274, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Githiru, M.; Lens, L.; Bennur, L.A.; Ogol, C.P. Effects of site and fruit size on the composition of avian frugivore assemblages in a fragmented Afrotropical forest. Oikos 2002, 96, 320–330. [Google Scholar] [CrossRef]
- Morante-Filho, J.C.; Arroyo-Rodríguez, V.; Faria, D. Patterns and predictors of β-diversity in the fragmented Brazilian Atlantic forest: A multiscale analysis of forest specialist and generalist birds. J. Anim. Ecol. 2016, 85, 240–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leso, P.; Kropil, R. A comparison of three different approaches for the classification of bird foraging guilds: An effect of leaf phenophase. Folia Zoölogica 2007, 56, 51–70. [Google Scholar]
- Cordeiro, N.J.; Howe, H.F. Forest fragmentation severs mutualism between seed dispersers and an endemic African tree. Proc. Natl. Acad. Sci. USA 2003, 100, 14052–14056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa-Esquivel, E.; Puebla-Olivares, F.; Godínez-Álvarez, H.; Núñez-Farfán, J. Seed dispersal effectiveness by understory birds on Dendropanax arboreus in a fragmented landscape. Biodivers. Conserv. 2009, 18, 3357–3365. [Google Scholar] [CrossRef]
- Viswanathan, A.; Naniwadekar, R.; Datta, A. Seed dispersal by avian frugivores: Non-random heterogeneity at fine scales. Biotropica 2015, 47, 77–84. [Google Scholar] [CrossRef]
- Herrera, J.M.; García, D. The role of remnant trees in seed dispersal through the matrix: Being alone is not always so sad. Biol. Conserv. 2009, 142, 149–158. [Google Scholar] [CrossRef]
- Alvarez-Alvarez, E.A.; Corcuera, P.; Almazán-Núñez, R.C. Spatiotemporal variation in the structure and diet types of bird assemblages in tropical dry forest in southwestern Mexico. Wilson J. Ornithol. 2018, 130, 457–469. [Google Scholar] [CrossRef]
- Kalacska, M.; Sánchez-Azofeita, G.A.; Calvo-Alvarado, J.C.; Rivard, B.; Quesada, M. Effects of season and successional stage on leaf area index and spectral vegetation indices in three Mesoamerican tropical dry forest. Biotropica 2005, 37, 486–496. [Google Scholar] [CrossRef]
- Ruiz, J.; Fandino, M.C.; Chazdon, R.L. Vegetation structure, composition, and species richness across a 56-year chronosequence of dry tropical forest on Providencia island, Colombia. Biotropica 2005, 37, 520–530. [Google Scholar] [CrossRef]
- Vidal, M.M.; Hasui, E.; Pizo, M.A.; Tamashiro, J.Y.; Silva, W.R.; Guimarães, P.R., Jr. Frugivores at higher risk of extinction are the key elements of a mutualistic network. Ecology 2014, 95, 3440–3447. [Google Scholar] [CrossRef] [Green Version]
- Godínez-Álvarez, H.; Ríos-Casanova, L.; Peco, B. Are large frugivorous birds better seed dispersers than medium- and small-sized ones? Effect of body mass on seed dispersal effectiveness. Ecol. Evol. 2020, 10, 6136–6143. [Google Scholar] [CrossRef]
- Pessoa, M.S.; Rocha-Santos, L.; Talora, D.C.; Faria, D.; Mariano-Neto, E.; Hambuckers, A.; Cazetta, E. Fruit biomass availability along a forest cover gradient. Biotropica 2017, 49, 45–55. [Google Scholar] [CrossRef]
