Association between Single Nucleotide Polymorphisms in SIRT1 and SIRT2 Loci and Growth in Tibetan Sheep


 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
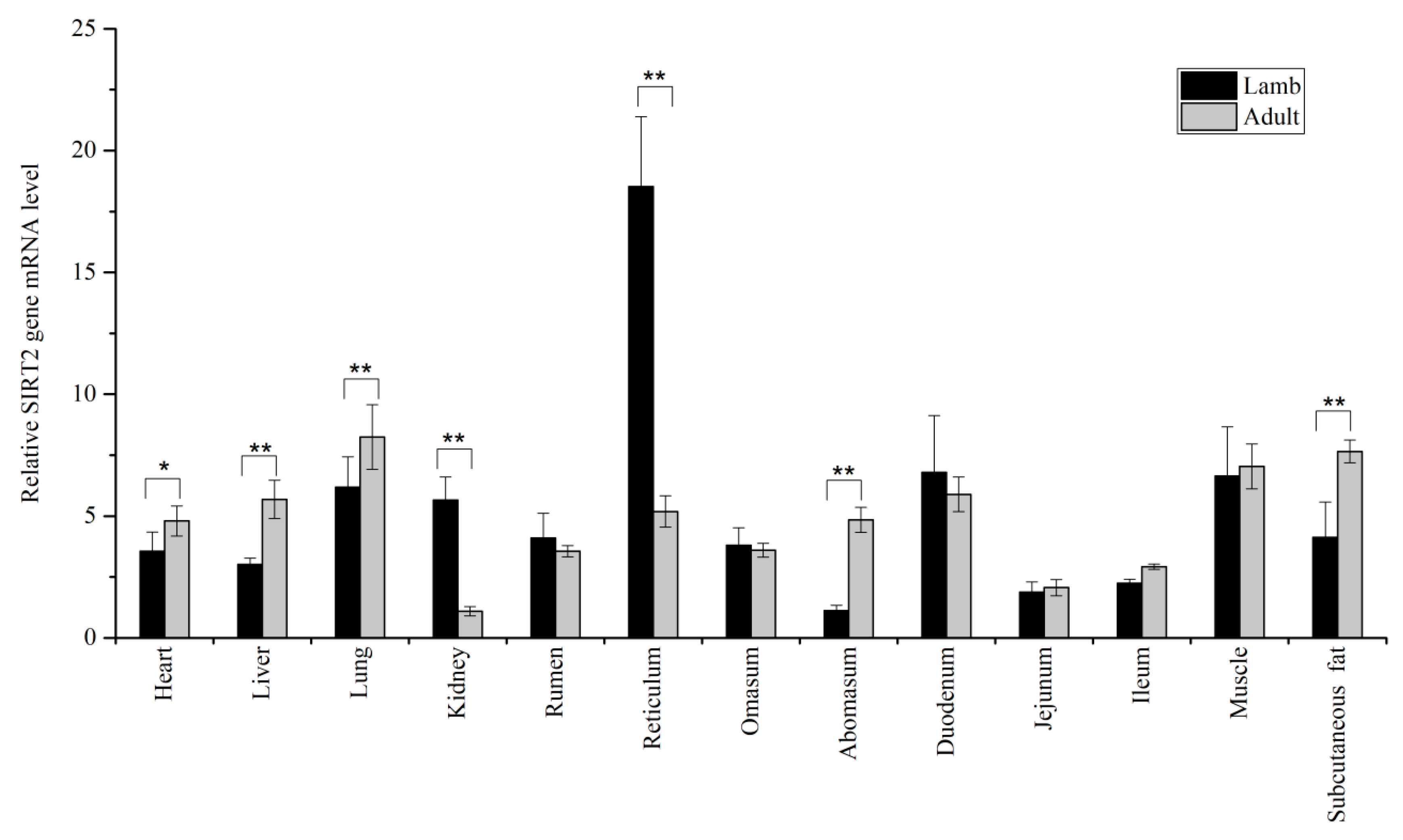
3.1. Expression Levels of SIRT1 and SIRT2 in Tissues
3.2. Polymorphisms and Genetic Diversity
3.3. LD Analysis
3.4. Association Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.Q.; Dong, Z.; Ke, X.X.; Hou, J.B.; Zhao, E.H.; Zhang, K.; Wang, F.; Yang, L.Q.; Xiang, Z.H.; Cui, H.J. The roles of sirtuins family in cell metabolism during tumor development. Semin. Cancer Biol. 2019, 57, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Wilking, M.J.; Singh, C.; Nihal, M.; Zhong, W.X.; Ahmad, N. SIRT1 deacetylase is overexpressed in human melanoma and its small molecule inhibition imparts anti-proliferative response via p53 activation. Arch. Biochem. Biophys. 2014, 563, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Kikuchi, O.; Shimpuku, M.; Susanti, V.Y.; Yokota-Hashimoto, H.; Taguchi, R.; Shibusawa, N.; Sato, T.; Tang, L.J.; Amano, K.; et al. Hypothalamic SIRT1 prevents age-associated weight gain by improving leptin sensitivity in mice. Diabetologia 2014, 57, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Zheng, X.B.; Lin, B.S.; Liang, H.; Cai, M.Y.; Cao, H.Y.; Ye, J.P.; Weng, J.P. Diet-Induced Obesity and Insulin Resistance are Associated with Brown Fat Degeneration in SIRT1-Deficient Mice. Obesity 2016, 24, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Erburu, M.; Munoz-Cobo, I.; Dominguez-Andres, J.; Beltran, E.; Suzuki, T.; Mai, A.; Valente, S.; Puerta, E.; Tordera, R.M. Chronic stress and antidepressant induced changes in Hdac5 and Sirt2 affect synaptic plasticity. Eur. Neuropsychopharmacol. 