Evaluation of Poultry Stunning with Low Atmospheric Pressure, Carbon Dioxide or Nitrogen Using a Single Aversion Testing Paradigm

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval and Humane Endpoints
2.2. Animals and Housing
2.3. Training and Test Arena
2.4. Selection of Focal Animals
2.5. Training Sessions
2.6. Final Testing
2.7. Behavioural Scoring
- -
- Gasping was defined as wide open-mouth breathing with neck extension (Videos S2 and S3).
- -
- Headshaking was defined as rapid side-to-side movement of the head, which occurred whilst the animal was standing or walking (Video S4).
- -
- Wing-flapping was defined by rapid movement of the wings in a motion similar to attempted flight (Video S5).
- -
- Jumping was defined as any vertical movement from a plantar stance, resulting in both feet leaving contact with the floor.
- -
- Ataxia was defined as uncoordinated walking with exaggerated lateral movement or as the use of wing when standing to maintain posture (Videos S6 and S7).
- -
- Loss of posture (LOP) was defined by cessation of standing with the head resting against either the floor or wall of the chamber (Video S8).
2.8. Statistical Analyses
3. Results
3.1. Animals and Training Outcomes
3.2. General Behaviour Patterns
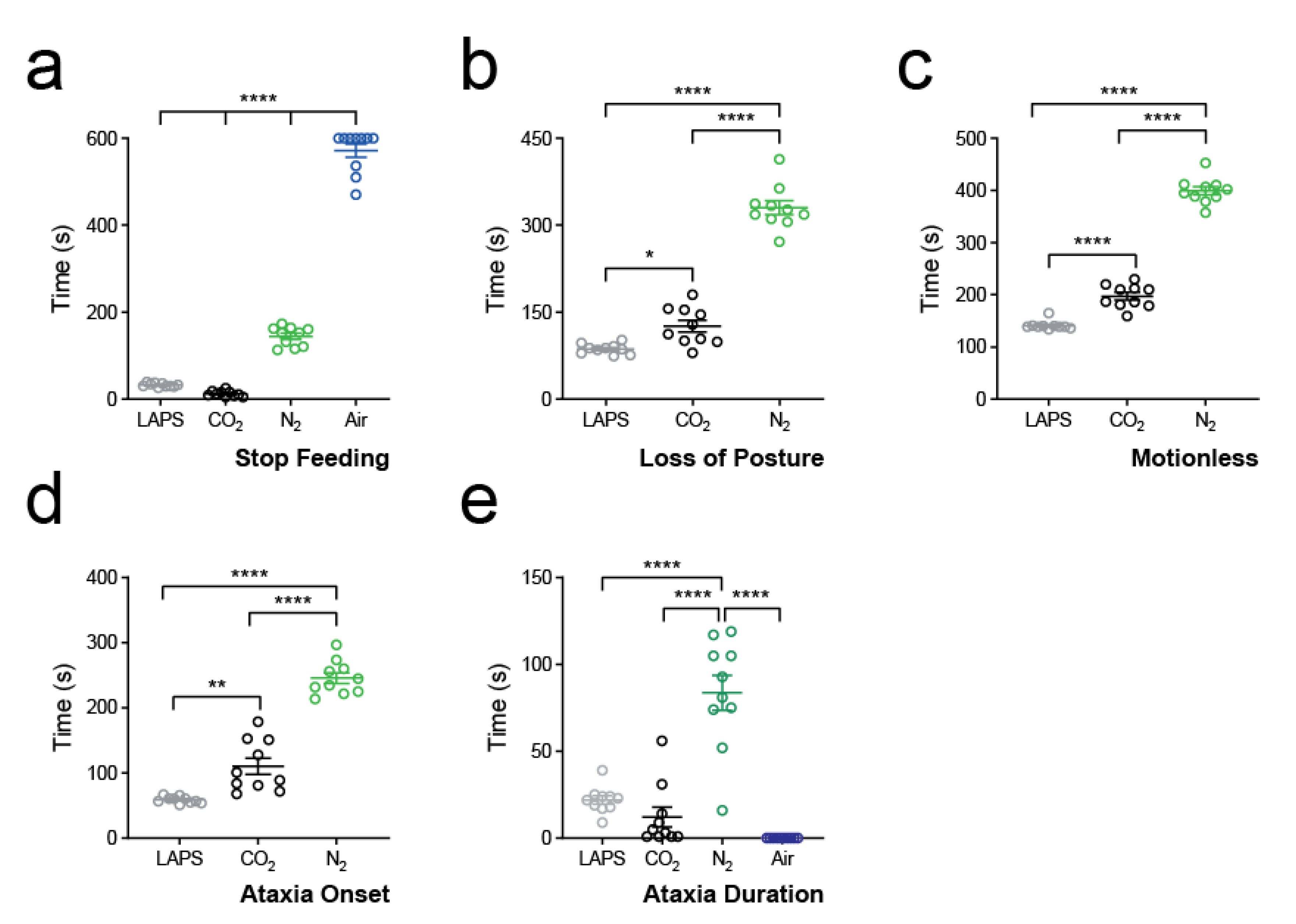
3.3. Primary Aversion Behaviours
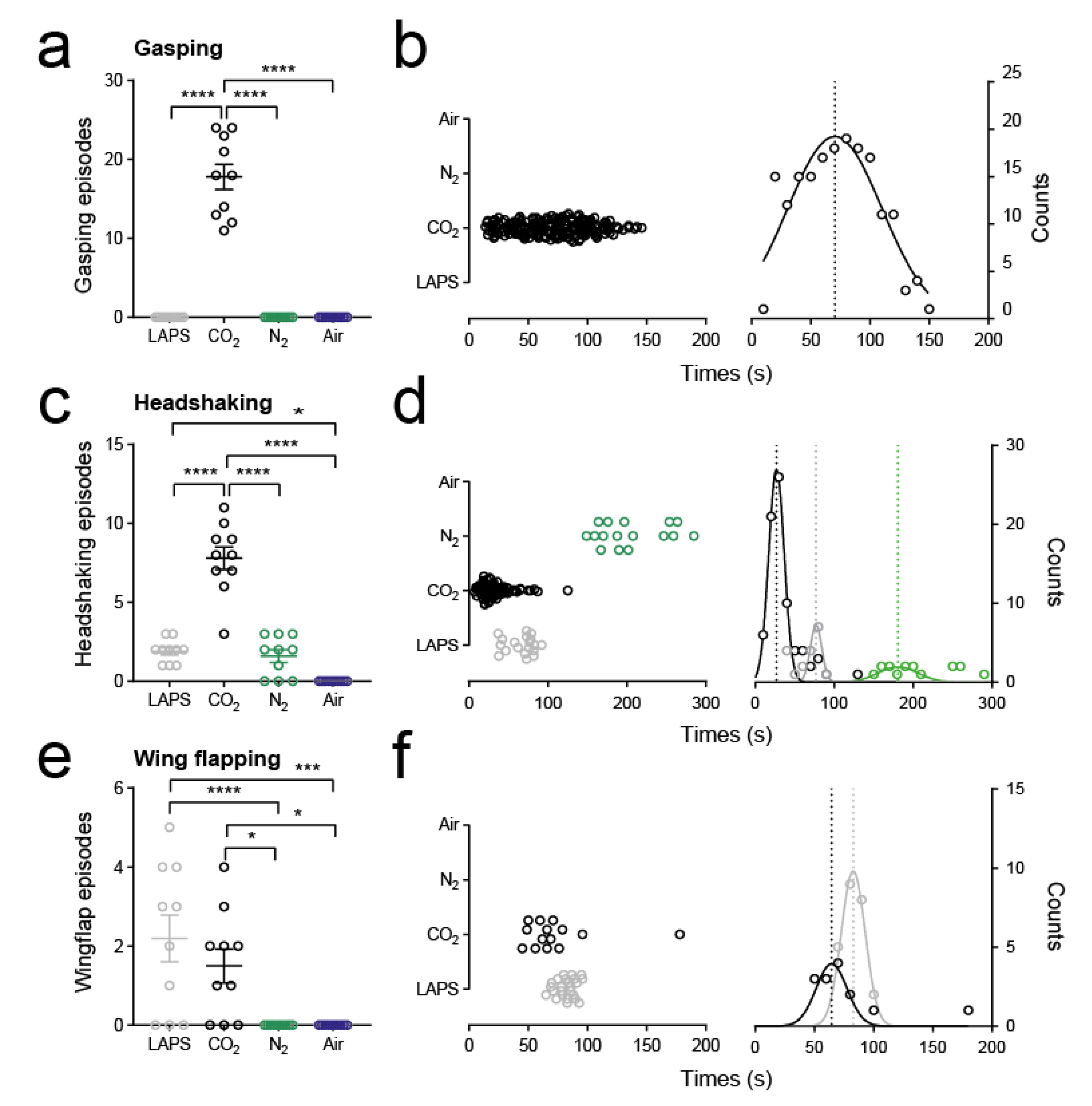
3.4. Secondary Aversion Behaviours
4. Discussion
4.1. Novelty of Methods
4.2. General Behaviour
4.3. Signs of Aversion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steiner, A.R.; Flammer, S.A.; Beausoleil, N.; Berg, L.; Bettschart-Wolfensberger, R.; Pinillos, R.G.; Golledge, H.D.; Marahrens, M.; Meyer, R.E.; Schnitzer, T.; et al. Humanely Ending the Life of Animals: Research Priorities to Identify Alternatives to Carbon Dioxide. Animals 2019, 9, 911. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, P.; Prescott, M.J.; Carbone, L.; Dennison, N.; Johnson, C.B.; Makowska, I.J.; Marquardt, N.; Readman, G.D.; Weary, D.M.; Golledge, H.D. A Good Death? Report of the Second Newcastle Meeting on Laboratory Animal Euthanasia. Animals 2016, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, N.; Feja, M.; Hünigen, H.; Plendl, J.; Menken, L.; Fink, H.; Bert, B. Euthanasia of laboratory mice: Are isoflurane and sevoflurane real alternatives to carbon dioxide? PLoS ONE 2018, 13, e0203793. [Google Scholar] [CrossRef] [PubMed]
