
Effects of Ionophores on Ruminal Function of Beef Cattle


Abstract
Simple Summary
Abstract
1. Introduction
2. Ionophores Mode of Action
3. Ionophores and Animal Production
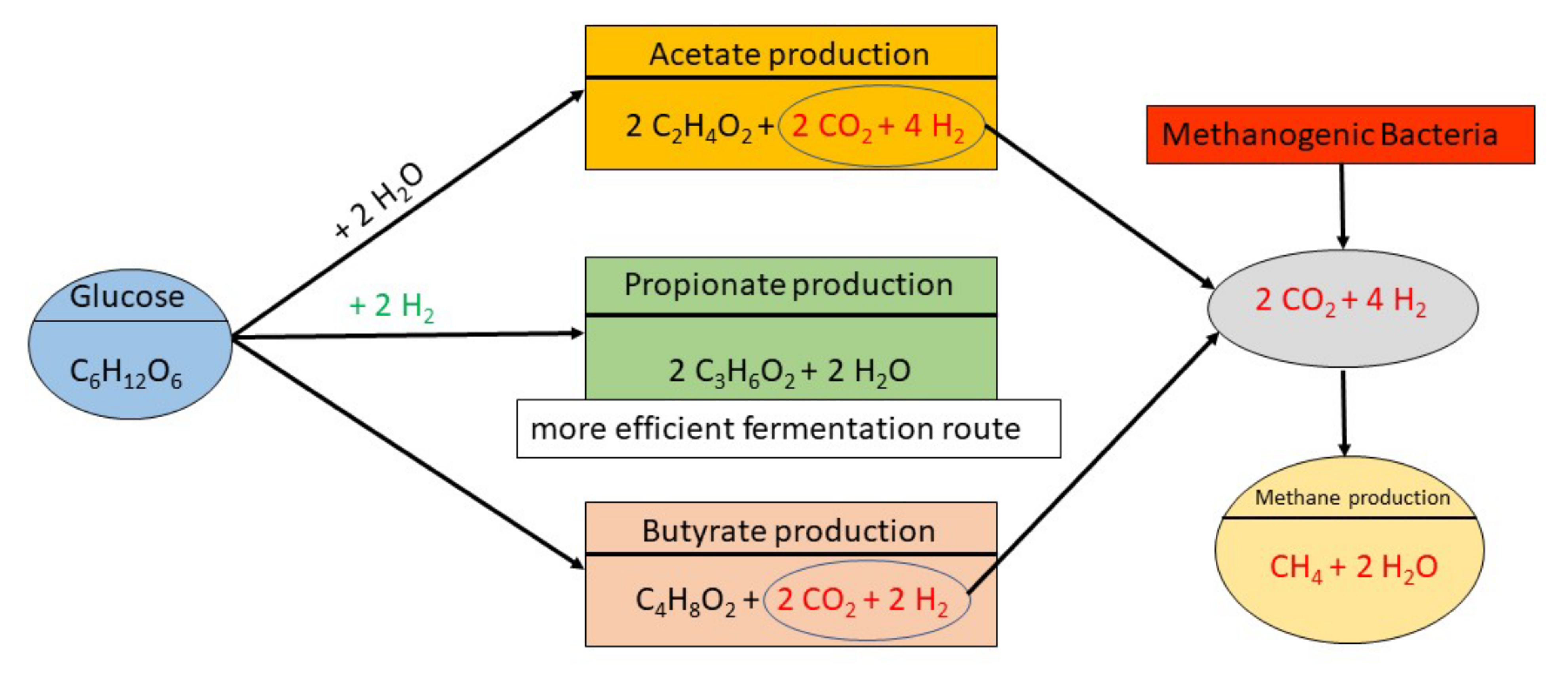
4. Ionophores and Rumen Fermentation Function
5. Ionophores and Ruminal Nitrogen Metabolism
6. Ionophores’ Persistence
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bretschneider, G.; Elizalde, J.C.; Pérez, F.A. The effect of feeding antibiotic growth promoters on the performance of beef cattle consuming forage-based diets: A Review. Livest. Sci. 2008, 114, 135–149. [Google Scholar] [CrossRef]
- Duffield, T.F.; Merrill, J.K.; Bagg, R.N. Meta-analysis of the effects of monensin in beef cattle on feed efficiency, body weight gain, and dry matter intake. J. Anim. Sci. 2012, 90, 4583–4592. [Google Scholar] [CrossRef]
- Weimer, P.J.; Stevenson, D.M.; Mertens, D.R.; Thomas, E.E. Effect of monensin feeding and withdrawal on populations of individual bacterial species in the rumen of lactating dairy cows fed high-starch rations. Appl. Microbiol. Biotechnol. 2008, 80, 135–145. [Google Scholar] [CrossRef]
- Schären, M.; Drong, C.; Kiri, K.; Riede, S.; Gardener, M.; Meyer, U.; Hummel, J.; Urich, T.; Breves, G.; Dänicke, S. Differential effects of monensin and a blend of essential oils on rumen microbiota composition of transition dairy cows. J. Dairy Sci. 2017, 100, 2765–2783. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O.; Fox, D.G.; Tylutki, T.P. Potential environmental benefits of ionophores in ruminant diets. J. Environ. Qual. 2003, 32, 1591–1602. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Lechtenberg, K.F. Acidosis in feedlot cattle. Vet. Clin. N. Am. Food Anim. Pract. 2007, 23, 333–350. [Google Scholar] [CrossRef]
- Russell, J.B.; Strobel, H.J. Effect of ionophores on ruminal fermentation. Appl. Environ. Microbiol. 1989, 55, 1–6. [Google Scholar] [CrossRef] [PubMed]
- McGuffey, R.K.; Richardson, L.F.; Wilkinson, J.I.D. Ionophores for dairy cattle: Current status and future outlook. J. Dairy Sci. 2001, 84, E194–E203. [Google Scholar] [CrossRef]
- Nagaraja, T.G. Ionophores and antibiotics in ruminants. In Biotechnology in Animal Feeds and Animal Feeding; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1995; pp. 173–204. ISBN 978-3-527-61535-3. [Google Scholar]
- Ensley, S. Ionophore use and toxicosis in cattle. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 641–652. [Google Scholar] [CrossRef]
- Dinius, D.A.; Simpson, M.E.; Marsh, P.B. Effect of monensin fed with forage on digestion and the ruminal ecosystem of steers. J. Anim. Sci. 1976, 42, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.F.; Raun, A.P.; Potter, E.L.; Cooley, C.O.; Rathmacher, R.P. Effect of monensin on rumen fermentation in vitro and in vivo. J. Anim. Sci. 1976, 43, 657–664. [Google Scholar] [CrossRef]
- Dennis, S.M.; Nagaraja, T.G.; Bartley, E.E. Effect of lasalocid or monensin on lactate production from in vitro rumen fermentation of various carbohydrates. J. Dairy Sci. 1981, 64, 2350–2356. [Google Scholar] [CrossRef]