2015, 25, 2036–2048. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Dey, C.S. SIRT2 negatively regulates insulin resistance in C2C12 skeletal muscle cells. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Zhao, Z.G.; Tang, D.H.; Zhou, Q.; Li, Y.; Zhou, L.X.; Yin, Y.Y.; Wang, Y.M.; Pan, Y.D.; Dorfman, R.G.; et al. Downregulation of SIRT2 Inhibits Invasion of Hepatocellular Carcinoma by Inhibiting Energy Metabolism. Transl. Oncol. 2017, 10, 917–927. [Google Scholar] [CrossRef]
- Lantier, L.; Williams, A.S.; Hughey, C.C.; Bracy, D.P.; James, F.D.; Ansari, M.A.; Gius, D.; Wasserman, D.H. SIRT2 knockout exacerbates insulin resistance in high fat-fed mice. PLoS ONE 2018, 13, e0208634. [Google Scholar] [CrossRef]
- Fourcade, S.; Morato, L.; Parameswaran, J.; Ruiz, M.; Ruiz-Cortes, T.; Jove, M.; Naudi, A.; Martinez-Redondo, P.; Dierssen, M.; Ferrer, I.; et al. Loss of SIRT2 leads to axonal degeneration and locomotor disability associated with redox and energy imbalance. Aging Cell 2017, 16, 1404–1413. [Google Scholar] [CrossRef]
- Ghinis-Hozumi, Y.; Antaramian, A.; Villarroya, F.; Pina, E.; Mora, O. Potential role of sirtuins in livestock production. Animal 2013, 7, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.G.; Han, Y.C. Genetic Polymorphisms of Myogenin Gene and Their Associations with Growth Traits in the Chinese Tibetan Sheep. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 253–258. [Google Scholar]
