
Does Social Complexity Drive Vocal Complexity? Insights from the Two African Elephant Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Forest Elephants
2.2. Savanna Elephants
3. Results
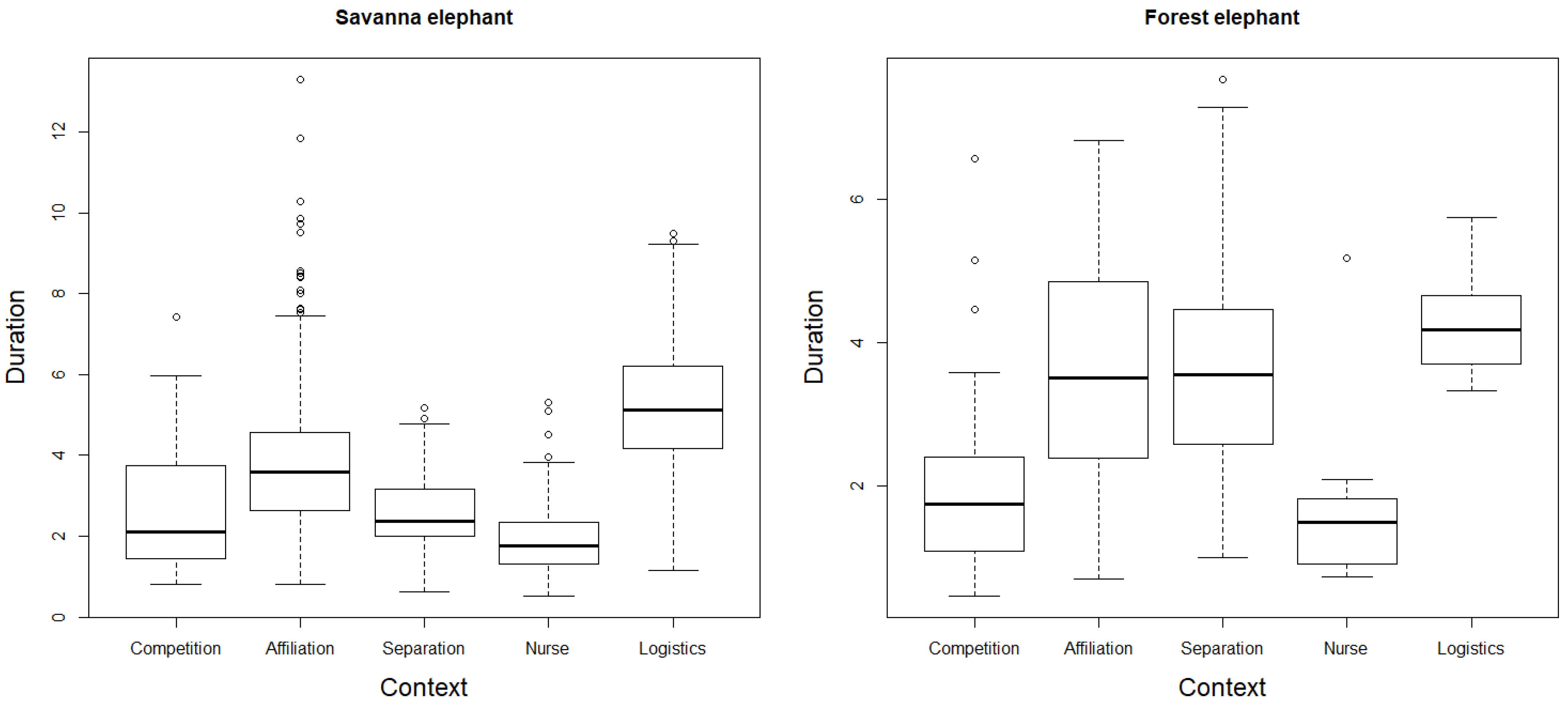
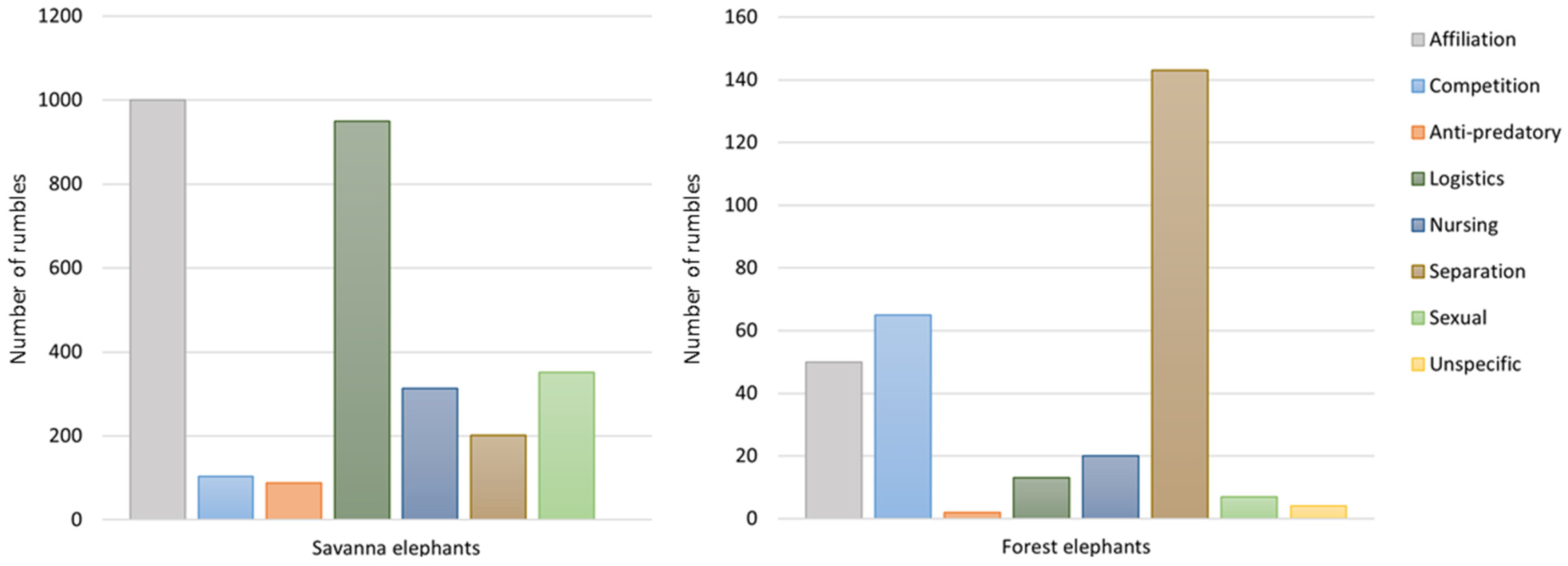
3.1. Rumble Repertoire Structure, Repertoire Size and Contextual Use
3.2. New Findings
3.3. Formant Modulation
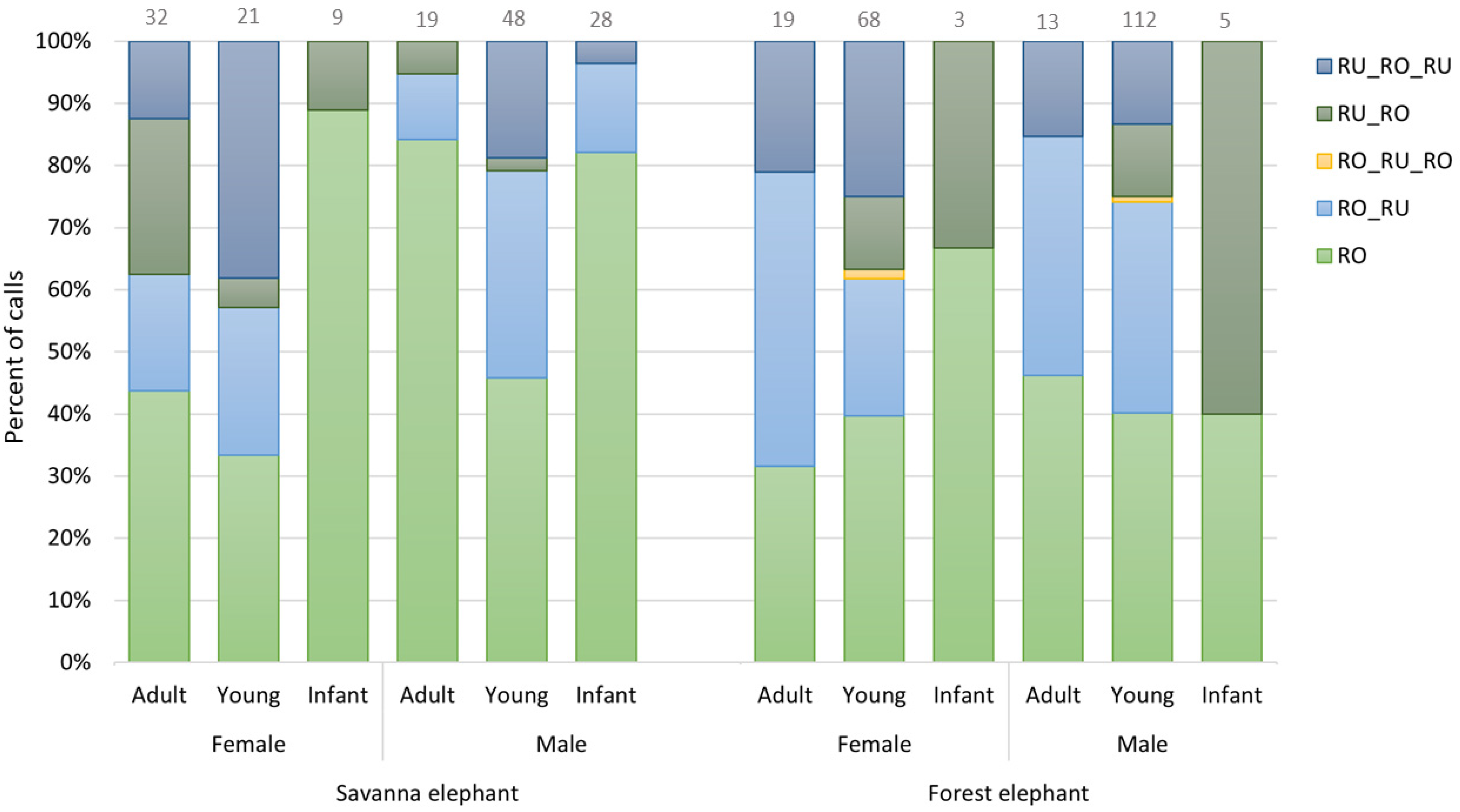
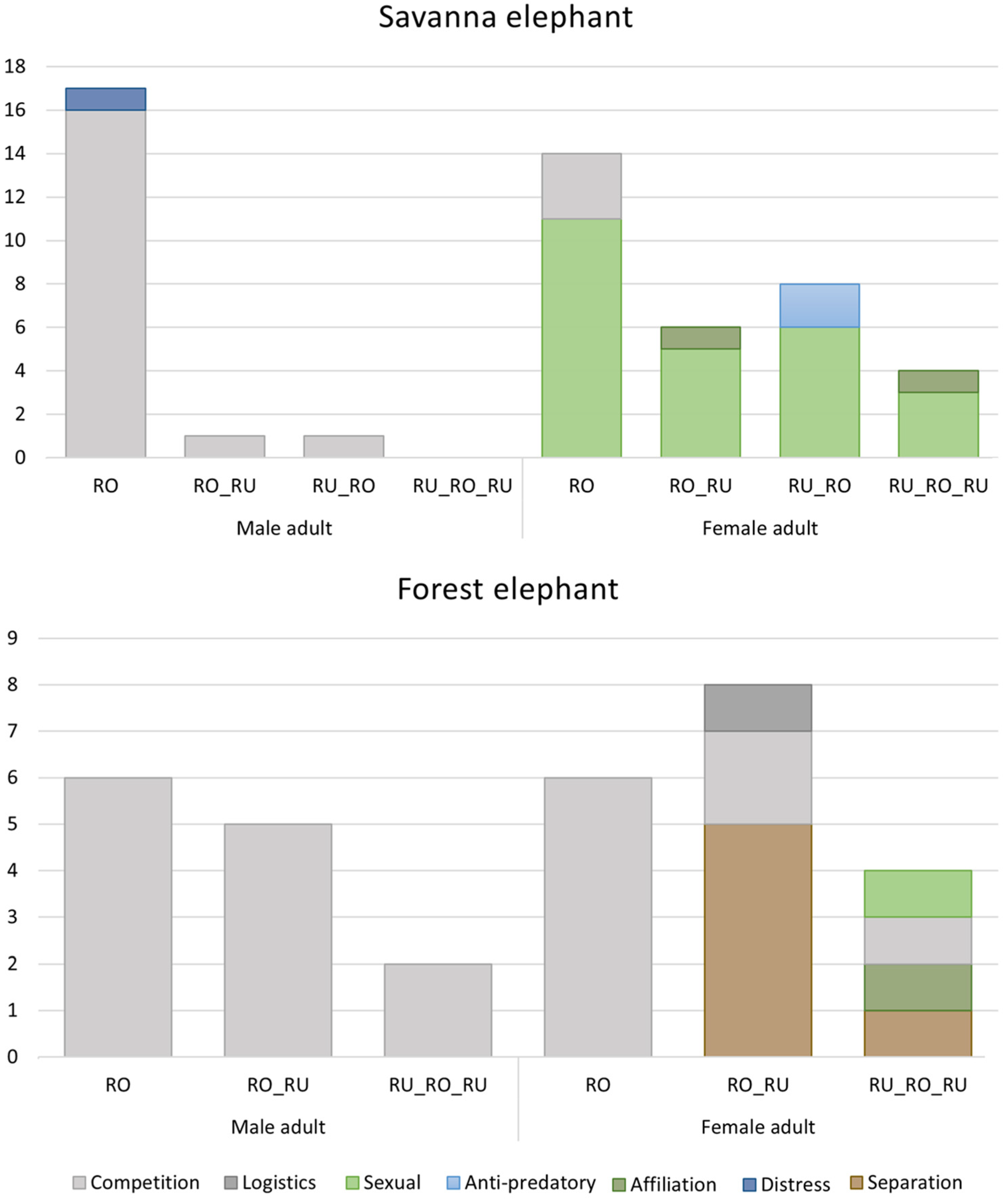
3.4. Call Combination
3.5. New Findings
3.6. Environmental Constraints on Vocal Mediation of Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krause, J.; Ruxton, G.D.; Ruxton, G.D. Living in Groups; OUP Oxford: Oxford, UK, 2002; ISBN 978-0-19-850818-2.
- Bradbury, J.W.; Vehrencamp, S.L. Principles of Animal Communication; Sinauer Associates Sunderland: Oxford, UK, 2011. [Google Scholar]
- Freeberg, T.M.; Dunbar, R.I.M.; Ord, T.J. Social complexity as a proximate and ultimate factor in communicative complexity. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1785–1801. [Google Scholar] [CrossRef] [PubMed]
- McComb, K.; Semple, S. Coevolution of vocal communication and sociality in primates. Biol. Lett. 2005, 1, 381–385. [Google Scholar] [CrossRef]
- Dunbar, R.I.M. Group size, vocal grooming and the origins of language. Psychon. Bull. Rev. 2017, 24, 209–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwin, C. The Expression of the Emotions in Man and Animals; Oxford University Press: Oxford, UK, 1872; ISBN 0-19-515806-7. [Google Scholar]
- Blumstein, D.T.; Armitage, K.B. Does sociality drive the evolution of communicative complexity? A comparative test with ground-dwelling sciurid alarm calls. Am. Nat. 1997, 150, 179–200. [Google Scholar] [CrossRef] [PubMed]
- Gustison, M.L.; le Roux, A.; Bergman, T.J. Derived Vocalizations of geladas (Theropithecus Gelada) and the evolution of vocal complexity in primates. Phil. Trans. R. Soc. B 2012, 367, 1847–1859. [Google Scholar] [CrossRef] [Green Version]
- Kroodsma, D.E. Correlates of song organization among North American Wrens. Am. Nat. 1977, 111, 995–1008. [Google Scholar] [CrossRef]