- Golder, H.M.; Lean, I.J. A meta-analysis of lasalocid effects on rumen measures, beef and dairy performance, and carcass traits in cattle. J. Anim. Sci. 2016, 94, 306–326. [Google Scholar] [CrossRef]
- Ellis, J.L.; Dijkstra, J.; Bannink, A.; Kebreab, E.; Hook, S.E.; Archibeque, S.; France, J. Quantifying the effect of monensin dose on the rumen volatile fatty acid profile in high-grain-fed beef cattle. J. Anim. Sci. 2012, 90, 2717–2726. [Google Scholar] [CrossRef]
- Goodrich, R.D.; Garrett, J.E.; Gast, D.R.; Kirick, M.A.; Larson, D.A.; Meiske, J.C. Influence of monensin on the performance of cattle. J. Anim. Sci. 1984, 58, 1484–1498. [Google Scholar] [CrossRef]
- Rogers, M.; Jouany, J.P.; Thivend, P.; Fontenot, J.P. The effects of short-term and long-term monensin supplementation, and its subsequent withdrawal on digestion in sheep. Anim. Feed Sci. Tech. 1997, 65, 113–127. [Google Scholar] [CrossRef]
- Pressman, B.C. Biological applications of ionophores. Annu. Rev. Biochem. 1976, 45, 501–530. [Google Scholar] [CrossRef]
- Russell, J.B. A Proposed mechanism of monensin action in inhibiting ruminant bacterial growth: Effects on ion flux and protonmotive force. J. Anim. Sci. 1987, 64, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, Y.A. Physico-chemical basis of ion transport through biological membranes: Ionophores and ion channels. Eur. J. Biochem. 1979, 94, 321–336. [Google Scholar] [CrossRef]
- Booth, I.R. Regulation of cytoplasmic ph in bacteria. Microbiol. Rev. 1985, 49, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Painter, G.R.; Pollack, R.; Pressman, B.C. Conformational dynamics of the carboxylic ionophore lasalocid a underlying cation complexation-decomplexation and membrane transport. Biochemistry 1982, 21, 5613–5620. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Houlihan, A.J. Ionophore resistance of ruminal bacteria and its potential impact on human health. FEMS Microbiol. Rev. 2003, 27, 65–74. [Google Scholar] [CrossRef]
- Kim, M.; Felix, T.L.; Loerch, S.C.; Yu, Z. Effect of haylage and monensin supplementation on ruminal bacterial communities of feedlot cattle. Curr. Microbiol. 2014, 69, 169–175. [Google Scholar] [CrossRef]
- Kim, M.; Eastridge, M.L.; Yu, Z. Investigation of ruminal bacterial diversity in dairy cattle fed supplementary monensin alone and in combination with fat, using pyrosequencing analysis. Can. J. Microbiol. 2014, 60, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.J.; Costerton, J.W. Ultrastructure of butyrivibrio fibrisolvens: A gram-positive bacterium. J. Bacteriol. 1977, 129, 1506–1512. [Google Scholar] [CrossRef]
- Chen, M.; Wolin, M.J. Effect of monensin and lasalocid-sodium on the growth of methanogenic and rumen saccharolytic bacteria. Appl. Environ. Microbiol. 1979, 38, 72–77. [Google Scholar] [CrossRef]
- Azzaz, H.H.; Murad, H.A.; Morsy, T.A. Utility of ionophores for ruminant animals: A review. Asian J. Anim. Sci. 2015, 9, 254–265. [Google Scholar] [CrossRef]
- Duffield, T.F.; Sandals, D.; Leslie, K.E.; Lissemore, K.; McBride, B.W.; Lumsden, J.H.; Dick, P.; Bagg, R. Efficacy of monensin for the prevention of subclinical ketosis in lactating dairy cows. J. Dairy Sci. 1998, 81, 2866–2873. [Google Scholar] [CrossRef]