- Wang, L.; Zhang, K.; Zhang, C.G.; Feng, Y.Z.; Zhang, X.W.; Wang, X.L.; Wu, G.F. Dynamics and stabilization of the rumen microbiome in yearling Tibetan sheep. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, L.S.; Jiang, Y.Y.; Liu, G.Y.; Jia, J.L.; Chu, M.D.; Hou, S.Z.; Wang, Z.Y. Polymorphism in GHRH Gene and Its Association with Growth Traits in Tibetan Sheep. Kafkas Universitesi Veteriner Fakultesi Dergisi 2020, 26, 151–155. [Google Scholar]
- Balestrieri, M.L.; Rienzo, M.; Felice, F.; Rossiello, R.; Grimaldi, V.; Milone, L.; Casamassimi, A.; Servillo, L.; Farzati, B.; Giovane, A.; et al. High glucose downregulates endothelial progenitor cell number via SIRT1. Biochim. Et Biophys. Acta-Proteins Proteom. 2008, 1784, 936–945. [Google Scholar] [CrossRef]
- Figarska, S.M.; Vonk, J.M.; Boezen, H.M. SIRT1 Polymorphism, Long-Term Survival and Glucose Tolerance in the General Population. PLoS ONE 2013, 8, e58636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, L.S.; Wang, H.B.; Wei, S.J.; Zhang, Y.R.; Zan, L.S. Molecular characterization, expression profiles, and analysis of Qinchuan cattle SIRT1 gene association with meat quality and body measurement traits (Bos taurus). Mol. Biol. Rep. 2014, 41, 5237–5246. [Google Scholar] [CrossRef]
- Ghinis-Hozumi, Y.; Gonzalez-Gallardo, A.; Gonzalez-Davalos, L.; Antaramian, A.; Villarroya, F.; Shimada, A.; Varela-Echavarria, A.; Mora, O. Bovine sirtuins: Initial characterization and expression of sirtuins 1 and 3 in liver, muscle, and adipose tissue. J. Anim. Sci. 2011, 89, 2529–2536. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.B.; Ma, Y.; Yang, T.; Lin, F.; Fu, W.W.; Xu, Y.J. Tissue expression and predicted protein structures of the bovine ANGPTL3 and association of novel SNPs with growth and meat quality traits. Animal 2015, 9, 1285–1297. [Google Scholar] [CrossRef]
