Comparative Effects of Milk Containing A1 versus A2 β-Casein on Health, Growth and β-Casomorphin-7 Level in Plasma of Neonatal Dairy Calves

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Milk Protein Typing
2.2. Animals, Housing, and Feeding
2.3. Experimental Measurements
2.4. Blood Sampling
2.5. Analysis of β-Casomorphin-7 from Plasma
2.6. Phenotypic Data of the Calves
2.7. Statistical Analyses
2.7.1. Impact of Milk Feeding Group (A1- vs. A2-Milk) on Milk Intake and Health Status of Calves
2.7.2. Impact of Milk Feeding Group (A1- vs. A2-Milk) on Growth Traits of Calves
2.7.3. Quantification of β-Casomorphin-7 in Plasma
3. Results and Discussion
3.1. Allele and Genotype Frequencies at the β-Casein Locus
3.2. Impact of the Milk Feeding Group (A1- vs. A2-Milk) on Milk Intake and Growth Traits of Calves
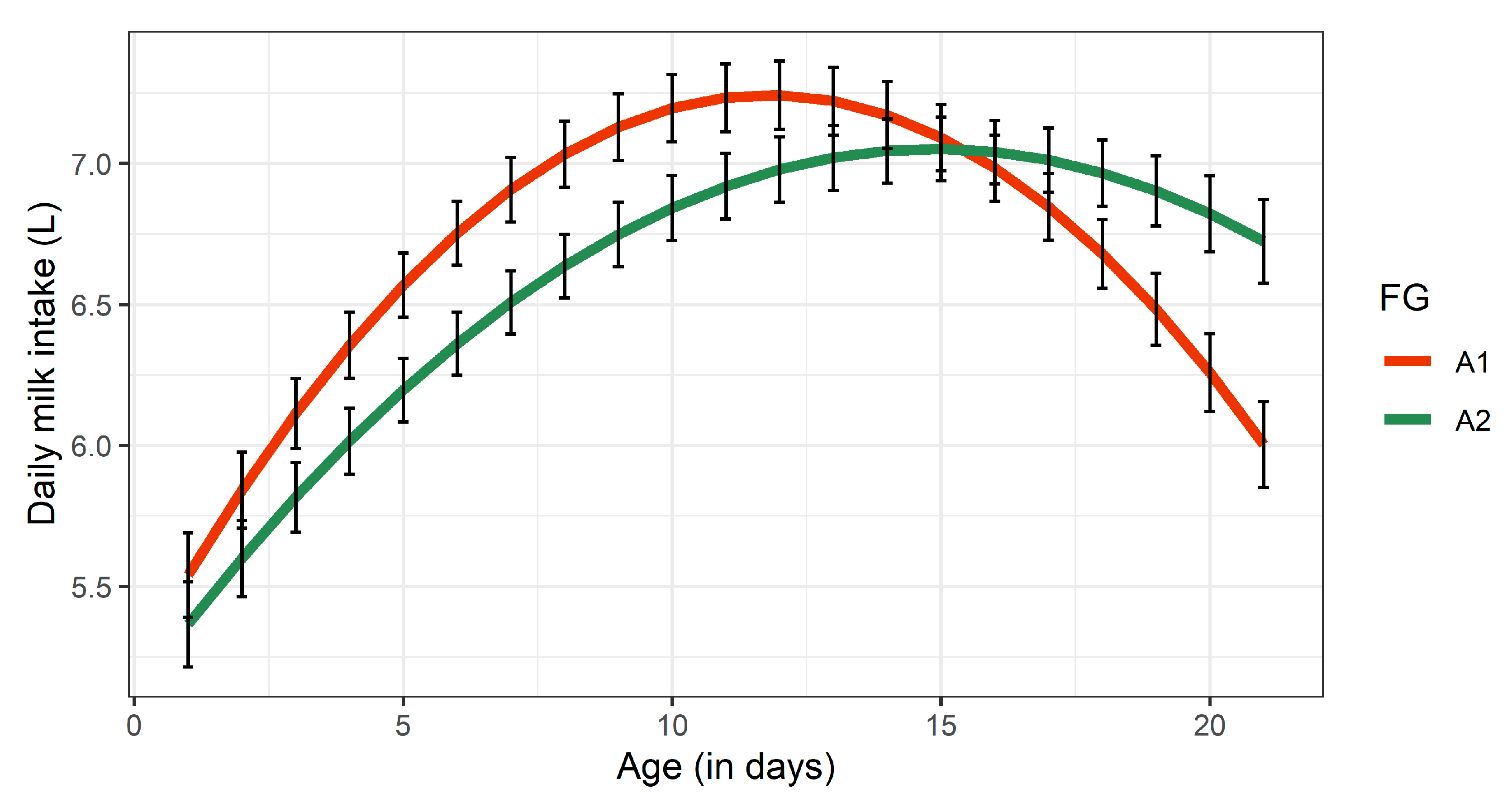
3.2.1. Daily Milk Intake
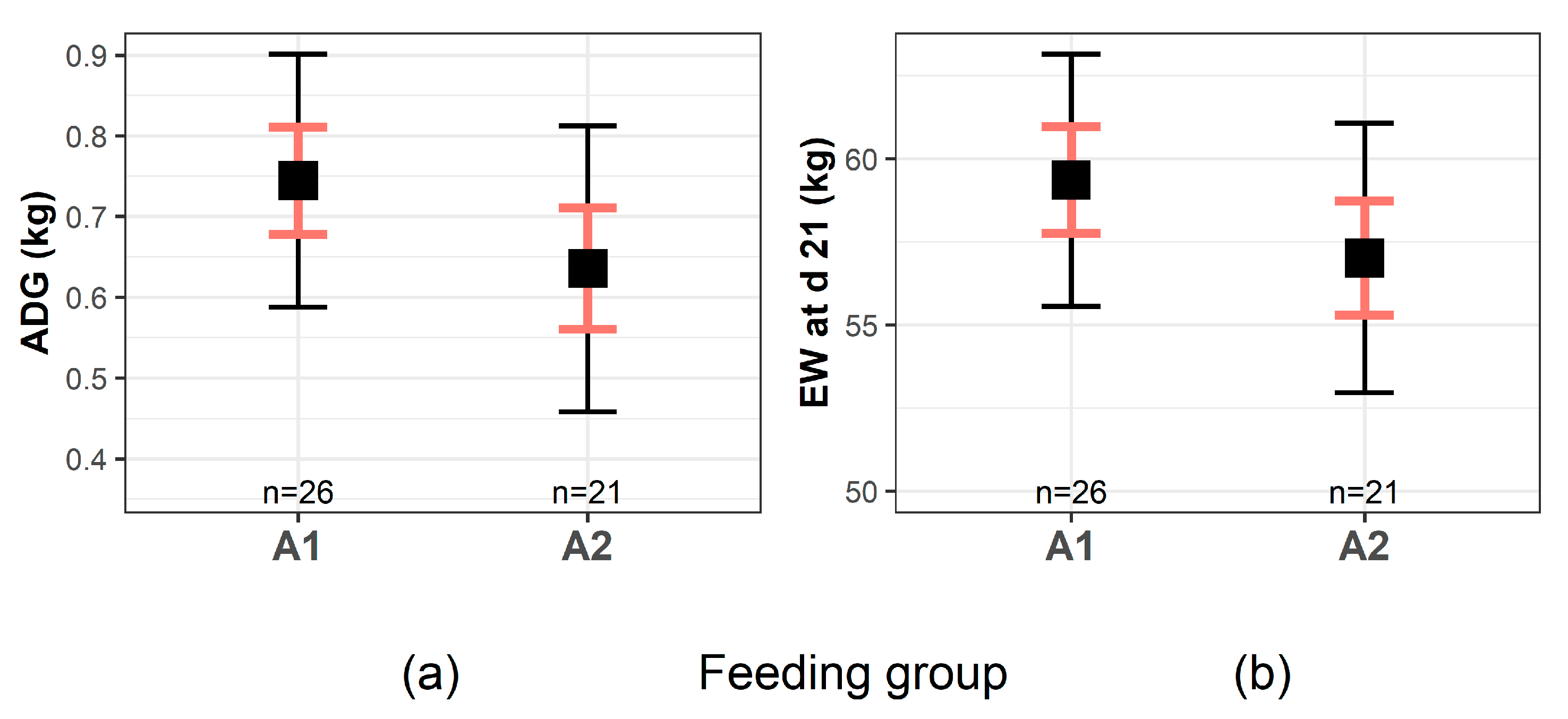
3.2.2. Growth Performance
3.3. Impact of the Milk Feeding Group (A1- vs. A2-Milk) on Health Parameters of Calves
3.3.1. Rectal Body Temperature
3.3.2. Respiratory Frequency
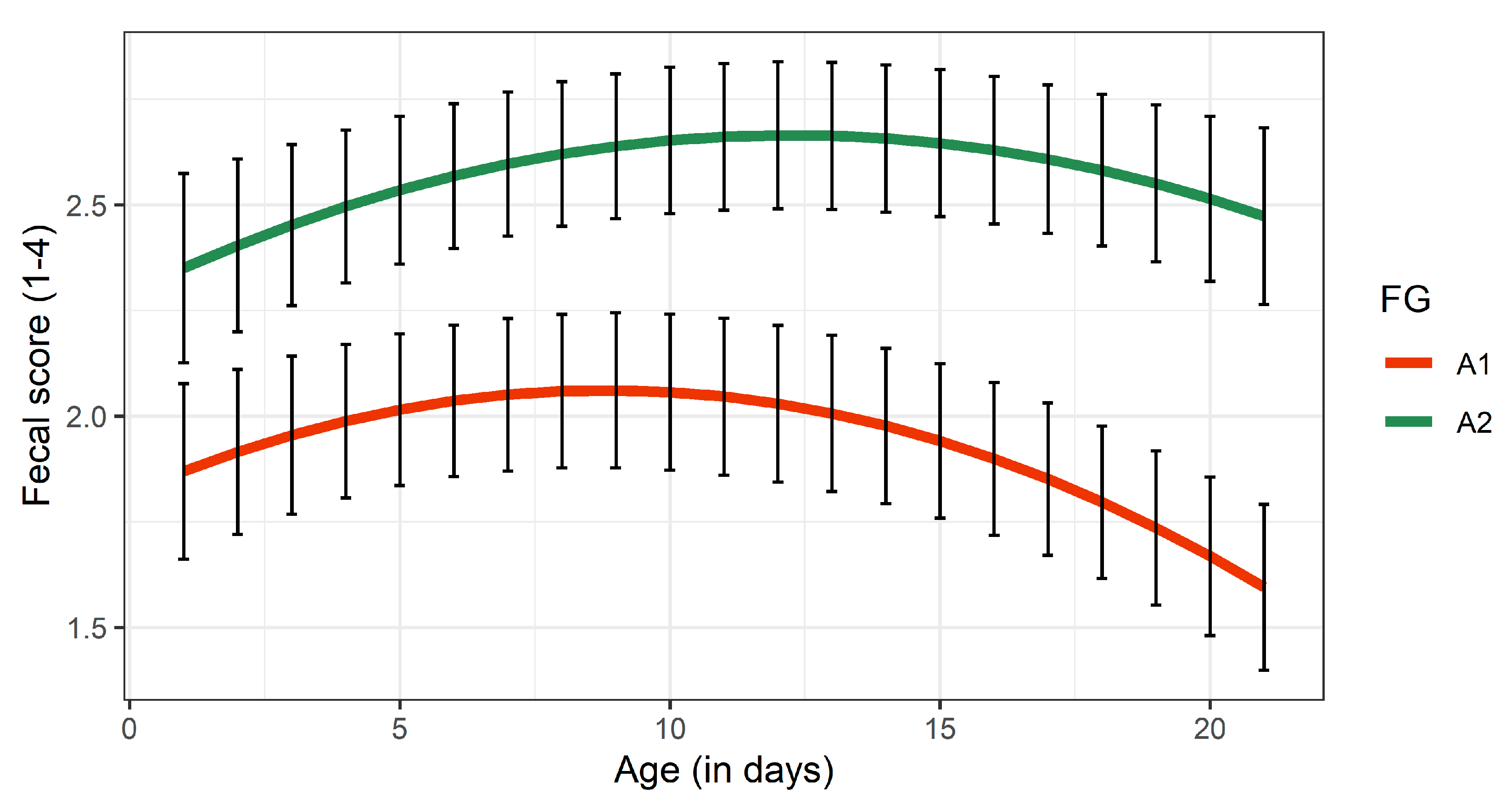
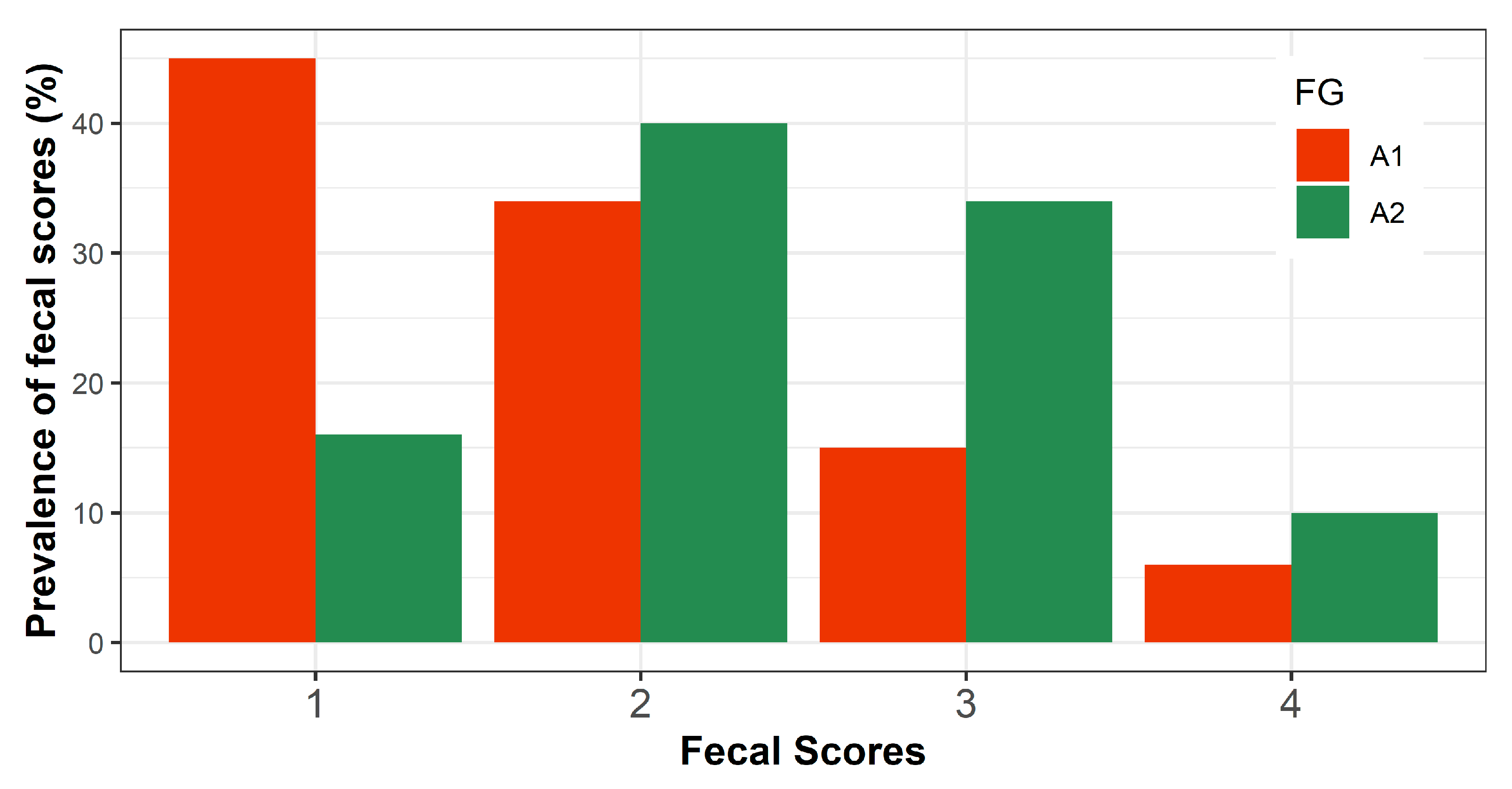
3.3.3. Fecal Consistency
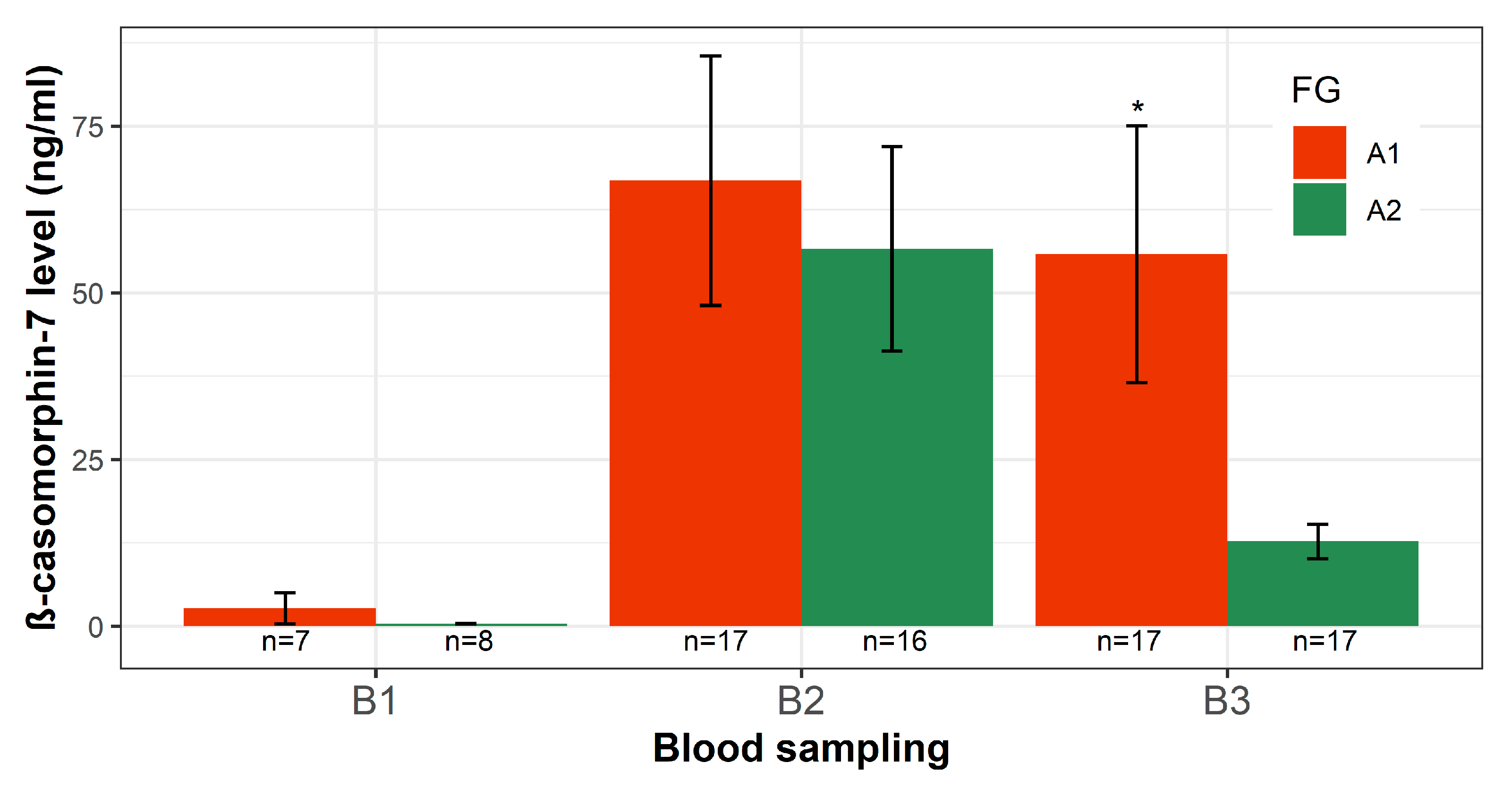
3.4. β-Casomorphin-7 Level in Plasma of Calves after A1- or A2-Milk Intake
3.5. Use of β-Casein Polymorphisms in Selection
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mee, J.F. Newborn Dairy Calf Management. Vet. Clin. North Am. Food Anim. Pr. 2008, 24, 1–17. [Google Scholar] [CrossRef]
- Schäff, C.T.; Gruse, J.; Maciej, J.; Mielenz, M.; Wirthgen, E.; Hoeflich, A.; Schmicke, M.; Pfuhl, R.; Jawor, P.; Stefaniak, T.; et al. Effects of Feeding Milk Replacer Ad Libitum or in Restricted Amounts for the First Five Weeks of Life on the Growth, Metabolic Adaptation, and Immune Status of Newborn Calves. PLoS ONE 2016, 11, e0168974. [Google Scholar] [CrossRef]
- Brown, E.; Vande-Haar, M.; Daniels, K.; Liesman, J.; Chapin, L.; Keisler, D.; Nielsen, M.W. Effect of Increasing Energy and Protein Intake on Body Growth and Carcass Composition of Heifer Calves. J. Dairy Sci. 2005, 88, 585–594. [Google Scholar] [CrossRef]