- Jordano, P.; García, C.; Godoy, J.A.; García-Cataño, J.L. Differential contribution of frugivores to complex seed dispersal patterns. Proc. Nat. Acad. Sci. USA 2007, 104, 3278–3282. [Google Scholar] [CrossRef] [Green Version]
- Flörchinger, M.; Braun, J.; Böhning-Gaese, K.; Schaefer, H.M. Fruit size, crop mass, and plant height explain diferential fruit choice of primates and birds. Oecologia 2010, 164, 151–161. [Google Scholar] [CrossRef]
- Rodríguez-Godínez, R.; Sánchez-González, L.A.; Arizmendi, M.C.; Almazán-Núñez, R.C. Bursera fruit traits as drivers of fruit removal by flycatchers. Acta Oecol. 2021, in press. [Google Scholar]
- Greenberg, R.; Foster, M.S.; Márquez-Valdelamar, L. The role of the white-eyed vireo in the dispersal of Bursera fruit on the Yucatan Peninsula. J. Trop. Ecol. 1995, 11, 619–639. [Google Scholar] [CrossRef]
- Ramos-Ordoñez, M.F. Dispersión biótica de semillas y caracterización de los frutos de Bursera morelensis en el Valle de Tehuacán, Puebla. Ph.D. Thesis, Universidad Nacional Autónoma de México, Mexico City, Mexico, 2009. [Google Scholar]
- Carranza, E. Sapotaceae. Flora del Bajío y de Regiones Adyacentes. Fascículo 132; Instituto de Ecología A.C. Centro Regional del Bajío: Pátzcuaro, México, 2005; Volume 132, pp. 1–34. [Google Scholar]
- García, E.G.; Di Stefano, J.F. Fenología de árbol Sideroxylon capiri (Sapotaceae) en el bosque tropical seco de Costa Rica. Rev. Biol. Trop. 2005, 53, 5–14. [Google Scholar]
- Jordano, P. Angiosperm fleshy fruits and seed disperser: A comparative analysis of adaptation and constraints in plant-animal interactions. Am. Nat. 1995, 145, 163–191. [Google Scholar] [CrossRef] [Green Version]
- Padilla, F.M.; Pugnaire, F.I. The role of nurse plants in the restoration of degraded environments. Front. Ecol. Environ. 2006, 4, 196–202. [Google Scholar] [CrossRef]
- Bender, I.M.; Kissling, W.D.; Blendinger, P.G.; Böhning-Gaese, K.; Hensen, I.; Kühn, I.; Muñoz, M.C.; Neuschulz, E.L.N.; Nowak, L.; Quitián, M.; et al. Morphological trait matching shapes plant–frugivore networks across the Andes. Ecography 2018, 41, 1910–1919. [Google Scholar] [CrossRef] [Green Version]
- García, D.; Martínez, D.; Herrera, J.M.; Morales, J.M. Functional heterogeneity in a plant-frugivore assemblage enhances seed dispersal resilience to habitat loss. Ecography 2013, 36, 197–208. [Google Scholar] [CrossRef]
- González-Castro, A.; Calviño-Cancela, M.; Nogales, M. Comparing seed dispersal effectiveness by frugivores at the community levels. Ecology 2015, 96, 808–818. [Google Scholar] [CrossRef]
- Guízar, E.; Sánchez, A. Guía para el Reconocimiento de los Principales Árboles del Alto Balsas; Universidad Autónoma Chapingo: Chapingo, Mexico, 1991. [Google Scholar]
- De-Nova, J.A.; Medina, R.; Montero, J.C.; Weeks, A.; Rosell, J.A.; Olson, M.E.; Eguiarte, L.E.; Magallón, S. Insights into the historical construction of species-rich Mesoamerican seasonally dry tropical forests: The diversification of Bursera (Burseraceae, Sapindales). New Phytol. 2012, 193, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Secretaria de Medio Ambiente y Recursos Naturales. Available online: https://backend.aprende.sep.gob.mx/media/uploads/proedit/resources/inventario_estatal_f_c471db15.pdf (accessed on 20 July 2021).
