
Infection of Mammary Glands of Small Mammals in Eastern North America by Helminths



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Examination of Hosts
2.2. Processing and Examination of Helminths and Mammary Tissue
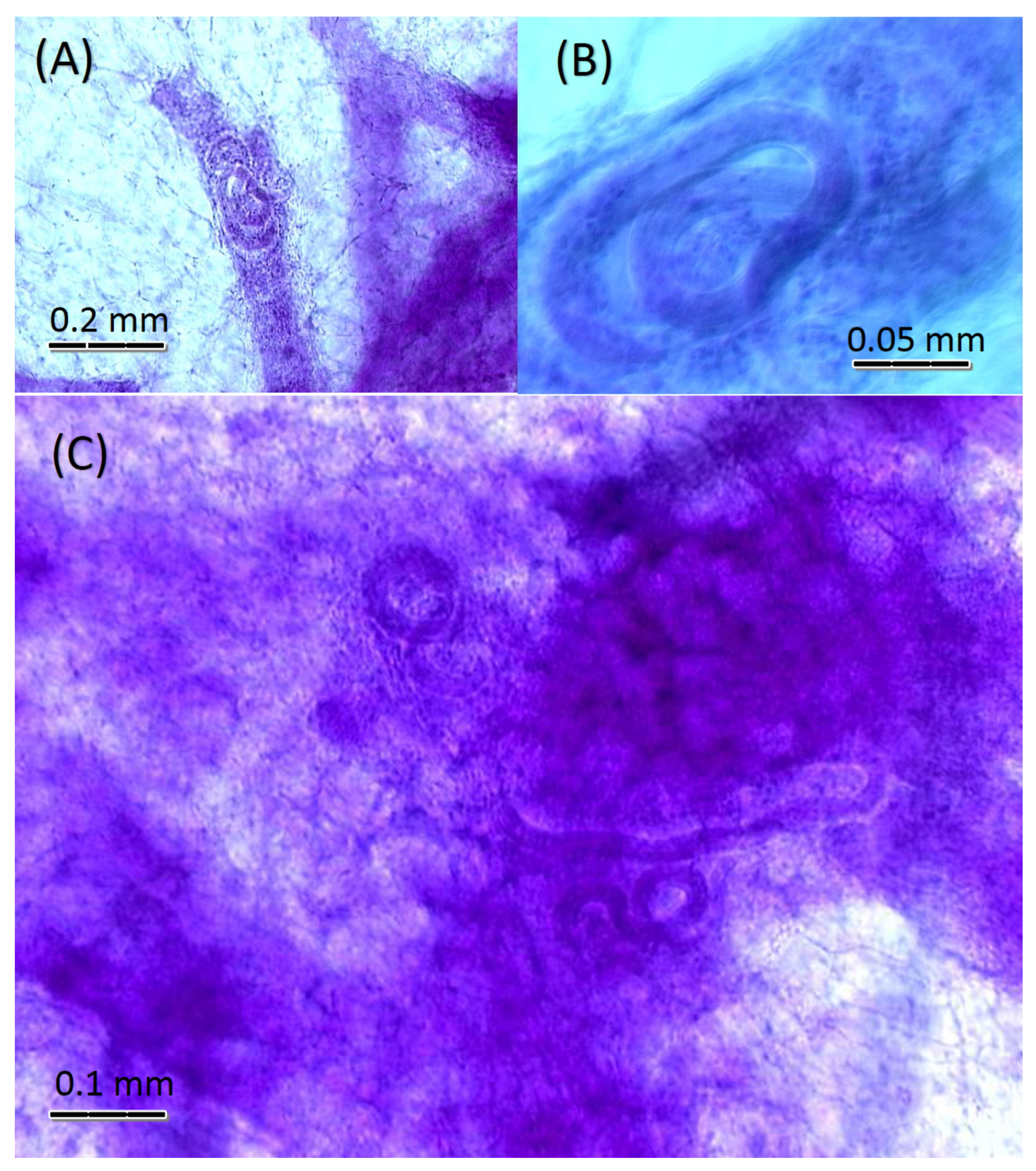
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baer, J.G. Transmission d’helminthes larvaires par le lait. Parassitologia 1972, 14, 11–15. [Google Scholar]
- Miller, G.C. Helminths and the transmammary route of infection. Parasitology 1981, 82, 335–342. [Google Scholar] [CrossRef]
- Shoop, W.L. Vertical transmission of helminths: Hypobiosis and amphiparatenesis. Parasitol. Today 1991, 7, 51–54. [Google Scholar] [CrossRef]
- Shoop, W.L. Vertical transmission in the Trematoda. J. Helminthol. Soc. Wash. 1994, 61, 153–161. [Google Scholar]
- Conn, D.B. Cestode infections of mammary glands and female reproductive organs: Potential for vertical transmission? J. Helminthol. Soc. Wash. 1994, 61, 162–168. [Google Scholar]
- Lyons, E.T. Vertical transmission of nematodes: Emphasis on Uncinaria lucasi in northern fur seals and Strongyloides westeri in equids. J. Helminthol. Soc. Wash. 1994, 61, 169–178. [Google Scholar]
- Stoye, V.M. Übersichtsreferat: Pränatale und galaktogene Helmintheninfektionen bei Haustieren. Dtsch. Tierärztl. Wochenschr. 1976, 83, 569–576. [Google Scholar] [PubMed]
- Conn, D.B.; Etges, F.J. Maternal transmission of asexually proliferative Mesocestoides corti tetrathyridia (Cestoda) in mice. J. Parasitol. 1983, 69, 922–925. [Google Scholar] [CrossRef]
- Williams, C.A.; Conn, D.B. Localization and associated histopathology of asexually proliferative Mesocestoides corti tetrathyridia (Cestoda) infecting mouse mammary glands. Int. J. Parasitol. 1985, 15, 245–248. [Google Scholar] [CrossRef]
- Matenga, E.; Mukaratirwa, S.; Bhebhe, E.; Willingham, A.L. Evidence of congenital and transmammary transmission of Trichinella zimbabwensis in rats (Rattus norvegicus) and its epidemiological implications. Intern. J. Appl. Res. Vet. Med. 2006, 4, 308–312. [Google Scholar]