- Bennett, L.E.; Crittenden, R.; Khoo, E.; Forsyth, S. Evaluation of immune-modulatory properties of selected dairy peptide fractions. Aust. J. Dairy Technol. 2005, 60, 106–109. [Google Scholar]
- Gauthier, S.F.; Pouliot, Y.; Saint-Sauveur, D. Immunomodulatory peptides obtained by the enzymatic hydrolysis of whey proteins. Int. Dairy J. 2006, 16, 1315–1323. [Google Scholar] [CrossRef]
- López-Fandiño, R. Functional Improvement of Milk Whey Proteins Induced by High Hydrostatic Pressure. Crit. Rev. Food Sci. Nutr. 2006, 46, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Expósito, I.L.; Recio, I. Antibacterial activity of peptides and folding variants from milk proteins. Int. Dairy J. 2006, 16, 1294–1305. [Google Scholar] [CrossRef]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Meisel, H.; Fitzgerald, R.J. Opioid peptides encrypted in intact milk protein sequences. Br. J. Nutr. 2000, 84, 27–31. [Google Scholar] [CrossRef]
- Ng-Kwai-Hang, K.F.; Grosclaude, F. Genetic Polymorphism of Milk Proteins. In Advanced Dairy Chemistry-1 Proteins, 1st ed.; Fox, P.F., McSweeney, P.L.H., Eds.; Springer: Boston, MA, USA, 2003; Volume 1, pp. 739–816. ISBN 978-1-4419-8602-3. [Google Scholar]
- Damiani, G.; Pilla, F.; Leone, P.; Cacciò, S. Direct sequencing and bidirectional allelle specific polymerase chain reaction of the bovine β-casein B variant. Anim. Genet. 2009, 23, 561–566. [Google Scholar] [CrossRef]
- Cieślińska, A.; Kamiński, S.; Kostyra, E.; Sienkiewicz-Szłapka, E. Beta-casomorphin 7 in raw and hydrolyzed milk derived from cows of alternative beta-casein genotypes. Milchwissenschaft 2007, 62, 125–127. [Google Scholar]
- Tailford, K.A. A casein variant in cow’s milk is atherogenic. Atherosclerosis 2003, 170, 13–19. [Google Scholar] [CrossRef]
- Haq, M.R.U.; Kapila, R.; Sharma, R.; Saliganti, V.; Kapila, S. Comparative evaluation of cow β-casein variants (A1/A2) consumption on Th2-mediated inflammatory response in mouse gut. Eur. J. Nutr. 2013, 53, 1039–1049. [Google Scholar] [CrossRef]