- Boivin, G.P.; Bottomley, M.A.; Dudley, E.S.; Schiml, P.A.; Wyatt, C.N.; Grobe, N. Physiological, Behavioral, and Histological Responses of Male C57BL/6N Mice to Different CO2 Chamber Replacement Rates. J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 451–461. [Google Scholar]
- Raj, M.; O’Callaghan, M.; Thompson, K.; Beckett, D.; Morrish, I.; Love, A.; Hickman, G.; Howson, S. Large scale killing of poultry species on farm during outbreaks of diseases: Evaluation and development of a humane containerised gas killing system. World’s Poult. Sci. J. 2008, 64, 227–243. [Google Scholar] [CrossRef]
- Sandilands, V.; Raj, A.; Baker, L.; Sparks, N. Aversion of chickens to various lethal gas mixtures. Anim. Welf. UFAW J. 2011, 20, 253. [Google Scholar]
- Verhoeven, M.; Gerritzen, M.; Velarde, A.; Hellebrekers, L.; Kemp, B. Time to Loss of Consciousness and Its Relation to Behavior in Slaughter Pigs during Stunning with 80 or 95% Carbon Dioxide. Front. Veter Sci. 2016, 3, 38. [Google Scholar] [CrossRef]
- Gentle, M.; Tilston, V. Nociceptors in the legs of poultry: Implications for potential pain in pre-slaughter shackling. Anim. Welf. Potters Bar 2000, 9, 227–236. [Google Scholar]
- Webster, A.B.; Fletcher, D.L. Reactions of Laying Hens and Broilers to Different Gases Used for Stunning Poultry. Poultry Sci. 2001, 80, 1371–1377. [Google Scholar] [CrossRef]
- Berg, C.; Raj, M. A Review of Different Stunning Methods for Poultry-Animal Welfare Aspects (Stunning Methods for Poultry). Animals 2015, 5, 1207–1219. [Google Scholar] [CrossRef]
- Martoft, L.; Lomholt, L.; Kolthoff, C.; Rodriguez, B.E.; Jensen, E.W.; Jorgensen, P.F.; Pedersen, H.D.; Forslid, A. Effects of CO2 anaesthesia on central nervous system activity in swine. Lab. Anim. 2002, 36, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Peppel, P.; Anton, F. Responses of rat medullary dorsal horn neurons following intranasal noxious chemical stimulation: Effects of stimulus intensity, duration, and interstimulus interval. J. Neurophysiol. 1993, 70, 2260–2275. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, D.E.; Demmers, T.G.; Wathes, C.M.; Jones, R.B.; Gentle, M.J. Response characteristics of nasal trigeminal nociceptors in Gallus domesticus. NeuroReport 2002, 13, 1033–1035. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, D.E.F.; Smith, F.S.; Demmers, T.G.; Wathes, C.M.; Jones, R.B. Behavioral correlates of olfactory and trigeminal gaseous stimulation in chickens, Gallus domesticus. Physiol. Behav. 2005, 84, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M.; Wysocki, C.J.; Radil, T. Time-intensity ratings of nasal irritation from carbon dioxide. Chem. Senses 2003, 28, 751–760. [Google Scholar] [CrossRef]