- Le Hir, H.; Nott, A.; Moore, M.J. How introns influence and enhance eukaryotic gene expression. Trends Biochem. Sci. 2003, 28, 215–220. [Google Scholar] [CrossRef]
- Chai, Y.R.; Sun, Y.; Guo, L.X.; Li, D.; Ding, Y. Investigating the role of introns in the regulation of regenerating gene 1 expression. Oncol. Lett. 2014, 9, 875–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, T.; Sano, R.; Takahashi, Y.; Watanabe, K.; Kubo, R.; Kobayashi, M.; Takahashi, K.; Takeshita, H.; Kominato, Y. ABO alleles are linked with haplotypes of an erythroid cell-specific regulatory element in intron 1 with a few exceptions attributable to genetic recombination. Vox Sang. 2016, 110, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Thomsen, B.; Bai, J.; Peng, S.; Lan, X.; Huang, Y.; Wang, X.; Chen, H. Expression profiles of the MXD3 gene and association of sequence variants with growth traits in Xianan and Qinchuan cattle. Vet. Med. Sci. 2020, 6, 399–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, L.S.; Huang, Y.M.; Hong, J.Y.; Zan, L.S. Polymorphisms in the LPL gene and their association with growth traits in Jiaxian cattle. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Sun, X.M.; Li, M.X.; Hao, D.; Hua, L.S.; Lan, X.Y.; Lei, C.Z.; Hu, S.R.; Qi, X.L.; Chen, H. Two novel polymorphisms of bovine SIRT2 gene are associated with higher body weight in Nanyang cattle. Mol. Biol. Rep. 