- Duffield, T.F.; Rabiee, A.R.; Lean, I.J. A meta-analysis of the impact of monensin in lactating dairy cattle. part 1. metabolic effects. J. Dairy Sci. 2008, 91, 1334–1346. [Google Scholar] [CrossRef]
- Limede, A.C.; Marques, R.S.; Polizel, D.M.; Cappellozza, B.I.; Miszura, A.A.; Barroso, J.P.R.; Storti Martins, A.; Sardinha, L.A.; Baggio, M.; Pires, A.V. Effects of supplementation with narasin, salinomycin, or flavomycin on performance and ruminal fermentation characteristics of bos indicus nellore cattle fed with forage-based diets. J. Anim. Sci. 2021, 99. [Google Scholar] [CrossRef]
- Beck, P.; Galyen, W.; Galloway, D.; Kegley, E.B.; Rorie, R.; Hubbell, D.; Tucker, J.; Hess, T.; Cravey, M.; Hill, J.; et al. Effect of supplementation of developing replacement heifers with monensin or bambermycins on gain and pregnancy rates. Prof. Anim. Sci. 2016, 32, 619–626. [Google Scholar] [CrossRef]
- Polizel, D.M.; Cappellozza, B.I.; Hoe, F.; Lopes, C.N.; Barroso, J.P.; Miszura, A.; Oliveira, G.B.; Gobato, L.; Pires, A.V. Effects of narasin supplementation on dry matter intake and rumen fermentation characteristics of bos indicus steers fed a high-forage diet. Transl. Anim. Sci. 2020, 4, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Soares, L.C.B.; Marques, R.S.; Pires, A.V.; Cruz, V.A.; Limede, A.C.; Maia, K.S.; Baggio, M.; Barroso, J.P.R.; Biava, J.S.; Ferreira, E.M.; et al. Effects of narasin supplementation frequency on intake, ruminal fermentation parameters, and nutrient digestibility of bos indicus nellore steers fed with forage-based diets. Transl. Anim. Sci. 2021. [Google Scholar] [CrossRef]
- DelCurto, T.; Earley, A.; May, T.; Nichols, W. Comparison of bambermycins, lasalocid, and monensin on the nutritional physiology of beef cattle consuming concentrate and forage base diets. In Special Report 991; EOARC Annual Report; Agricultural Experiment Station, Oregon State University: Corvallis, OR, USA, 1999. [Google Scholar]
- Sutton, J.D. Carbohydrate fermentation in the rumen—Variations on a theme. P. Nutr. Soc. 1979, 38, 275–281. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Wolin, M.J.; Miller, T.L.; Stewart, C.S. Microbe-microbe interactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 467–491. ISBN 978-94-009-1453-7. [Google Scholar]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- NASEM. National Academies of Sciences, Engineering and Medicine. Nutrient Requirements of Beef Cattle; Eighth Revised Edition; The National Academies Press: Washington, DC, USA, 2016; ISBN 978-0-309-31702-3. [Google Scholar]
- Huntington, G.B. Starch utilization by ruminants: From basics to the bunk. J. Anim. Sci. 1997, 75, 852–867. [Google Scholar] [CrossRef]
- Wolin, M.J. A theoretical rumen fermentation balance. J. Dairy Sci. 1960, 43, 1452–1459. [Google Scholar] [CrossRef]
- Bell, N.L.; Anderson, R.C.; Callaway, T.R.; Franco, M.O.; Sawyer, J.E.; Wickersham, T.A. Effect of monensin inclusion on intake, digestion, and ruminal fermentation parameters by bos taurus indicus and bos taurus taurus steers consuming bermudagrass hay. J. Anim. Sci. 2017, 95, 2736–2746. [Google Scholar] [CrossRef]