- Danneman, P.J.; Stein, S.; Walshaw, S.O. Humane and practical implications of using carbon dioxide mixed with oxygen for anesthesia or euthanasia of rats. Lab. Anim. Sci. 1997, 47, 376–385. [Google Scholar]
- Gerritzen, M.A.; Lambooij, B.; Reimert, H.; Stegeman, A.; Spruijt, B. On-farm euthanasia of broiler chickens: Effects of different gas mixtures on behavior and brain activity. Poult. Sci. 2004, 83, 1294–1301. [Google Scholar] [CrossRef]
- Raj, A. Recent developments in stunning and slaughter of poultry. World’s Poult. Sci. J. 2006, 62, 467–484. [Google Scholar] [CrossRef]
- Mackie, N.; McKeegan, D.E.; Information, P.E.K.F.C. Behavioural responses of broiler chickens during low atmospheric pressure stunning. Appl. Anim. Behav. Sci. 2016, 174, 90–98. [Google Scholar] [CrossRef]
- Martin, J.E.; Christensen, K.; Vizzier-Thaxton, Y.; McKeegan, D.E.F. Effects of light on responses to low atmospheric pressure stunning in broilers. Br. Poult. Sci. 2016, 57, 1–16. [Google Scholar] [CrossRef]
- Martin, J.; Christensen, K.; Vizzier-Thaxton, Y.; Mitchell, M.A.; McKeegan, D.E. Behavioural, brain and cardiac responses to hypobaric hypoxia in broiler chickens. Physiol. Behav. 2016, 163, 25–36. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Animal Health and Welfare (AHAW); More, S.; Bicout, D.; Bøtner, A.; Butterworth, A.; Calistri, P.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; et al. Low atmospheric pressure system for stunning broiler chickens. EFSA J. 2017, 15, e05056. [Google Scholar] [CrossRef]
- Weary, D.M.; Droege, P.; Braithwaite, V.A. Behavioural Evidence of Felt Emotions: Approaches, Inferences and Refinements. In Advances in the Study of Behavior; Naguib, M., Podos, J., Simmons, L.W., Barrett, L., Healy, S., Zuk, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 49, pp. 27–48. [Google Scholar]
- Poole, G.H.; Fletcher, D.L. A Comparison of Argon, Carbon Dioxide, and Nitrogen in a Broiler Killing System. Poult. Sci. 1995, 74, 1218–1223. [Google Scholar] [CrossRef]
- Zuidhof, M.; Schneider, B.L.; Carney, V.L.; Korver, D.R.; Robinson, F.E. Growth, efficiency, and yield of commercial broilers from 1957, 1978, and 2005. Poult. Sci. 2014, 93, 2970–2982. [Google Scholar] [CrossRef]
- De Jong, I.C.; Van Voorst, A.S.; Blokhuis, H. Parameters for quantification of hunger in broiler breeders. Physiol. Behav. 2003, 78, 773–783. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare. Scientific Opinion on welfare aspects of the management and housing of the grand-parent and parent stocks raised and kept for breeding purposes. EFSA J. 2010, 8, 1667. [Google Scholar] [CrossRef]
