Odour-Evoked Memory in Dogs: Do Odours Help to Retrieve Memories of Food Location?


{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Apparatus
2.3. Procedure
2.3.1. Phase 1: Memory Encoding
2.3.2. Phase 2: Memory Retrieval
- Group 1: control odour (i.e., apple)—vanilla odour;
- Group 2: vanilla odour—control odour;
- Group 3: no odour—no odour.
2.4. Data Analysis
- 2: dogs touched with their muzzle or paw a correct item;
- 1: dogs made an olfactory check to explore a correct item;
- 0: dogs explore again the same item (for both the correct and incorrect ones);
- −1: dogs made an olfactory check to explore an incorrect item;
- −2: dogs touched with their muzzle or paw an incorrect item.
2.5. Ethics Statement
3. Results
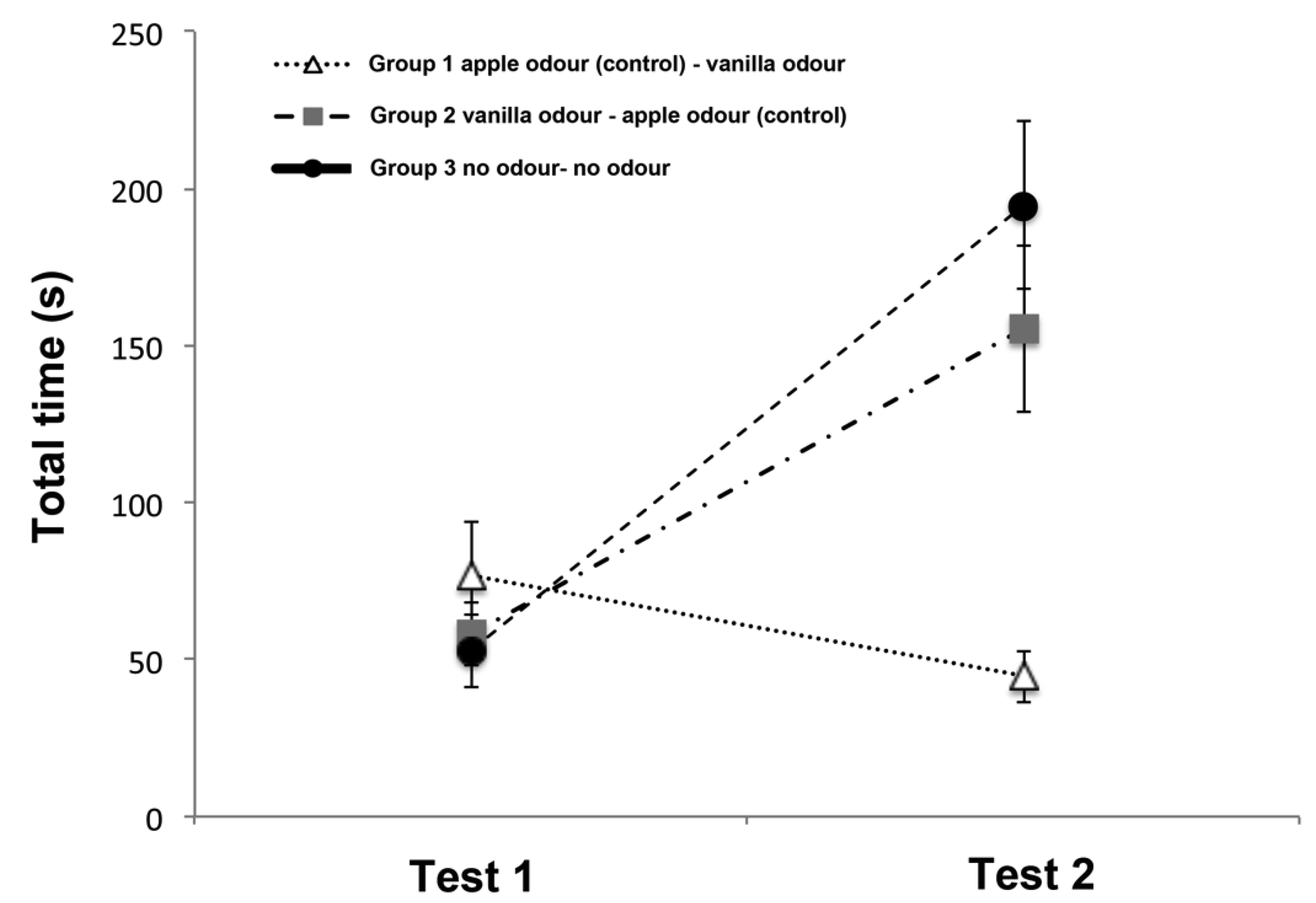
3.1. Total Time
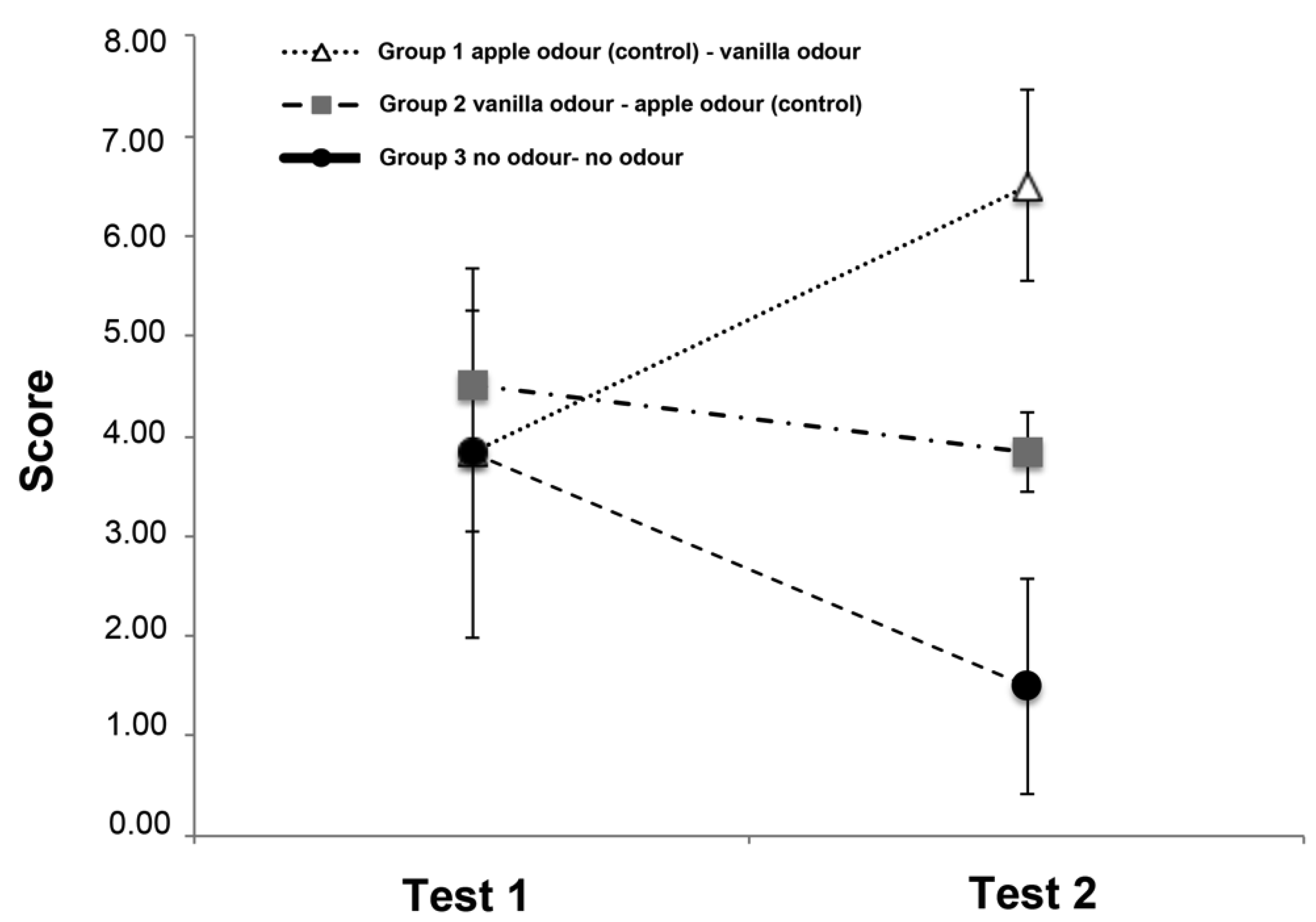
3.2. Score
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Walker, D.B.; Walker, J.C.; Cavnar, P.J.; Taylor, J.L.; Pickel, D.H.; Hall, S.B.; Suarez, J.C. Naturalistic quantification of canine olfactory sensitivity. Appl. Anim. Behav. Sci. 2006, 97, 241–254. [Google Scholar] [CrossRef]
- Papet, L.E. Narcotic and explosive odors: Volatile organic compounds as training aids for olfactory detection. In Canine Olfaction Science and Law; Jezierski, T., Ensminger, J., Papet, L.E., Eds.; CRC Press: Cleveland, OH, USA, 2016; pp. 265–278. ISBN 978-1482260236. [Google Scholar]
- Bijland, L.R.; Bomers, M.K.; Smulders, Y.M. Smelling the diagnosis a review on the use of scent in diagnosing. Neth. J. Med. 2013, 71, 300–307. [Google Scholar] [PubMed]