- Rogers, J.A.; Davis, C.L. Rumen volatile fatty acid production and nutrient utilization in steers fed a diet supplemented with sodium bicarbonate and monensin. J. Dairy Sci. 1982, 65, 944–952. [Google Scholar] [CrossRef]
- Callaway, T.R.; Adams, K.A.; Russell, J.B. The ability of “low g + c gram-positive” ruminal bacteria to resist monensin and counteract potassium depletion. Curr. Microbiol. 1999, 39, 226–230. [Google Scholar] [CrossRef]
- Koike, S.; Kobayashi, Y. Fibrolytic rumen bacteria: Their ecology and functions. Asian Austral. J. Anim. 2009, 22, 131–138. [Google Scholar] [CrossRef]
- Appuhamy, J.A.D.R.N.; Strathe, A.B.; Jayasundara, S.; Wagner-Riddle, C.; Dijkstra, J.; France, J.; Kebreab, E. Anti-methanogenic effects of monensin in dairy and beef cattle: A meta-analysis. J. Dairy Sci. 2013, 96, 5161–5173. [Google Scholar] [CrossRef] [PubMed]
- Morvan, B.; Bonnemoy, F.; Fonty, G.; Gouet, P. Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and methanogenic archaea from digestive tract of different mammals. Curr. Microbiol. 1996, 32, 129–133. [Google Scholar] [CrossRef]
- Yang, C.M.J.; Russell, J.B. Effect of monensin on the specific activity of ammonia production by ruminal bacteria and disappearance of amino nitrogen from the rumen. Appl. Environ. Microbiol. 1993. [Google Scholar] [CrossRef] [PubMed]
- Chalupa, W.; Corbett, W.; Brethour, J.R. Effects of monensin and amicloral on rumen fermentation. J. Anim. Sci. 1980, 51, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Whetstone, H.D.; Davis, C.L.; Bryant, M.P. Effect of monensin on breakdown of protein by ruminant microorganisms in vitro. J. Anim. Sci. 1981, 53, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Paster, B.J.; Russell, J.B.; Yang, C.M.J.; Chow, J.M.; Woese, C.R.; Tanner, R.Y.R. Phylogeny of the ammonia-producing ruminal bacteria Peptostreptococcus Anaerobius, Clostridium Sticklandii, and Clostridium Aminophilum Sp. Nov. Int. J. Syst. Evol. Microbiol. 1993, 43, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Russell, J.B. Effect of monensin and a protonophore on protein degradation, peptide accumulation, and deamination by mixed ruminal microorganisms in vitro. J. Anim. Sci. 1991, 69, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Haïmoud, D.A.; Vernay, M.; Bayourthe, C.; Moncoulon, R. Avoparcin and monensin effects on the digestion of nutrients in dairy cows fed a mixed diet. Can. J. Anim. Sci. 1995, 75, 379–385. [Google Scholar] [CrossRef]
- Muntifering, R.B.; Theurer, B.; Noon, T.H. Effects of monensin on site and extent of whole corn digestion and bacterial protein synthesis in beef steers. J. Anim. Sci. 1981, 53, 1565–1573. [Google Scholar] [CrossRef]
- Faulkner, D.B.; Klopfenstein, T.J.; Trotter, T.N.; Britton, R.A. Monensin effects on digestibility, ruminal protein escape and microbial protein synthesis on high-fiber diets. J. Anim. Sci. 1985, 61, 654–660. [Google Scholar] [CrossRef]