2015, 42, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.S.; Hao, R.J.; Zhang, Y.R.; Zhao, X.L.; Zan, L.S. Haplotype distribution in the class I sirtuin genes and their associations with ultrasound carcass traits in Qinchuan cattle (Bos taurus). Mol. Cell. Probes 2015, 29, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.Y.; Yang, Q.; Gao, J.Y.; Tang, Q.; Duan, B.; Yu, T.; Qi, X.L.; Liu, J.M.; Wang, R.M.; Dang, R.H.; et al. Detection of Insertions/Deletions Within SIRT1, SIRT2 and SIRT3 Genes and Their Associations with Body Measurement Traits in Cattle. Biochem. Genet. 2018, 56, 663–676. [Google Scholar] [CrossRef]
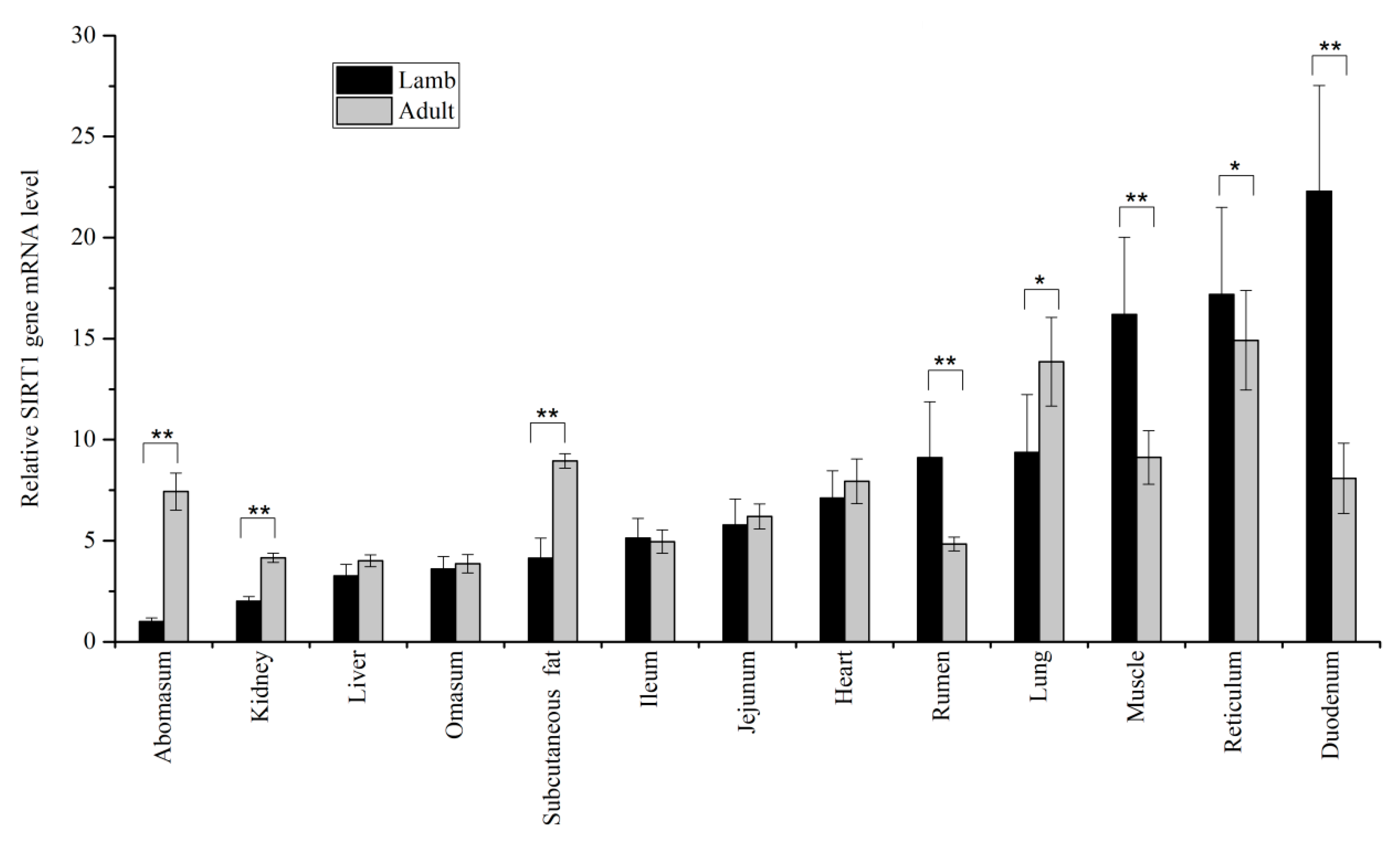
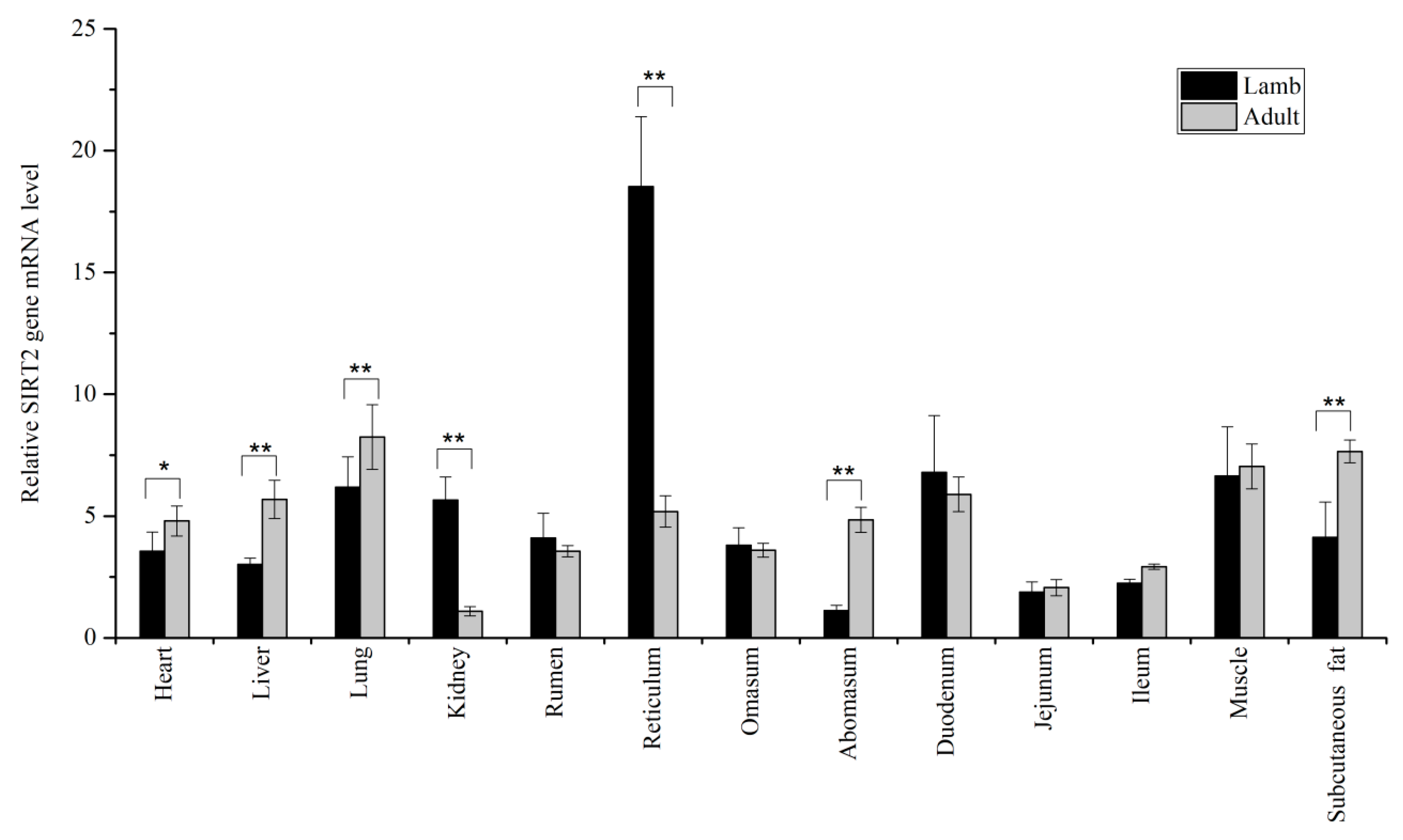

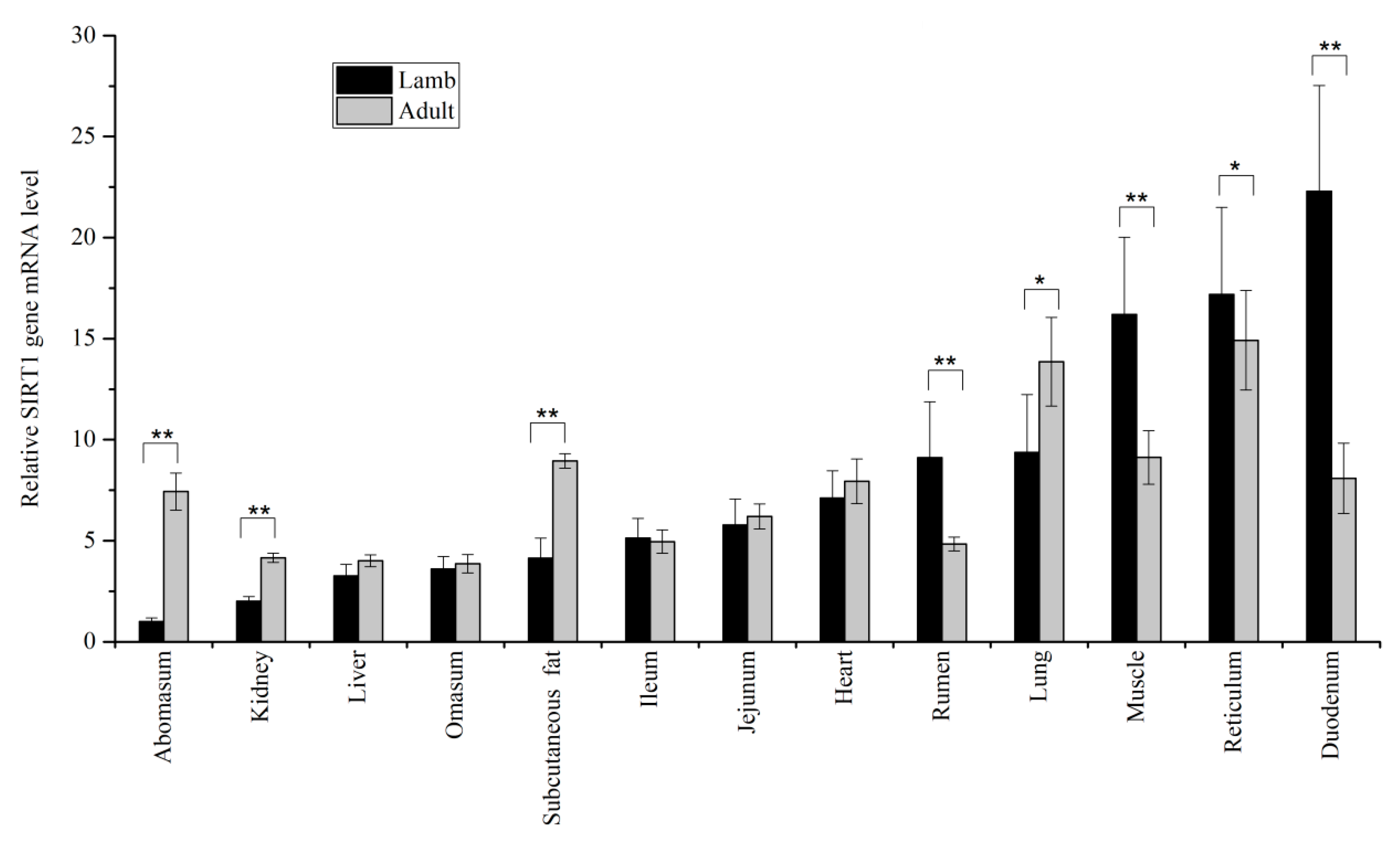{kind=link}
{kind=link}
| Name | Primer Sequence (5’ to 3’) | Tm (°C) | Product Location | Product Length |
|---|---|---|---|---|
| SIRT1-SNP | CCTTAGCTCTGAAGATGTTCT | 55.7 | Part of intron 2, exon 3, and part of intron 3 | 629 bp |
| TACTGTGTCAATAAATAGAA | ||||
| SIRT2-SNP | TTTTGGAGGAATGTTTGATG | 57.0 | Part of exon 5, intron 5, exon 6, intron 6, exon 7, and part of intron 7 | 764 bp |
| ACTATGCTTCAAGGCTTGCA | ||||
| SIRT1-qPCR | ACTTCTACGACGACGACGACGAG | 61.0 | - | 182 bp |
| CCGAGGTCTTGGAGTCCAGTCAC | ||||
| SIRT2-qPCR | TGGCGGAGAAGCAGAGATGGAC | 61.0 | - | 116 bp |
| TGTAGCGGCTCACTCCTTCCAG | ||||
| β-actin-qPCR | CGTCCGTGACATCAAGGAGAAGC | 61.0 | - | 143 bp |
| GGAACCGCTCATTGCCGATGG | ||||
| GAPD-qPCR | CGGCACAGTCAAGGCAGAGAAC | 61.0 | - | 115 bp |
| CACGTACTCAGCACCAGCATCAC |
| Loci (Gene) | Genotypic Frequency (%) | PIC | Maximum Allele Frequency (%) | HWE | ||