- Barnett, M.P.G.; McNabb, W.C.; Roy, N.C.; Woodford, K.B.; Clarke, A.J. Dietary A1β-casein affects gastrointestinal transit time, dipeptidyl peptidase-4 activity, and inflammatory status relative to A2β-casein in Wistar rats. Int. J. Food Sci. Nutr. 2014, 65, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Hedner, J.; Hedner, T. β-casomorphins induce apnea and irregular breathing in adult rats and newborn rabbits. Life Sci. 1987, 41, 2303–2312. [Google Scholar] [CrossRef]
- Sun, J.; Leiming, X.; Lu, X.; Yelland, G.W.; Ni, J.; Clarke, A. Effects of milk containing only A2 beta casein versus milk containing both A1 and A2 beta casein proteins on gastrointestinal physiology, symptoms of discomfort, and cognitive behavior of people with self-reported intolerance to traditional cows’ milk. Nutr. J. 2015, 15, 1–16. [Google Scholar] [CrossRef]
- Daniel, H.; Vohwinkel, M.; Rehner, G. Effect of Casein and β-Casomorphins on Gastrointestinal Motility in Rats. J. Nutr. 1990, 120, 252–257. [Google Scholar] [CrossRef]
- Zoghbi, S.; Trompette, A.; Claustre, J.; el Homsi, M.; Garzón, J.; Jourdan, G.; Scoazec, J.-Y.; Plaisancié, P. β-Casomorphin-7 regulates the secretion and expression of gastrointestinal mucins through a μ-opioid pathway. Am. J. Physiol. Liver Physiol. 2006, 290, G1105–G1113. [Google Scholar] [CrossRef]
- Brandsch, M.; Brust, P.; Neubert, K.; Ermisch, A. systems. In β-Casomorphins and Related Peptides: Recent Developments, 1st ed.; Brantl, V., Teschemacher, H., Eds.; VCH: Weinheim, Germany, 1994; pp. 207–219. ISBN 978-3527300389. [Google Scholar]
- Daniel, H.; Wessendorf, A.; Vohwinkel, M.; Brantl, V. Effect of D-Ala2,4, Tyr5-β-casomorphin-5-amide on gastrointestinal functions. In β-Casomorphins and Related Peptides: Recent Developments, 1st ed.; Brantl, V., Teschemacher, H., Eds.; VCH: Weinheim, Germany, 1994; pp. 95–104. ISBN 978-3527300389. [Google Scholar]
- Ho, S.; Woodford, K.; Kukuljan, S.; Pal, S. Comparative effects of A1 versus A2 beta-casein on gastrointestinal measures: A blinded randomised cross-over pilot study. Eur. J. Clin. Nutr. 2014, 68, 994–1000. [Google Scholar] [CrossRef]
- Svedberg, J.; de Haas, J.; Leimenstoll, G.; Paul, F.; Teschemacher, H. Demonstration of β-casomorphin immunoreactive materials in in vitro digests of bovine milk and in small intestine contents after bovine milk ingestion in adult humans. Peptides 1985, 6, 825–830. [Google Scholar] [CrossRef]