- Russell, J.B.; Strobel, H.J.; Chen, G.J. Enrichment and isolation of a ruminal bacterium with a very high specific activity of ammonia production. Appl. Environ. Microbiol. 1988, 54, 872–877. [Google Scholar] [CrossRef]
- Argyle, J.L.; Baldwin, R.L. Effects of amino acids and peptides on rumen microbial growth yields. J. Dairy Sci. 1989, 72, 2017–2027. [Google Scholar] [CrossRef]
- Davenport, R.W.; Galyean, M.L.; Branine, M.E.; Hubbert, M.E. Effects of a monensin ruminal delivery device on daily gain, forage intake and ruminal fermentation of steers grazing irrigated winter wheat pasture. J. Anim. Sci. 1989, 67, 2129–2139. [Google Scholar] [CrossRef] [PubMed]
- Cappellozza, B.I.; Lasmar, P.V.F.; Reis, F.T.; Oliveira, L.; Hoe, F.; Boehler, R.M.; Leibovich, J.; Starkey, R.; Simas, J.; Cooke, R.F. Effects of supplement type and narasin inclusion on supplement intake by bos indicus beef bulls grazing a warm-season forage. Transl. Anim. Sci. 2019, 3, 263–273. [Google Scholar] [CrossRef]
- Horn, G.W. Growing Cattle on Winter Wheat Pasture: Management and herd health considerations. Vet. Clin. N. Am.-Food Anim. 2006, 22, 335–356. [Google Scholar] [CrossRef]
- Dawson, K.A.; Boling, J.A. Monensin-resistant bacteria in the rumens of calves on monensin-containing and unmedicated diets. Appl. Environ. Microbiol. 1983, 46, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Potchoiba, M.J.; Nocerini, M.R.; Carlson, J.R.; Breeze, R.G. Effect of energy or protein supplements containing monensin on ruminal 3-methylindole formation in pastured cattle. Am. J. Vet. Res. 1984, 45, 1389–1392. [Google Scholar]
- Pasqualino, L.F.; Oliveira, G.B.; Miszura, A.A.; Barroso, J.P.R.; Limede, A.C.; Sardinha, L.A.; Biava, J.S.; Ferreira, E.M.; Pires, A.V.; Polizel, D.M. Residual effect of narasin on ruminal fermentation characteristics in lambs. Livest. Sci. 2020, 240, 104141. [Google Scholar] [CrossRef]
- Davies, A.; Nwaonu, H.N.; Stanier, G.; Boyle, F.T. Properties of a novel series of inhibitors of rumen methanogenesis; in vitro and in vivo experiments including growth trials on 2,4-bis (Trichloromethyl)-Benzo [1,3]Dioxin-6-Carboxylic Acid. Brit. J. Nutr. 1982, 47, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.; Franz, C.M.A.P. Chapter 9 The farm animal as potential reservoir of antibiotic resistant bacteria in the food chain. In Biology of Growing Animals; Holzapfel, W.H., Naughton, P.J., Pierzynowski, S.G., Zabielski, R., Salek, E., Eds.; Microbial Ecology in Growing Animals; Elsevier: Amsterdam, The Netherlands, 2005; Volume 2, pp. 191–207. [Google Scholar]
- Odongo, N.E.; Bagg, R.; Vessie, G.; Dick, P.; Or-Rashid, M.M.; Hook, S.E.; Gray, J.T.; Kebreab, E.; France, J.; McBride, B.W. Long-term effects of feeding monensin on methane production in lactating dairy cows. J. Dairy Sci. 2007, 90, 1781–1788. [Google Scholar] [CrossRef] [PubMed]
- Mbanzamihigo, L.; van Nevel, C.; Demeyer, D. Essai sur l’adaptation de la fermentation ruminale au monensin. Reprod. Nutr. Dev. 1995, 35, 353–365. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guan, H.; Wittenberg, K.M.; Ominski, K.H.; Krause, D.O. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. 2006, 84, 1896–1906. [Google Scholar] [CrossRef]