|---|---|---|---|---|---|---|
| g.3148 C > T (SIRT1) | CC | CT | TT | |||
| 58.46 | 34.08 | 7.46 | 0.30 | 75.50 (C) | p > 0.05 | |
| g.3570 G > A (SIRT1) | GG | GA | AA | |||
| 67.16 | 27.36 | 5.48 | 0.26 | 80.85 (G) | p > 0.05 | |
| g.8074 T > A (SIRT2) | TT | TA | AA | |||
| 72.39 | 23.88 | 3.73 | 0.23 | 84.33 (T) | p > 0.05 | |
| Haplotype | g.3148 C > T | g.3570 G > A | Frequency |
|---|---|---|---|
| Hap1 | C | A | 13.30% |
| Hap2 | C | G | 62.20% |
| Hap3 | T | A | 5.90% |
| Hap4 | T | G | 18.60% |
| Loci (Gene) | Genotypes (N) | Body Weight (kg) | Withers Height (cm) | Body Length (cm) | Heart Girth (cm) |
|---|---|---|---|---|---|
| g.3148 C > T (SIRT1) | CC (235) | 58.63 ± 0.46 | 71.60 ± 0.82 | 75.00 ± 0.65 | 96.08 ± 1.03 b |
| CT (137) | 56.24 ± 0.59 | 70.54 ± 0.57 | 73.83 ± 0.54 | 96.73 ± 0.79 b | |
| TT (30) | 54.48 ± 0.79 | 70.78 ± 0.68 | 72.51 ± 0.77 | 88.38 ± 0.85 a | |
| p1 | 0.072 | 0.253 | 0.109 | 0.002 | |
| g.3570 G > A (SIRT1) | GG (270) | 58.54 ± 0.65 | 71.62 ± 0.76 | 75.03 ± 0.82 | 96.77 ± 0.88 |
| GA (110) | 55.88 ± 0.48 | 70.32 ± 0.59 | 73.39 ± 0.69 | 94.15 ± 0.72 | |
| AA (22) | 52.94 ± 0.72 | 70.01 ± 0.60 | 72.02 ± 0.66 | 90.78 ± 0.99 | |
| p1 | 0.038 | 0.157 | 0.070 | 0.019 | |
| g.8074 T>A (SIRT2) | TT (291) | 57.53 ± 0.27 b | 71.17 ± 0.20 | 74.44 ± 0.19 b | 96.03 ± 0.29 |
| TA (96) | 56.09 ± 0.47 b | 70.41 ± 0.35 | 73.23 ± 0.33 b | 94.43 ± 0.53 | |
| AA (15) | 66.07 ± 0.92 a | 76.22 ± 0.89 | 81.63 ± 0.83 a | 98.00 ± 1.17 | |
| p2 | 0.011 | 0.031 | 0.008 | 0.074 |
| Genotypes (N) | Body Weight (kg) | Withers Height (cm) | Body Length (cm) | Heart Girth (cm) |