- Peckre, L.; Kappeler, P.M.; Fichtel, C. Clarifying and expanding the social complexity hypothesis for communicative complexity. Behav. Ecol. Sociobiol. 2019, 73, 11. [Google Scholar] [CrossRef]
- Lieberman, P.; Blumstein, S.E. Speech Physiology, Speech Perception, and Acoustic Phonetics; Cambridge University Press: Cambridge, MA, USA, 1988. [Google Scholar]
- Jackendoff, R. Possible stages in the evolution of the language capacity. Trends Cogn. Sci. 1999, 3, 272–279. [Google Scholar] [CrossRef]
- Aureli, F.; Schaffner, C.M.; Boesch, C.; Bearder, S.K.; Call, J.; Chapman, C.A.; Connor, R.; Fiore, A.D.; Dunbar, R.I.M.; Henzi, S.P.; et al. Fission-fusion dynamics: New research frameworks. Curr. Anthropol. 2008, 49, 627–654. [Google Scholar] [CrossRef]
- Couzin, I.D. Behavioral ecology: Social organization in fission–fusion societies. Curr. Biol. 2006, 16, R169–R171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlowe, F.W. Hunter-gatherers and human Evolution. Evol. Anthropol. Issues News Rev. 2005, 14, 54–67. [Google Scholar] [CrossRef]
- Bergman, T.J.; Beehner, J.C.; Painter, M.C.; Gustison, M.L. The speech-like properties of nonhuman primate vocalizations. Anim. Behav. 2019, 151. [Google Scholar] [CrossRef]
- Brandt, A.L.; Ishida, Y.; Georgiadis, N.J.; Roca, A.L. Forest elephant mitochondrial genomes reveal that elephantid diversification in Africa tracked climate transitions. Mol. Ecol. 2012, 21, 1175–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubb, P.; Groves, C.P.; Dudley, J.P.; Shoshani, J. Living African elephants belong to two species: Loxodonta Africana (Blumenbach, 1797) and Loxodonta Cyclotis (Matschie, 1900). Elephant 2000, 2, 3. [Google Scholar] [CrossRef]
- Murata, Y.; Yonezawa, T.; Kihara, I.; Kashiwamura, T.; Sugihara, Y.; Nikaido, M.; Okada, N.; Endo, H.; Hasegawa, M. Chronology of the extant African elephant species and case study of the species identification of the small African elephant with the molecular phylogenetic method. Gene 2009, 441, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Rohland, N.; Reich, D.; Mallick, S.; Meyer, M.; Green, R.E.; Georgiadis, N.J.; Roca, A.L.; Hofreiter, M. Genomic DNA sequences from mastodon and woolly mammoth reveal deep speciation of forest and savanna elephants. PLoS Biol. 2010, 8, e1000564. [Google Scholar] [CrossRef] [Green Version]