- Bell, S.J.; Grochoski, G.T.; Clarke, A.J. Health Implications of Milk Containing β-Casein with the A2Genetic Variant. Crit. Rev. Food Sci. Nutr. 2006, 46, 93–100. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Review of the potential health impact of β-casomorphins and related peptides. EFSA J. 2009, 7, 1–107. [Google Scholar] [CrossRef]
- Umbach, M.; Teschemacher, H.; Praetorius, K.; Hirschhäuser, R.; Bostedt, H. Demonstration of a β-casomorphin immunoreactive material in the plasma of newborn calves after milk intake. Regul. Pept. 1985, 12, 223–230. [Google Scholar] [CrossRef]
- Singh, M.; Rosen, C.L.; Chang, K.J.; Haddad, G.G. Plasma? Casomorphin-7 Immunoreactive Peptide Increases after Milk Intake in Newborn but not in Adult Dogs. Pediatr. Res. 1989, 26, 34–38. [Google Scholar] [CrossRef]
- Kost, N.V.; Sokolov, O.Y.; Kurasova, O.B.; Dmitriev, A.D.; Tarakanova, J.N.; Gabaeva, M.V.; Zolotarev, Y.A.; Dadayan, A.K.; Grachev, S.A.; Korneeva, E.V.; et al. β-Casomorphins-7 in infants on different type of feeding and different levels of psychomotor development. Peptides 2009, 30, 1854–1860. [Google Scholar] [CrossRef]
- Seibert, B.; Erhardt, G.; Senft, B. Procedure for simultaneous phenotyping of genetic variants in cow’s milk by isoelectric focusing. Anim. Blood Groups Biochem. Genet. 2009, 16, 183–191. [Google Scholar] [CrossRef]
- Erhardt, G. K-Kaseine in Rindermilch—Nachweis eines weiteren Allels (k-CnE) in verschiedenen Rassen. J. Anim. Breed. Genet. 1989, 106, 225–231. [Google Scholar] [CrossRef]
- Wickramasinghe, H.; Kramer, A.; Appuhamy, J. Drinking water intake of newborn dairy calves and its effects on feed intake, growth performance, health status, and nutrient digestibility. J. Dairy Sci. 2019, 102, 377–387. [Google Scholar] [CrossRef]
- Kibler, H.H. Thermal effects of various temperature-humidity combinations on Holstein cattle as measured by eight physiological responses. Res. Bull. Missouri. Agric. Exp. Stn. 1964, 862, 142. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 10 November 2020).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.1-7. 