- Chesser, R.T.; Billerman, S.M.; Burns, K.J.; Cicero, C.; Dunn, J.L.; Kratter, A.W.; Lovette, I.J.; Mason, N.A.; Rasmussen, P.C.; Remsen, J.V., Jr.; et al. Check-list of North American birds (online); American Ornithological Society: Chicago, IL, USA, 2020; Available online: http://checklist.americanornithology.org/taxa (accessed on 15 August 2021).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Fruit Size (mm) | Foliar Cover (m2) | Foliage Height Diversity (H’) | ||||
|---|---|---|---|---|---|---|---|
| Early | Intermediate | Mature | Early | Intermediate | Mature | ||
| Bursera aptera | 6.5 | 73.27 | 1241.53 | 1114.4 | 1.04 | 1.84 | 1.84 |
| Bursera chemapodicta | 7.5 | - | 144.25 | 123.11 | - | 1.04 | 1.58 |
| Bursera fagaroides | 6.7 | 37.47 | 409.67 | 463.04 | 0.64 | 1.84 | 1.47 |
| Bursera grandifolia | 8.9 | - | 22.46 | - | - | 1.57 | - |
| Bursera longipes | 10.4 | 468.51 | 2763.64 | 1669.78 | 1.9 | 1.91 | 2.08 |
| Bursera morelensis | 6.2 | 139.85 | 1736.57 | 1179.67 | 1.68 | 2 | 1.98 |
| Bursera schlechtendalii | 6.0 | 21.07 | 581.74 | 446.22 | 0.69 | 1.79 | 1.89 |
| Bursera submoniliformis | 7.5 | 6.83 | 1055 | 819.84 | 0.1 | 1.71 | 1.81 |
| Bursera suntui | 4.2 | 17.17 | 460.39 | 154.23 | 0.1 | 1.77 | 1.55 |
| Bursera vejarvazquezii | 9.8 | 8.71 | 1848.18 | 1410.25 | 0.1 | 2.02 | 2.02 |
| Bursera xochipalensis | 9.5 | - | 489.52 | 272.22 | - | 1.06 | 1.68 |
| Capparis spp. | 10 | - | 61.51 | - | - | 0.1 | - |
| Pachycereus weberi | 41.9 | 11.32 | - | - | 0.1 | - | - |
| Sideroxylon capiri | 29.4 | 81.07 | - | - | 1.11 | - | - |
| Variables | Estimate | SE | Z-Value | p-Value | AIC |
|---|---|---|---|---|---|
| Fixed effects | All fruit-eating birds | ||||
| Foliage height diversity | 3.83 | 0.18 | 20.57 | *** | 672.64 |
| Fruit size | 0.12 | 0.01 | 12.78 | *** | |
| Potential legitimate dispersers | |||||
| Foliage height diversity | 2.65 | 0.17 | 15.62 | *** | 560.58 |
| Fruit size | 0.11 | 0.01 | 10.20 | *** | |
| Seed predators | |||||
| Foliage height diversity | 7.41 | 0.91 | 8.12 | *** | 110.94 |
| Fruit size | 0.19 | 0.05 | 3.97 | ** | |
| Pulp consumers | |||||
| Foliage height diversity | 4.41 | 0.61 | 7.23 | *** | 180.43 |
| Fruit size | 0.09 | 0.03 | 3.19 | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almazán-Núñez, R.C.; Alvarez-Alvarez, E.A.; Sierra-Morales, P.; Rodríguez-Godínez, R. Fruit Size and Structure of Zoochorous Trees: Identifying Drivers for the Foraging Preferences of Fruit-Eating Birds in a Mexican Successional Dry Forest. Animals 2021, 11, 3343. https://doi.org/10.3390/ani11123343
Almazán-Núñez RC, Alvarez-Alvarez EA, Sierra-Morales P, Rodríguez-Godínez R. Fruit Size and Structure of Zoochorous Trees: Identifying Drivers for the Foraging Preferences of Fruit-Eating Birds in a Mexican Successional Dry Forest. Animals. 2021; 11(12):3343. https://doi.org/10.3390/ani11123343
Chicago/Turabian StyleAlmazán-Núñez, R. Carlos, Edson A. Alvarez-Alvarez, Pablo Sierra-Morales, and Rosalba Rodríguez-Godínez. 2021. "Fruit Size and Structure of Zoochorous Trees: Identifying Drivers for the Foraging Preferences of Fruit-Eating Birds in a Mexican Successional Dry Forest" Animals 11, no. 12: 3343. https://doi.org/10.3390/ani11123343
APA StyleAlmazán-Núñez, R. C., Alvarez-Alvarez, E. A., Sierra-Morales, P., & Rodríguez-Godínez, R. (2021). Fruit Size and Structure of Zoochorous Trees: Identifying Drivers for the Foraging Preferences of Fruit-Eating Birds in a Mexican Successional Dry Forest. Animals, 11(12), 3343. https://doi.org/10.3390/ani11123343