- Moss, C.; Poole, J. Relationships and social structure of African elephants. In Primate Social Relationships: An Integrated Approach; Blackwell Scientific Publications: Oxford, UK, 1983; pp. 315–325. [Google Scholar]
- Wittemyer, G.; Douglas-Hamilton, I.; Getz, W.M. The socioecology of elephants: Analysis of the processes creating multitiered social structures. Anim. Behav. 2005, 69, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Archie, E.A.; Moss, C.J.; Alberts, S.C. The ties that bind: Genetic relatedness predicts the fission and fusion of social groups in wild African Elephants. Proc. R. Soc. Lond. B Biol. Sci. 2006, 273, 513–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McComb, K.; Moss, C.; Sayialel, S.; Baker, L. Unusually extensive networks of vocal recognition in African elephants. Anim. Behav. 2000, 59, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Douglas-Hamilton, I. On the Ecology and Behaviour of the African Elephant: The Elephants of Lake Manyara. Ph.D. Thesis, Oxford University, Oxford, UK, 1972. [Google Scholar]
- Dublin, H.T. Cooperation and Reproductive competition among female African elephants. In Social Behavior of Female Vertebrates; Academic Press: Cambridge, MA, USA, 1983; pp. 291–313. [Google Scholar]
- Lee, P. Allomothering among African Elephants. Anim. Behav. 1987, 35, 278–291. [Google Scholar] [CrossRef]
- Foley, L.S. The Influence of Environmental Factors and Human Activity on Elephant Distribution. Ph.D. Thesis, International Institute for Geo-Information Science and Earth Observations, Wageningen, The Netherlands, 2002. [Google Scholar]
- Querouil, S.; Magliocca, F.; Gautier-Hion, A. Structure of population, grouping patterns and density of forest elephants in North-West Congo. Afr. J. Ecol. 2001, 37, 161–167. [Google Scholar] [CrossRef]