- Sałamatin, R.V.; Pavlikovska, T.M.; Sagach, O.S.; Nikolayenko, S.M.; Kornyushin, V.V.; Kharchenko, V.O.; Masny, A.; Cielecka, D.; Konieczna-Sałamatin, J.; Conn, D.B.; et al. Human dirofilariasis due to Dirofilaria repens in Ukraine, an emergent zoonosis: Epidemiological report of 1465 cases. Acta Parasitol 2013, 58, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Nava, V.E.; Ejilemele, A.A. Images in clinical tropical medicine: Onchocerca volvulus mimicking metastatic breast carcinoma. Am. J. Trop. Med. Hyg. 2020, 102, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Koo, M.; Kim, J.H.; Kim, J.S.; Lee, J.E.; Nam, S.J.; Yang, J.H. Cases and literature review of breast sparganosis. World J. Surg. 2011, 35, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Ohbayashi, M.; Orihara, M.; Fujimake, Y. Mammaniduloides hokkaidensis n. g., n. sp. (Nematoda: Heligmosomatidae) from voles in Hokkaido. Jap. J. Vet. Res. 1968, 16, 23–29. [Google Scholar]
- Kinsella, J.M. Comparison of helminth parasites of the cotton rat, Sigmodon hispidus, from several habitats in Florida. Amer. Mus. Novitates 1974, 2540, 1–12. [Google Scholar]
- Boggs, J.F.; McMurry, S.T.; Leslie, D.M., Jr.; Engle, D.M.; Lochmiller, R.L. Influence of habitat modification on the community of gastrointestinal helminths of cotton rats. J. Wildl. Dis. 1991, 27, 584–593. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Melvin, D.M.; Chandler, A.D. New helminth records from the cotton rat, Sigmodon hispidus, including a new species, Strongyloides sigmodontis. J. Parasitol. 1950, 36, 505–510. Available online: https://www.jstor.org/stable/3273280 (accessed on 20 December 2020). [CrossRef]
- Harkema, R.; Kartman, L. Observations on the helminths and ectoparasites of the cotton rat, Sigmodon hispidus hispidus Say and Ord, in Georgia and North Carolina. J. Elisha Mitchell Sci. Soc. 1948, 64, 183–191. Available online: https://www.jstor.org/stable/24332885 (accessed on 20 December 2020).