|---|---|---|---|---|
| Hap2/1 (63) | 56.84 ± 0.44 | 70.59 ± 0.63 | 74.03 ± 0.45 | 95.52 ± 0.73 |
| Hap2/2 (159) | 59.72 ± 0.67 | 72.08 ± 0.70 | 75.59 ± 0.43 | 96.63 ± 1.23 |
| Hap2/3 (34) | 55.61 ± 0.48 | 70.12 ± 0.79 | 73.17 ± 0.57 | 94.60 ± 0.88 |
| Hap2/4 (99) | 56.50 ± 0.40 | 70.64 ± 0.53 | 74.03 ± 0.61 | 97.67 ± 0.74 |
| p1 | 0.032 | 0.225 | 0.438 | 0.040 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gui, L.-s.; Raza, S.H.A.; Zhou, L.; Garcia, M.; Abd El-Aziz, A.H.; Wei, D.; Hou, S.; Jia, J.; Wang, Z. Association between Single Nucleotide Polymorphisms in SIRT1 and SIRT2 Loci and Growth in Tibetan Sheep. Animals 2020, 10, 1362. https://doi.org/10.3390/ani10081362
Gui L-s, Raza SHA, Zhou L, Garcia M, Abd El-Aziz AH, Wei D, Hou S, Jia J, Wang Z. Association between Single Nucleotide Polymorphisms in SIRT1 and SIRT2 Loci and Growth in Tibetan Sheep. Animals. 2020; 10(8):1362. https://doi.org/10.3390/ani10081362
Chicago/Turabian StyleGui, Lin-sheng, Sayed Haidar Abbas Raza, Li Zhou, Matthew Garcia, Ayman Hassan Abd El-Aziz, Dawei Wei, Shengzhen Hou, Jianlei Jia, and Zhiyou Wang. 2020. "Association between Single Nucleotide Polymorphisms in SIRT1 and SIRT2 Loci and Growth in Tibetan Sheep" Animals 10, no. 8: 1362. https://doi.org/10.3390/ani10081362
APA StyleGui, L.-s., Raza, S. H. A., Zhou, L., Garcia, M., Abd El-Aziz, A. H., Wei, D., Hou, S., Jia, J., & Wang, Z. (2020). Association between Single Nucleotide Polymorphisms in SIRT1 and SIRT2 Loci and Growth in Tibetan Sheep. Animals, 10(8), 1362. https://doi.org/10.3390/ani10081362