- Morgan, B.J.; Lee, P.C. Forest elephant group composition, frugivory and coastal use in the Réserve de Faune Du Petit Loango, Gabon. Afr. J. Ecol. 2007, 45, 519–526. [Google Scholar] [CrossRef]
- Turkalo, A.K.; Wrege, P.H.; Wittemyer, G. Long-term monitoring of Dzanga Bai Forest Elephants: Forest clearing use patterns. PLoS ONE 2013, 8, e85154. [Google Scholar] [CrossRef]
- Turkalo, A.; Fay, J. Studying forest elephants by direct observation. Pachyderm 1995, 20, 45–54. [Google Scholar]
- Turkalo, A.K.; Fay, J.M. Forest Elephant Behavior and Ecology; Yale University Press: London, UK, 2001. [Google Scholar]
- Fishlock, V.; Lee, P.C.; Breuer, T. Quantifying forest elephant social structure in Central African Bai environments. Pachyderm 2008, 44, 19–28. [Google Scholar]
- Fishlock, V.; Lee, P.C. Forest elephants: Fission–fusion and social arenas. Anim. Behav. 2013, 85, 357–363. [Google Scholar] [CrossRef]
- Goldenberg, S.Z.; Turkalo, A.K.; Wrege, P.H.; Hedwig, D.; Wittemyer, G. Entry and aggregation at a Central African Bai reveal social patterns in the Elusive Forest Elephant Loxodonta Cyclotis. Anim. Behav. 2021, 171, 77–85. [Google Scholar] [CrossRef]
- Poole, J.H.; Payne, K.; Langbauer, W.R.; Moss, C.J. The social contexts of some very low frequency calls of African Elephants. Behav. Ecol. Sociobiol. 1988, 22, 385–392. [Google Scholar] [CrossRef]
- Payne, K.B.; Thompson, M.; Kramer, L. Elephant calling patterns as indicators of group size and composition: The basis for an acoustic monitoring system. Afr. J. Ecol. 2003, 41, 99–107. [Google Scholar] [CrossRef]
- Hedwig, D.; Verahrami, A.K.; Wrege, P.H. Acoustic structure of Forest Elephant rumbles: A test of the ambiguity reduction hypothesis. Anim. Cognit. 2019, 22, 1115–1128. [Google Scholar] [CrossRef]
- Poole, J.H. Behavioral contexts of elephant acoustic communication. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; The University of Chicago: Chicago, IL, USA, 2011; pp. 125–161. [Google Scholar]
- Poole, J.; Granli, P. The Elephant Ethogram: A Library of African Elephant Behavior. Available online: https://www.elephantvoices.org/elephant-ethogram.html (accessed on 1 May 2021).