{kind=link}
| Ionophore | Produced by | Molecular Weight | Ion-Binding Selectivity Sequence |
|---|---|---|---|
| Monensin | Streptomyces cinnamonensins | 671 | Na+ > K+, Li+ > Rb+ > Cs+ |
| Lasalocid | Streptomyces lasaliensis | 591 | Ba++, K+ >Rb+ > Na+ > Cs+ > Li+ |
| Narasin | Streptomyces aureofaciens | 765 | Na+ > K+, Rb+, Cs+, Li+ |
| Salinomycin | Streptomyces albus | 751 | Rb+, Na+ > K+ >> Cs+, Sr+, Ca++, Mg+ |
| Fermentation Products and Species | Gram Type Reaction | Sensitivity to Ionophores |
|---|---|---|
| Hydrogen and formic acid producers | ||
| Lachnospira multiparus | Gram+ | insensitive |
| Ruminococcus albus | Gram+ | insensitive |
| Ruminococcus flavefaciens | Gram+ | insensitive |
| Butyric acid producers | ||
| Butyvibrio fibrisolvens | Gram+ | insensitive |
| Eubacterium cellulosolvens | Gram+ | sensitive |
| Eubacterium ruminantium | Gram+ | sensitive |
| Lactic acid producers | ||
| Lactobacillus ruminis | Gram+ | sensitive |
| Lactobacillus vitulinis | Gram+ | sensitive |
| Streptococcus bovis | Gram+ | sensitive |
| Propionic and succinic acid producers | ||
| Anaerovibrio lipolytica | Gram− | insensitive |
| Fibrobacter succinogenes | Gram− | insensitive |
| Megasphaera elsdenii | Gram− | insensitive |
| Prevatella Ruminicola | Gram− | insensitive |
| Ruminobacter amylophilus | Gram− | insensitive |
| Selenomonas ruminantium | Gram− | insensitive |
| Succinimonas amylolytica | Gram− | insensitive |
| Succinivibrio dextrinosolvens | Gram− | insensitive |
| Ammonia producers | ||
| Clostridium aminophilum | Gram+ | Sensitive |
| Clostridium sticklandii | Gram+ | sensitive |
| Peptostreptococcus anaerobius | Gram+ | sensitive |
| Methane producers | ||
| Methanobrevibacter ruminantium | Gram− | insensitive |
| Methanobacterium formicum | Gram− | insensitive |
| Methanosrcina barkeri | Gram− | Insensitive |
| Item | Treatments | |||
|---|---|---|---|---|
| CON | NAR | SEM | p–Value | |
| Limede et al. [31] | ||||
| Acetate | 73.46 | 72.98 | 0.14 | <0.01 |
| Propionate | 13.77 | 14.53 | 0.14 | <0.01 |
| Butyrate | 9.05 | 8.60 | 0.10 | 0.01 |
| Acetate:Proprionate | 5.39 | 5.01 | 0.05 | <0.01 |
| Polizel et al. [33] | ||||
| Acetate | 74.21 | 72.71 | 0.16 | <0.01 |
| Propionate | 13.83 | 15.82 | 0.13 | <0.01 |
| Butyrate | 8.89 | 8.54 | 0.07 | <0.01 |
| Acetate:Proprionate | 5.40 | 4.63 | 0.04 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, R.d.S.; Cooke, R.F. Effects of Ionophores on Ruminal Function of Beef Cattle. Animals 2021, 11, 2871. https://doi.org/10.3390/ani11102871
Marques RdS, Cooke RF. Effects of Ionophores on Ruminal Function of Beef Cattle. Animals. 2021; 11(10):2871. https://doi.org/10.3390/ani11102871
Chicago/Turabian StyleMarques, Rodrigo da Silva, and Reinaldo Fernandes Cooke. 2021. "Effects of Ionophores on Ruminal Function of Beef Cattle" Animals 11, no. 10: 2871. https://doi.org/10.3390/ani11102871
APA StyleMarques, R. d. S., & Cooke, R. F. (2021). Effects of Ionophores on Ruminal Function of Beef Cattle. Animals, 11(10), 2871. https://doi.org/10.3390/ani11102871