2013. Available online: https://cran.r-project.org/ (accessed on 27 August 2020).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Meier, S.; Korkuć, P.; Arends, D.; Brockmann, G.A. DNA Sequence Variants and Protein Haplotypes of Casein Genes in German Black Pied Cattle (DSN). Front. Genet. 2019, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, T.; Ojala, M.; Ruottinen, O. Associations between Milk Protein Polymorphism and First Lactation Milk Production Traits in Finnish Ayrshire Cows. J. Dairy Sci. 1999, 82, 1026–1033. [Google Scholar] [CrossRef]
- Morrison, S.J.; Wicks, H.C.F.; Fallon, R.J.; Twigge, J.; Dawson, L.E.R.; Wylie, A.R.G.; Carson, A.F. Effects of feeding level and protein content of milk replacer on the performance of dairy herd replacements. Animal 2009, 3, 1570–1579. [Google Scholar] [CrossRef] [PubMed]
- Van Amburgh, M.; Tikofsky, J.; Smith, J. Requirements for and regulation of growth of Holstein calves—Implications for decreasing age at first calving. In Proceedings of the Tri-State Dairy Nutrition Conference, Fort Wayne, IN, USA, 17–18 April 2001; pp. 113–131. [Google Scholar]
- Sharma, B.; Nimje, P.; Tomar, S.; Dey, D.K.; Mondal, S.; Kundu, S.S. Effect of different fat and protein levels in calf ration on performance of Sahiwal calves. Asian-Australas. J. Anim. Sci. 2020, 33, 53–60. [Google Scholar] [CrossRef]
- Dhakal, K.; Maltecca, C.; Cassady, J.; Baloche, G.; Williams, C.; Washburn, S. Calf birth weight, gestation length, calving ease, and neonatal calf mortality in Holstein, Jersey, and crossbred cows in a pasture system. J. Dairy Sci. 2013, 96, 690–698. [Google Scholar] [CrossRef]
- Kertz, A.; Reutzel, L.; Barton, B.; Ely, R. Body Weight, Body Condition Score, and Wither Height of Prepartum Holstein Cows and Birth Weight and Sex of Calves by Parity: A Database and Summary. J. Dairy Sci. 1997, 80, 525–529. [Google Scholar] [CrossRef]
- Finch, V.A. Body Temperature in Beef Cattle: Its Control and Relevance to Production in the Tropics. J. Anim. Sci. 1986, 62, 531–542. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Wang, F.; Xiao, J.; Wang, Y.; Yang, H.; Li, S.; Cao, Z. Heat stress on calves and heifers: A review. J. Anim. Sci. Biotechnol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Beakley, W.R.; Findlay, J.D. The effect of environmental temperature and humidity on the skin temperature of Ayrshire calves. J. Agric. Sci. 1955, 45, 353–364. [Google Scholar] [CrossRef]