- Poole, J.H.; Granli, P. Signals, gestures, and behavior of African elephants. In The Amboseli Elephants; University of Chicago Press: Chicago, IL, USA, 2011; pp. 109–124. [Google Scholar]
- Pardo, M.A.; Poole, J.H.; Stoeger, A.S.; Wrege, P.H.; O’Connell-Rodwell, C.E.; Padmalal, U.K.; de Silva, S. Differences in combinatorial calls among the 3 elephant species cannot be explained by phylogeny. Behav. Ecol. 2019, 30, 809–820. [Google Scholar] [CrossRef]
- Pollard, K.A.; Blumstein, D.T. Evolving communicative complexity: Insights from rodents and beyond. Phil. Trans. R. Soc. B 2012, 367, 1869–1878. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, G.S. Social and vocal complexity in bats. In Animal Social Complexity: Intelligence, Culture, and Individualized Societies; Harvard University Press: Cambridge, MA, USA, 2003; pp. 322–341. ISBN 0-674-00929-0. [Google Scholar]
- Freeberg, T.M. Social complexity can drive vocal complexity: Group size influences vocal information in carolina chickadees. Psychol. Sci. 2006, 17, 557–561. [Google Scholar] [CrossRef]
- Manser, M.B.; Jansen, D.A.W.A.M.; Graw, B.; Hollén, L.I.; Bousquet, C.A.H.; Furrer, R.D.; le Roux, A. Chapter six-vocal complexity in meerkats and other mongoose species. In Advances in the Study of Behavior; Naguib, M., Barrett, L., Brockmann, H.J., Healy, S., Mitani, J.C., Roper, T.J., Simmons, L.W., Eds.; Academic Press: Cambidge, MA, USA, 2014; Volume 46, pp. 281–310. [Google Scholar]
- Stirling, I.; Thomas, J.A. Relationships between underwater vocalizations and mating systems in phocid seals. Aquat. Mamm. 2003, 29, 227–246. [Google Scholar] [CrossRef]
- Vanden Hole, C.; Van Daele, P.A.; Desmet, N.; Devos, P.; Adriaens, D. Does sociality imply a complex vocal communication system? A case study for Fukomys micklemi (Bathyergidae, Rodentia). Bioacoustics 2014, 23, 143–160. [Google Scholar] [CrossRef]
- Gustison, M.L.; Bergman, T.J. Vocal complexity influences female responses to gelada male calls. Sci. Rep. 2016, 6, 19680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ord, T.J.; Garcia-Porta, J. Is Sociality required for the evolution of communicative complexity? Evidence weighed against alternative hypotheses in diverse taxonomic groups. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1811–1828. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, E. Evolutionary origins of primate vocal communication: Diversity, flexibility, and complexity of vocalizations in basal primates. In Primate Hearing and Communication; Springer: Berlin/Heidelberg, Germany, 2017; pp. 109–140. [Google Scholar]
- Sugiura, H. Effects of proximity and behavioral context on acoustic variation in the coo calls of japanese macaques. Am. J. Primatol. 1993, 69, 1412–1424. [Google Scholar] [CrossRef]
- Elowson, A.M.; Snowdon, C.T. Pygmy marmosets, cebuella pygmaea, modify vocal structure in response to changed social environment. Anim. Behav. 1994, 47, 1267–1277. [Google Scholar] [CrossRef]
- Lemasson, A.; Hausberger, M. Acoustic variability and social significance of calls in female Campbell’s monkeys (Cercopithecus Campbelli Campbelli). J. Acoust. Soc. Am. 2011, 129, 3341–3352. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, H.; Blois-Heulin, C.; Lemasson, A. Social complexity parallels vocal complexity: A comparison of three non-human primate species. Front. Psychol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.D.; McCowan, B.; LANGBAUER JR, W.R.; Viljoen, J.J.; Hart, L.A. Classification of African elephant Loxodonta Africana rumbles using acoustic parameters and cluster analysis. Bioacoustics 2005, 15, 143–161. [Google Scholar] [CrossRef]
- Leong, K.M.; Ortolani, A.; Burks, K.D.; Mellen, J.D.; Savage, A. Quantifying acoustic and temporal characteristics of vocalizations for a group of captive African elephants Loxodonta Africana. Bioacoustics 2003, 13, 213–231. [Google Scholar] [CrossRef]
- Soltis, J.M.; Savage, A.; Leong, K.M. How many rumbles are there? Acoustic variation and individual identity in the rumble vocalizations of African elephants (Loxodonta Africana). J. Acoust. Soc. Am. 2004, 115, 2555. [Google Scholar] [CrossRef]
- Stoeger, A.S.; Heilmann, G.; Zeppelzauer, M.; Ganswindt, A.; Hensman, S.; Charlton, B.D. Visualizing sound emission of elephant vocalizations: Evidence for two rumble production types. PLoS ONE 2012, 7, e48907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, J.; King, L.E.; Douglas-Hamilton, I.; Vollrath, F.; Savage, A. African elephant alarm calls distinguish between threats from humans and bees. PLoS ONE 2014, 9, e89403. [Google Scholar] [CrossRef]
- Poole, J.H. Signals and assessment in African elephants: Evidence from playback experiments. Anim. Behav. 1999, 58, 185–193. [Google Scholar] [CrossRef]
- Fant, G. The source filter concept in voice production. STL-QPSR 1981, 1, 21–37. [Google Scholar]
- Taylor, A.M.; Charlton, B.D.; Reby, D. Vocal production by terrestrial mammals: Source, filter, and function. In Vertebrate Sound Production and Acoustic Communication; Springer Handbook of Auditory Research; Springer: Cham, Switzerland, 2016; pp. 229–259. ISBN 978-3-319-27719-6. [Google Scholar]