- Lambooij, E.; Gerritzen, M.; Engel, B.; Hillebrand, S.; Lankhaar, J.; Pieterse, C. Behavioural responses during exposure of broiler chickens to different gas mixtures. Appl. Anim. Behav. Sci. 1999, 62, 255–265. [Google Scholar] [CrossRef]
- Gerritzen, M.A.; Reimert, H.G.M.; Hindle, V.A.; Verhoeven, M.T.W.; Veerkamp, W.B. Multistage carbon dioxide gas stunning of broilers. Poult. Sci. 2012, 92, 41–50. [Google Scholar] [CrossRef]
- Martin, J.; McKeegan, D.E.F.; Magee, D.L.; Armour, N.; Pritchard, D.G. Pathological consequences of low atmospheric pressure stunning in broiler chickens. Animal 2019, 14, 129–137. [Google Scholar] [CrossRef]
- Kirkden, R.D.; Niel, L.; Lee, G.; Makowska, I.J.; Pfaffinger, M.J.; Weary, D.M. The validity of using an approach-avoidance test to measure the strength of aversion to carbon dioxide in rats. Appl. Anim. Behav. Sci. 2008, 114, 216–234. [Google Scholar] [CrossRef]
- Makowska, I.J.; Vickers, L.; Mancell, J.; Weary, D.M. Evaluating methods of gas euthanasia for laboratory mice. Appl. Anim. Behav. Sci. 2009, 121, 230–235. [Google Scholar] [CrossRef]
- Leach, M.C.; Bowell, V.A.; Allan, T.F.; Morton, D.B. Aversion to gaseous euthanasia agents in rats and mice. Comp. Med. 2002, 52, 249–257. [Google Scholar] [PubMed]
- Webster, A.; Fletcher, D.L. Assessment of the aversion of hens to different gas atmospheres using an approach-avoidance test. Appl. Anim. Behav. Sci. 2004, 88, 275–287. [Google Scholar] [CrossRef]
- Raj, A.B.M. Aversive reactions of turkeys to argon, carbon dioxide and a mixture of carbon dioxide and argon. Veter Rec. 1996, 138, 592–593. [Google Scholar] [CrossRef]
- Raj, A.B.M.; Gregory, N.G. Effect of argon stunning, rapid chilling and early filleting on texture of broiler breast meat. Br. Poult. Sci. 1991, 32, 741–746. [Google Scholar] [CrossRef]
- Coenen, A.; Lankhaar, J.; Lowe, J.; McKeegan, D.E. Animal Consciousness and Euthanasia: Chicken Euthanasia in the Two-Phase Stunning System. In Proceedings of the 17th European Symposium on the Quality of Poultry Meat, Doorwerth, The Netherlands, 23–26 May 2005. [Google Scholar]
- Bouton, M.E. Extinction of instrumental (operant) learning: Interference, varieties of context, and mechanisms of contextual control. Psychopharmacology (Berl.) 2018, 236, 7–19. [Google Scholar] [CrossRef]