- Kovács, L.; Kézér, F.; Póti, P.; Boros, N.; Nagy, K. Short communication: Upper critical temperature-humidity index for dairy calves based on physiological stress variables. J. Dairy Sci. 2020, 103, 2707–2710. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.T.; Henklein, A.; de Sousa, R.M.; de Oliveira, P.L.; Leite, S.B.P.; Novo, S.M.F.; Baccili, C.C.; Reis, J.F.; Gomes, V. Vital parameters of Holstein calves from birth to weaning. Rev. Bras. Med. Vet. 2016, 38, 299–304. [Google Scholar]
- Piccione, G.; Casella, S.; Pennisi, P.; Giannetto, C.; Costa, A.; Caola, G. Monitoring of physiological and blood parameters during perinatal and neonatal period in calves. Arqu. Brasil. Med. Vet. Zootec. 2010, 62, 1–12. [Google Scholar] [CrossRef]
- Wasilewska, J.; Kaczmarski, M.; Kostyra, E.; Iwan, M. Cow’s-milk–induced Infant Apnoea with Increased Serum Content of Bovine β-Casomorphin-5. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 772–775. [Google Scholar] [CrossRef]
- Linke, B.; Bostedt, H.; Richter, A. Computer Tomographic Illustration of the Development of the Pulmonary Function in Bovine Neonates until the Twenty-First Day Postnatum. Vet. Med. Int. 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Findlay, J.D. The respiratory activity of calves subjected to thermal stress. J. Physiol. 1957, 136, 300–309. [Google Scholar] [CrossRef]
- Gantner, V.; Bobic, T.; Gantner, R.; Gregic, M.; Kuterovac, K.; Novakovic, J.; Potocnik, K. Differences in response to heat stress due to production level and breed of dairy cows. Int. J. Biometeorol. 2017, 61, 1675–1685. [Google Scholar] [CrossRef]
- Curtis, C.R.; Erb, H.N.; White, M.E. Descriptive epidemiology of calfhood morbidity and mortality in New York holstein herds. Prev. Vet. Med. 1988, 5, 293–307. [Google Scholar] [CrossRef]
- Mihatsch, W.A.; Franz, A.R.; Kuhnt, B.; Pohlandt, F.; Högel, J. Hydrolysis of Casein Accelerates Gastrointestinal Transit via Reduction of Opioid Receptor Agonists Released from Casein in Rats. Biol. Neonatol. 2005, 87, 160–163. [Google Scholar] [CrossRef]