- Fitch, W.T.; Giedd, J. Morphology and development of the human vocal tract: A study using magnetic resonance imaging. J. Acoust. Soc. Am. 1999, 106, 1511–1522. [Google Scholar] [CrossRef] [Green Version]
- Rendall, D. Acoustic correlates of caller identity and affect intensity in the vowel-like grunt vocalizations of baboons. J. Acoust. Soc. Am. 2003, 113, 3390. [Google Scholar] [CrossRef]
- Reby, D.; McComb, K. Anatomical constraints generate honesty: Acoustic cues to age and weight in the roars of red deer stags. Anim. Behav. 2003, 65, 519–530. [Google Scholar] [CrossRef] [Green Version]
- González, J. Formant frequencies and body size of speaker: A weak relationship in adult humans. J. Phon. 2004, 32, 277–287. [Google Scholar] [CrossRef]
- Pisanski, K.; Fraccaro, P.J.; Tigue, C.C.; O’Connor, J.J.M.; Röder, S.; Andrews, P.W.; Fink, B.; DeBruine, L.M.; Jones, B.C.; Feinberg, D.R. Vocal indicators of body size in men and women: A meta-analysis. Anim. Behav. 2014, 95, 89–99. [Google Scholar] [CrossRef]
- Morton, E.S. On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. Am. Nat. 1977, 111, 855–869. [Google Scholar] [CrossRef]
- Riede, T.; Bronson, E.; Hatzikirou, H.; Zuberbühler, K. vocal production mechanisms in a non-human primate: Morphological data and a model. J. Hum. Evol. 2005, 48, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberman, P.H.; Klatt, D.H.; Wilson, W.H. Vocal Tract limitations on the vowel repertoires of Rhesus Monkey and other nonhuman primates. Science 1969, 164, 1185–1187. [Google Scholar] [CrossRef]
- Fitch, W.T.; de Boer, B.; Mathur, N.; Ghazanfar, A.A. Monkey vocal tracts are speech-ready. Sci. Adv. 2016, 2, e1600723. [Google Scholar] [CrossRef] [Green Version]
- Fitch, W.T.; de Boer, B.; Mathur, N.; Ghazanfar, A.A. Response to lieberman on “Monkey vocal tracts are speech-ready”. Sci. Adv. 2017, 3, e1701859. [Google Scholar] [CrossRef] [Green Version]
- Boë, L.-J.; Berthommier, F.; Legou, T.; Captier, G.; Kemp, C.; Sawallis, T.R.; Becker, Y.; Rey, A.; Fagot, J. Evidence of a vocalic proto-system in the baboon (Papio Papio) suggests pre-hominin speech precursors. PLoS ONE 2017, 12, e0169321. [Google Scholar] [CrossRef]
- Andrew, R.J. Use of formants in the grunts of baboons and other nonhuman primates*. Ann. N. Y. Acad. Sci. 1976, 280, 673–693. [Google Scholar] [CrossRef]
- Owren, M.J.; Bernacki, R.H. The acoustic features of vervet monkey alarm calls. J. Acoust. Soc. Am. 1988, 83, 1927–1935. [Google Scholar] [CrossRef]
- Riede, T.; Zuberbühler, K. Pulse register phonation in Diana Monkey alarm calls. J. Acoust. Soc. Am. 2003, 113, 2919–2926. [Google Scholar] [CrossRef] [Green Version]
- Riede, T.; Zuberbühler, K. The relationship between acoustic structure and semantic information in Diana monkey alarm vocalization. J. Acoust. Soc. Am. 2003, 114, 1132–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoshani, J.; Marchant, G. Hyoid Apparatus: A Little Known Complex of Bones and Its Contribution to Proboscidean Evolution; Consiglio Nazionale delle Ricerche Roma: Rome, Italy, 2001; pp. 668–675. [Google Scholar]
- Herbst, C.T.; Stoeger, A.S.; Frey, R.; Lohscheller, J.; Titze, I.R.; Gumpenberger, M.; Fitch, W.T. How low can you go? Physical production mechanism of elephant infrasonic vocalizations. Science 2012, 337, 595–599. [Google Scholar] [CrossRef]
- Poole, J.H.; Tyack, P.L.; Stoeger-Horwath, A.S.; Watwood, S. Animal behaviour: Elephants are capable of vocal learning. Nature 2005, 434, 455–456. [Google Scholar] [CrossRef]
- Stoeger, A.S.; Baotic, A. Operant control and call usage learning in African elephants. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200254. [Google Scholar] [CrossRef] [PubMed]
- McComb, K.; Reby, D.; Baker, L.; Moss, C.; Sayialel, S. Long-distance communication of acoustic cues to social identity in African elephants. Anim. Behav. 2003, 65, 317–329. [Google Scholar] [CrossRef] [Green Version]
- King, L.E.; Soltis, J.; Douglas-Hamilton, I.; Savage, A.; Vollrath, F. Bee threat elicits alarm call in African elephants. PLoS ONE 2010, 5, e10346. [Google Scholar] [CrossRef] [Green Version]
- Soltis, J.; Leighty, K.A.; Wesolek, C.M.; Savage, A. The expression of affect in African elephant (Loxodonta Africana) Rumble vocalizations. J. Comp. Psychol. 2009, 123, 222–225. [Google Scholar] [CrossRef]
- Marler, P. The structure of animal communication sounds. In Recognition of Complex Acoustic Signals, Dahlem Konferenzen; Abakon-Verlagsgesellschaft: Berlin, Germany, 1977; pp. 17–35. [Google Scholar]
- Hurford, J.R. The Origins of Grammar: Language in the Light of Evolution II; OUP Oxford: Oxford, UK, 2011. [Google Scholar]
- Fitch, W.T. The Evolution of Language; Cambridge University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Collier, K.; Bickel, B.; van Schaik, C.P.; Manser, M.B.; Townsend, S.W. Language evolution: Syntax before phonology? Proc. R. Soc. B Biol. Sci. 2014, 281, 20140263. [Google Scholar] [CrossRef] [Green Version]
- Schassburger, R. Vocal Communication in the Timber Wolf; Paul Parey Scientific Publishers: Berlin, Germany, 1993. [Google Scholar]