- Paul, E.S.; Edgar, J.; Caplen, G.; Nicol, C. Examining affective structure in chickens: Valence, intensity, persistence and generalization measured using a Conditioned Place Preference Test. Appl. Anim. Behav. Sci. 2018, 207, 39–48. [Google Scholar] [CrossRef]
- Wilcoxon, H.C.; Dragoin, W.B.; Kral, P.A. Illness-Induced Aversions in Rat and Quail: Relative Salience of Visual and Gustatory Cues. Science 1971, 171, 826–828. [Google Scholar] [CrossRef]
- Gregory, N.G.; Wotton, S.B. Effect of stunning on spontaneous physical activity and evoked activity in the brain. Br. Poult. Sci. 1990, 31, 215–220. [Google Scholar] [CrossRef]
- Hughes, B. Headshaking in fowls: The effect of environmental stimuli. Appl. Anim. Ethol. 1983, 11, 45–53. [Google Scholar] [CrossRef]
- Gerritzen, M.A.; Lambooij, E.; Hillebrand, S.J.; Lankhaar, J.A.; Pieterse, C. Behavioral responses of broilers to different gaseous atmospheres. Poult. Sci. 2000, 79, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Beausoleil, N.; Mellor, D. Introducing breathlessness as a significant animal welfare issue. N. Z. Veter J. 2014, 63, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Parshall, M.B.; Schwartzstein, R.M.; Adams, L.; Banzett, R.B.; Manning, H.L.; Bourbeau, J.; Calverley, P.M.; Gift, A.G.; Harver, A.; Lareau, S.C.; et al. An Official American Thoracic Society Statement: Update on the Mechanisms, Assessment, and Management of Dyspnea. Am. J. Respir. Crit. Care Med. 2012, 185, 435–452. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Group | Average (Mean) | Variance (s.e.m.) | ANOVA | CO2 | N2 | Air |
|---|---|---|---|---|---|---|---|
| Body mass (kg) | LAPS | 4.25 | 0.10 | p = 0.6324; F = 0.5797 | 0.9172 | 0.7450 | 0.9329 |
| CO2 | 4.38 | 0.12 | 0.9329 | 0.9329 | |||
| N2 | 4.45 | 0.11 | 0.9329 | ||||
| Air | 4.36 | 0.07 | |||||
| Feeding time (s) | LAPS | 32.30 | 1.391 | p < 0.0001; F = 958.1 | <0.0001 | <0.0001 | <0.0001 |
| CO2 | 12.40 | 2.023 | <0.0001 | <0.0001 | |||
| N2 | 144.7 | 7.002 | <0.0001 | ||||
| Air | 571.9 | 15.14 | |||||
| Headshaking (no. episodes) | LAPS | 1.900 | 0.2333 | p < 0.0001; F = 64.88 | <0.0001 | 0.9586 | 0.0159 |
| CO2 | 7.800 | 0.7118 | <0.0001 | <0.0001 | |||
| N2 | 1.600 | 0.4000 | 0.0534 | ||||
| Air | 0.000 | 0.000 | |||||
| Headshaking (time of first episode) | LAPS | 58.40 | 6.344 | p < 0.0001; F = 155 | 0.0001 | <0.0001 | - |
| CO2 | 14.90 | 1.958 | <0.0001 | - | |||
| N2 | 185.6 | 12.46 | - | ||||
| Air | - | - | |||||
| Gasping (no. episodes) | LAPS | 0.000 | 0.000 | p < 0.0001; F = 139.5 | <0.0001 | >0.9999 | >0.9999 |