- Tome, D.; Dumontier, A.M.; Hautefeuille, M.; Desjeux, J.F. Opiate activity and transepithelial passage of intact beta-casomorphins in rabbit ileum. Am. J. Physiol. Liver Physiol. 1987, 253, G737–G744. [Google Scholar] [CrossRef]
- Britton, J.R.; Koldovsky, O. Gastric luminal digestion of lactoferrin and transferrin by preterm infants. Early Hum. Dev. 1989, 19, 127–135. [Google Scholar] [CrossRef]
- Cattaneo, S. Release of β-casomorphins 5 and 7 during simulated gastro-intestinal digestion of bovine β-casein variants and milk-based infant formulas. Food Chem. 2008, 110, 897–903. [Google Scholar] [CrossRef]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic release of neocasomorphin and β-casomorphin from bovine β-casein. Peptide 1999, 20, 957–962. [Google Scholar] [CrossRef]
- Asledottir, T.; Le, T.T.; Petrat-Melin, B.; Devold, T.G.; Larsen, L.B.; Vegarud, G.E. Identification of bioactive peptides and quantification of β-casomorphin-7 from bovine β-casein A1, A2 and I after ex vivo gastrointestinal digestion. Int. Dairy J. 2017, 71, 98–106. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Solah, V.A.; Johnson, S.K.; Nguyen, H.A.; Nguyen, T.L.D.; Tran, T.L.H.; Mai, T.K.; Busetti, F. Identification and quantification of beta-casomorphin peptides naturally yielded in raw milk by liquid chromatography-tandem mass spectrometry. LWT 2019, 111, 465–469. [Google Scholar] [CrossRef]
- Rincker, L.D.; van de Haar, M.; Wolf, C.; Liesman, J.; Chapin, L.; Nielsen, M.W. Effect of intensified feeding of heifer calves on growth, pubertal age, calving age, milk yield, and economics. J. Dairy Sci. 2011, 94, 3554–3567. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BW (kg) | ADG (kg/day) | EW (kg) | dMI (L) | BT (°C) | RF (bpm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| GH | 9 | 44.30 | 3.05 | 0.71 | 0.22 | 59.72 | 4.77 | 6.78 | 0.79 | 38.8 | 0.51 | 51.6 | 23.20 |
| GS | 33 | 43.70 | 6.51 | 0.65 | 0.16 | 57.81 | 7.70 | 6.53 | 1.09 | 38.9 | 0.41 | 44.4 | 18.00 |
| GH × GS | 5 | 41.11 | 6.37 | 0.65 | 0.19 | 55.21 | 3.77 | 6.64 | 1.10 | 38.8 | 0.46 | 49.6 | 19.80 |