- Coggins, J.R.; McDaniel, J.S. Helminth population dynamics in the cotton rat, Sigmodon hispidus. Proc. Okla. Acad. Sci. 1975, 55, 112–115. [Google Scholar]
- Martin, J.L.; Huffman, D.G. An analysis of the community and population dynamics of the helminths of Sigmodon hispidus (Rodentia: Cricetidae) from three central Texas vegetational regions. Proc. Helminthol. Soc. Wash. 1980, 47, 247–255. [Google Scholar]
- Kinsella, J.M.; Del Rosario Robles, M.; Preisser, W.C. A review of Gongylonema spp. (Nematoda: Gongylonematidae) in North American rodents with description of a new species from the cotton rat, Sigmodon hispidus (Mammalia: Cricetidae). Zootaxa 2016, 4107, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Harkema, R. The parasites of some North Carolina rodents. Ecol. Monogr. 1936, 6, 151–232. Available online: https://www.jstor.org/stable/1943242 (accessed on 20 December 2020). [CrossRef]
- Pung, O.J.; Durden, L.A.; Patrick, M.J.; Conyers, T.; Mitchell, L.R. Ectoparasites and gastrointestinal helminths of Southern Flying Squirrels in southeast Georgia. J. Parasitol. 2000, 86, 1051–1055. [Google Scholar] [CrossRef]
- Thatcher, V.E.; Scott, J.A. The life cycle of Trichostrongylus sigmodontis Baylis, 1945, and the susceptibility of various laboratory animals to this nematode. J. Parasitol. 1962, 48, 558–561. Available online: https://www.jstor.org/stable/3274910 (accessed on 20 December 2020). [CrossRef] [PubMed]
- Katz, F.F. Strongyloides ratti (Nematoda) in newborn offspring of inoculated rats. Proc. Pa. Acad. Sci. 1969, 43, 221–225. [Google Scholar]
- Nolan, T.J.; Katz, F.F. Transmammary transmission of Strongyloides venezuelensis (Nematoda) in rats. Proc. Helminthol. Soc. Wash. 1981, 48, 8–12. [Google Scholar]
- Wibawa, H.; Noviana, D.; Tsuchiya, K.; Satruja, F.; Horii, Y. Resistance of cotton rats, Sigmodon hispidus, to primary infection by Nippostrongylus brasiliensis. J. Vet. Med. Sci. 2002, 64, 423–426. [Google Scholar] [CrossRef][Green Version]
- Stone, W.; Smith, F.W. Infection of mammalian hosts by milk-borne nematode larvae: A review. Exp. Parasitol. 1973, 34, 306–312. [Google Scholar] [CrossRef]
- Conn, D.B.; Galán-Puchades, M.-T.; Fuentes, M.V. Interactions between anomalous excretory and tegumental epithelia in aberrant Mesocestoides tetrathyridia from Apodemus sylvaticus in Spain. Parasitol. Res. 2010, 96, 1109–1115. [Google Scholar] [CrossRef]
- Conn, D.B.; Galán-Puchades, M.-T.; Fuentes, M.V. Normal and aberrant Mesocestoides tetrathyridia from Crocidura spp. (Soricimorpha) in Corsica and Spain. J. Parasitol. 2011, 97, 915–919. [Google Scholar] [CrossRef]


{kind=link}
| Host Species | Collection Site | Number of Hosts Examined | Parasite Identity (Number of Hosts Infected; Prevalence as % Infected) |
|---|---|---|---|
| Blarina carolinensis Southern short-tailed shrew | Georgia | 1 | None present (0; 0%) |
| Clethrionomys gapperi Southern red-backed vole | New York | 4 | None present (0; 0%) |
| Mus musculus House mouse | Georgia | 11 | Pterygodermatites coloradensis (1; 9%) Syphacia obvelata (2; 18%) |
| Peromyscus leucopus White-footed mouse | New York | 5 | Hymenolepis nana (2; 40%) Schistosomatium douthitti (1; 20%) |
| Peromyscus leucopus | Tennessee | 25 | Pterygodermatites coloradensis (3; 12%) |
| Peromyscus leucopus | Georgia | 2 | Heligmosomoides sp. (1; 50%) |
| Sigmodon hispidus Hispid cotton rat | Georgia | 5 | Hymenolepis diminuta (1; 20%) Syphacia sigmodontis (1; 20%) Trichostrongylus sigmodontis (4; 80%) Mammary nematode larvae (4; 80%) |
| Total (5 host species) | 3 states | 53 | 8 species of adult; 1 species of larva (14; 26.4%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conn, D.B.; Hefty, C.A.; Owen, S.C. Infection of Mammary Glands of Small Mammals in Eastern North America by Helminths. Animals 2021, 11, 3207. https://doi.org/10.3390/ani11113207
Conn DB, Hefty CA, Owen SC. Infection of Mammary Glands of Small Mammals in Eastern North America by Helminths. Animals. 2021; 11(11):3207. https://doi.org/10.3390/ani11113207
Chicago/Turabian StyleConn, David Bruce, Cary A. Hefty, and Sarah Cross Owen. 2021. "Infection of Mammary Glands of Small Mammals in Eastern North America by Helminths" Animals 11, no. 11: 3207. https://doi.org/10.3390/ani11113207
APA StyleConn, D. B., Hefty, C. A., & Owen, S. C. (2021). Infection of Mammary Glands of Small Mammals in Eastern North America by Helminths. Animals, 11(11), 3207. https://doi.org/10.3390/ani11113207