- Collier, K.; Townsend, S.W.; Manser, M.B. Call Concatenation in wild meerkats. Anim. Behav. 2017, 134, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Zuberbühler, K. Combinatorial capacities in primates. Curr. Opin. Behav. Sci. 2018, 21, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Brenowitz, E.A. The active space of red-winged blackbird song. J. Comp. Physiol. 1982, 147, 511–522. [Google Scholar] [CrossRef]
- Miller, P.J.O. Diversity in sound pressure levels and estimated active space of resident killer whale vocalizations. J. Comp. Physiol. A 2006, 192, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, N.P.M. Estimated source intensity and active space of the American alligator (Alligator Mississippiensis) vocal display. J. Acoust. Soc. Am. 2007, 122, 2906–2915. [Google Scholar] [CrossRef] [PubMed]
- Charlton, B.D.; Reby, D.; Ellis, W.A.; Brumm, J.; Fitch, W.T. Estimating the active space of male koala bellows: Propagation of cues to size and identity in a eucalyptus forest. PLoS ONE 2012, 7, e45420. [Google Scholar] [CrossRef] [PubMed]
- Hedwig, D.; DeBellis, M.; Wrege, P.H. Not so far: Attenuation of low-frequency vocalizations in a rainforest environment suggests limited acoustic mediation of social interaction in African forest elephants. Behav. Ecol. Sociobiol. 2018, 72, 33. [Google Scholar] [CrossRef]
- Marten, K.; Marler, P. Sound transmission and its significance for animal vocalization. Behav. Ecol. Sociobiol. 1977, 2, 271–290. [Google Scholar] [CrossRef]
- Larom, D.; Garstang, M.; Payne, K.; Raspet, R.; Lindeque, M. The influence of surface atmospheric conditions on the range and area reached by animal vocalizations. J. Exp. Biol. 1997, 200, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Stoeger, A.S.; Charlton, B.D.; Kratochvil, H.; Fitch, W.T. Vocal cues indicate level of arousal in infant African elephant roars. J. Acoust. Soc. Am. 2011, 130, 1700–1710. [Google Scholar] [CrossRef]
- Seyfarth, R.M.; Cheney, D.L.; Marler, P. Vervet Monkey Alarm Calls: Semantic communication in a free-ranging primate. Anim. Behav. 1980, 28, 1070–1094. [Google Scholar] [CrossRef]
- Ouattara, K.; Lemasson, A.; Zuberbühler, K. Campbell’s monkeys use affixation to alter call meaning. PLoS ONE 2009, 4, e7808. [Google Scholar] [CrossRef] [Green Version]
- Ord, T.J.; Martins, E.P. Tracing the origins of signal diversity in anole lizards: Phylogenetic approaches to inferring the evolution of complex behaviour. Anim. Behav. 2006, 71, 1411–1429. [Google Scholar] [CrossRef]
- Wischmann, S.; Floreano, D.; Keller, L. Historical contingency affects signaling strategies and competitive abilities in evolving populations of simulated robots. Proc. Natl Acad. Sci. USA 2012, 109, 864–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkalo, A.K.; Wrege, P.H.; Wittemyer, G. Slow Intrinsic growth rate in forest elephants indicates recovery from poaching will require decades. J. Appl. Ecol. 2017, 54, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Sibly, R.M.; Brown, J.H. Effects of body size and lifestyle on evolution of mammal life histories. Proc. Natl Acad. Sci. USA 2007, 104, 17707–17712. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.H. Primates and the evolution of long, slow life histories. Curr. Biol. 2011, 21, R708–R717. [Google Scholar] [CrossRef] [Green Version]
- Emery, N.J.; Seed, A.M.; von Bayern, A.M.; Clayton, N.S. Cognitive adaptations of social bonding in birds. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 489–505. [Google Scholar] [CrossRef] [Green Version]
- Moss, C.J. The demography of an African elephant (Loxodonta Africana) population in Amboseli, Kenya. J. Zool. 2001, 255, 145–156. [Google Scholar] [CrossRef]
- Gobush, K.S.; Edwards, C.T.T.; Maisels, F.; Wittemyer, G.; Balfour, D.; Taylor, R.D. Loxodonta Cyclotis; International Union for Conservation of Nature: Gland, Switzerland, 2021. [Google Scholar]
- Langbauer, W.R.; Payne, K.B.; Charif, R.A.; Rapaport, L.; Osborn, F. African elephants respond to distant playbacks of low-frequency conspecific calls. J. Exp. Biol. 1991, 157, 35–46. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Elephants | Savanna Elephants | ||
|---|---|---|---|
| Context | Description | Context | Definition |
| Affiliation | Individuals in the Bai, or upon entering the Bai approaching one another, or walking up to a stationary individual, usually leading to benign physical contact. Individuals may stay in proximity, feeding, traveling, resting, or sharing a mineral pit. | Affiliative | Vocalizations given during the formation and maintenance of social bonds with other elephants (greeting-rumble, little-greeting-rumble, rumbles associated with greeting-ceremonies, some bonding-ceremonies and reconciliations, rumble-roar, rumble-roar-rumble). |
| Birth | Vocalizations given by a newborn, its family members, or nearby elephants during, or within 24 h prior to, or after, parturition, that relates to the birth (coo-rumble, bonding-rumble). | ||
| Calf Reassurance and Protection | Vocalizations given when defending, protecting, helping, guiding, comforting or consoling calves, or by calves responding to such actions (coo-rumble, as-touched-rumble). | ||