| CO2 | 18.40 | 1.558 | <0.0001 | <0.0001 | |||
| N2 | 0.000 | 0.000 | >0.9999 | ||||
| Air | 0.000 | 0.000 | |||||
| Jumping (no. episodes) | LAPS | 0.4 | 0.3055 | p = 0.4206; F = 0.9633 | 0.8480 | 0.6190 | 0.3776 |
| CO2 | 0.2 | 0.1333 | 0.9770 | 0.8480 | |||
| N2 | 0.1 | 0.1000 | 0.9770 | ||||
| Air | 0.0 | 0.000 | |||||
| Wing flaps (no. episodes pre-LOP) | LAPS | 2.1 | 0.5667 | p = 0.0001; F = 9.04 | 0.6341 | 0.001 | 0.001 |
| CO2 | 1.5 | 0.4282 | 0.0249 | 0.0249 | |||
| N2 | 0.0 | 0.000 | >0.9999 | ||||
| Air | 0.0 | 0.000 | |||||
| Wing flap duration (post-LOP(s)) | LAPS | 16.50 | 1.344 | p < 0.0001; F = 14.74 | 0.0016 | 0.3931 | - |
| CO2 | 7.100 | 1.362 | <0.0001 | - | |||
| N2 | 19.70 | 2.251 | - | ||||
| Air | - | - | |||||
| Ataxia onset (s) | LAPS | 59.00 | 1.626 | p < 0.0001; F = 125.1 | 0.0074 | <0.0001 | - |
| CO2 | 110.6 | 12.41 | <0.0001 | - | |||
| N2 | 246.0 | 8.192 | - | ||||
| Air | - | - | |||||
| Ataxia duration (s) | LAPS | 22.00 | 2.418 | p < 0.0001; F = 39.77 | 0.6474 | <0.0001 | 0.0573 |
| CO2 | 12.20 | 5.700 | <0.0001 | 0.4714 | |||
| N2 | 83.70 | 10.06 | <0.0001 | ||||
| Air | 0.0 | 0.000 | |||||
| Time to LOM (s) | LAPS | 141.2 | 2.724 | p < 0.0001; F = 464.8 | <0.0001 | <0.0001 | - |
| CO2 | 197.4 | 7.011 | <0.0001 | - | |||
| N2 | 399.4 | 7.901 | - | ||||
| Air | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gent, T.C.; Gebhardt-Henrich, S.; Schild, S.-L.A.; Rahman, A.A.; Toscano, M.J. Evaluation of Poultry Stunning with Low Atmospheric Pressure, Carbon Dioxide or Nitrogen Using a Single Aversion Testing Paradigm. Animals 2020, 10, 1308. https://doi.org/10.3390/ani10081308
Gent TC, Gebhardt-Henrich S, Schild S-LA, Rahman AA, Toscano MJ. Evaluation of Poultry Stunning with Low Atmospheric Pressure, Carbon Dioxide or Nitrogen Using a Single Aversion Testing Paradigm. Animals. 2020; 10(8):1308. https://doi.org/10.3390/ani10081308
Chicago/Turabian StyleGent, Thomas C., Sabine Gebhardt-Henrich, Sarah-Lina Aagaard Schild, Abdulsatar Abdel Rahman, and Michael J. Toscano. 2020. "Evaluation of Poultry Stunning with Low Atmospheric Pressure, Carbon Dioxide or Nitrogen Using a Single Aversion Testing Paradigm" Animals 10, no. 8: 1308. https://doi.org/10.3390/ani10081308
APA StyleGent, T. C., Gebhardt-Henrich, S., Schild, S.-L. A., Rahman, A. A., & Toscano, M. J. (2020). Evaluation of Poultry Stunning with Low Atmospheric Pressure, Carbon Dioxide or Nitrogen Using a Single Aversion Testing Paradigm. Animals, 10(8), 1308. https://doi.org/10.3390/ani10081308