| Allele Frequencies | Genotype Frequencies | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| n | A1 | A2 | B | A1A1 | A1A2 | A2A2 | A1B | A2B | |
| GS | 41 | 0.400 | 0.575 | 0.025 | 0.050 | 0.700 | 0.200 | 0.000 | 0.050 |
| GH | 8 | 0.500 | 0.500 | 0.000 | 0.125 | 0.750 | 0.125 | 0.000 | 0.000 |
| GH × GS | 62 | 0.492 | 0.475 | 0.033 | 0.164 | 0.590 | 0.180 | 0.066 | 0.000 |
| Total | 111 | 0.465 | 0.516 | 0.019 | 0.114 | 0.680 | 0.168 | 0.022 | 0.016 |
| Feeding Group | p-Value | t-Test-Statistic | |||
|---|---|---|---|---|---|
| A1-Milk | A2-Milk | ||||
| Growth | dMI (L) | 7.28 ± 0.12 a | 6.96 ± 0.11 b | 0.02 | 2.38 |
| ADG (kg/day) | 0.75 ± 0.07 | 0.64 ± 0.08 | 0.07 | 1.87 | |
| EW (kg) | 59.40 ± 1.61 | 57.0 ± 1.73 | 0.07 | 0.76 | |
| Health | BT (°C) | 38.94 ± 0.06 | 38.96 ± 0.05 | 0.75 | −0.32 |
| RF (bpm) | 46.60 ± 3.64 | 53.10 ± 3.50 | 0.13 | −1.56 | |
| FS | 1.97 ± 0.18 a | 2.56 ± 0.17 b | <0.001 | −3.86 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hohmann, L.G.; Yin, T.; Schweizer, H.; Giambra, I.J.; König, S.; Scholz, A.M. Comparative Effects of Milk Containing A1 versus A2 β-Casein on Health, Growth and β-Casomorphin-7 Level in Plasma of Neonatal Dairy Calves. Animals 2021, 11, 55. https://doi.org/10.3390/ani11010055
Hohmann LG, Yin T, Schweizer H, Giambra IJ, König S, Scholz AM. Comparative Effects of Milk Containing A1 versus A2 β-Casein on Health, Growth and β-Casomorphin-7 Level in Plasma of Neonatal Dairy Calves. Animals. 2021; 11(1):55. https://doi.org/10.3390/ani11010055
Chicago/Turabian StyleHohmann, Lisa G., Tong Yin, Helen Schweizer, Isabella J. Giambra, Sven König, and Armin M. Scholz. 2021. "Comparative Effects of Milk Containing A1 versus A2 β-Casein on Health, Growth and β-Casomorphin-7 Level in Plasma of Neonatal Dairy Calves" Animals 11, no. 1: 55. https://doi.org/10.3390/ani11010055
APA StyleHohmann, L. G., Yin, T., Schweizer, H., Giambra, I. J., König, S., & Scholz, A. M. (2021). Comparative Effects of Milk Containing A1 versus A2 β-Casein on Health, Growth and β-Casomorphin-7 Level in Plasma of Neonatal Dairy Calves. Animals, 11(1), 55. https://doi.org/10.3390/ani11010055