| Competition | Conflict over access to resources, mostly mineral pits, occasionally grass. Includes displacements, avoidance interactions, nonphysical and physical aggression, as well as disputes by individuals sharing a mineral pit. | Aggressive | Vocalizations given when displacing, intimidating, threatening or attacking other elephants (rumble). |
| Submissive | Vocalizations given in submission to avoid aggression and/or injury by other elephants (V8-rumble). | ||
| Protest | Vocalizations given when in pain, or when complaining or protesting at some perceived wrong—in a competitive situation (grumble-rumble, roar, roar-rumble, rumble-roar). | ||
| Separation | Individuals travelling alone at a large distance from associates or when associates have left the clearing. Individuals often wander around aimlessly, smelling ground with their trunk, frequently stopping to listen, or running in distress with ears and tail erect. | Separation | Vocalizations given by a calf or juvenile elephant when separated from its mother or family. May call repeatedly while wandering around sniffing the ground or air with the trunk, stopping to listen, or running in distress with ears spread and tail raised (separated-rumble, roar-rumble, rumble-roar-rumble). |
| Anti-predatory | Individual mobbing (charging, kicking dust or flinging trunk) animal of other species, or when in a state of alert (ears erect and tail up) upon visitors arriving at the observation platform. | Vigilance | Vocalizations given when attending to, or alerting companions to, possible danger posed by potential predators (comment-rumble). |
| Conflict and Confrontation | Vocalizations given when signaling hostility toward a perceived non-elephant threat (rumble). | ||
| Coalition Building | Vocalizations given during the formation of a defensive coalition (rumble). | ||
| Mobbing and Attacking | Vocalizations given by an elephant, or a group of elephants acting cooperatively, when advancing upon, harassing, or attacking a non-elephant threat (rumble, rumble-roar). | ||
| Avoidance | Vocalizations given when evading actual or perceived dangers posed by non-elephant threats (rumble). | ||
| Sexual | Female presenting her hindquarters allowing males to inspect genitals, often urinating while doing so. Males contesting over females. Females, often in distress, avoiding a male who is pursuing them. | Advertisement and Attraction | Vocalizations given by adult male and adult female elephants in the advertisement of sexual state and the attraction of, and search for, mates (musth-rumble, rumbles given during a female-chorus, estrous-running-rumble, estrous-roar, estrous-running-rumble-roar, roar, roar-rumble, rumble-roar, rumble-roar-rumble). |
| Courtship | Vocalizations given by adult male and female elephants during consorting and mating (estrous-running-rumble, estrous-roar, estrous-running-rumble-roar, estrous-rumble, calls within a mating-pandemonium or female-chorus including, rumbles, roar-rumble, rumble-roar, rumble-roar-rumble). | ||
| Nursing | Calf approaching or walking parallel along mother’s side with trunk raised, often touching mother’s side, leg or breast. The mother usually stops and starts nursing. | Calf Nourishment and Weaning | Vocalizations given by calves when attempting to meet their nutritional requirements and during the process of weaning (begging-rumble, cry, cry-rumble, roar, roar-rumble, rumble-roar, rumble-roar-rumble). |
| Logistics | Concerted group movement and group coordination as individuals travel or leave the clearing together. | Movement, Space and Leadership | Vocalizations given when maintaining spatial proximity, initiating group movement or influencing its timing, direction or form (let’s-go-rumble, cadenced-rumble, contact-rumble). |
| Distress | Vocalizations given when in pain, or when complaining or protesting at some perceived wrong in a non-competitive situation (roar, roar-rumble, rumble-roar-rumble). | ||
| Unspecific | Individuals are stationary feeding on grass, accessing mineral pits or resting, often within 20 m of associates, no obvious social interaction or change in activity, often exchanging rumbles. | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedwig, D.; Poole, J.; Granli, P. Does Social Complexity Drive Vocal Complexity? Insights from the Two African Elephant Species. Animals 2021, 11, 3071. https://doi.org/10.3390/ani11113071
Hedwig D, Poole J, Granli P. Does Social Complexity Drive Vocal Complexity? Insights from the Two African Elephant Species. Animals. 2021; 11(11):3071. https://doi.org/10.3390/ani11113071
Chicago/Turabian StyleHedwig, Daniela, Joyce Poole, and Petter Granli. 2021. "Does Social Complexity Drive Vocal Complexity? Insights from the Two African Elephant Species" Animals 11, no. 11: 3071. https://doi.org/10.3390/ani11113071
APA StyleHedwig, D., Poole, J., & Granli, P. (2021). Does Social Complexity Drive Vocal Complexity? Insights from the Two African Elephant Species. Animals, 11(11), 3071. https://doi.org/10.3390/ani11113071