- Catala, A.; Grandgeorge, M.; Schaff, J.L.; Cousillas, H.; Hausberger, M.; Cattet, J. Dogs demonstrate the existence of an epileptic seizure odour in humans. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bekoff, M. Observations of scent-marking and discriminating self from other by a domestic dog (Canis familiaris): Tales of displaced yellow snow. Behav. Process. 2001, 55, 75–79. [Google Scholar] [CrossRef]
- Siniscalchi, M.; D’Ingeo, S.; Minunno, M.; Quaranta, A. Communication in dogs. Animals 2018, 8, 131. [Google Scholar] [CrossRef]
- Siniscalchi, M.; D’Ingeo, S.; Quaranta, A. The dog nose “KNOWS” fear: Asymmetric nostril use during sniffing at canine and human emotional stimuli. Behav. Brain Res. 2016, 304, 34–41. [Google Scholar] [CrossRef]
- D’Aniello, B.; Semin, G.R.; Alterisio, A.; Aria, M.; Scandurra, A. Interspecies transmission of emotional information via chemosignals: From humans to dogs (Canis lupus familiaris). Anim. Cognit. 2018, 21, 67–78. [Google Scholar] [CrossRef]
- Graham, L.; Wells, D.L.; Hepper, P.G. The influence of olfactory stimulation on the behaviour of dogs housed in a rescue shelter. Appl. Anim. Behav. Sci. 2005, 91, 143–153. [Google Scholar] [CrossRef]
- Murtagh, K.; Farnworth, M.J.; Brilot, B.O. The scent of enrichment: Exploring the effect of odour and biological salience on behaviour during enrichment of kennelled dogs. Appl. Anim. Behav. Sci. 2020, 223, 104917. [Google Scholar] [CrossRef]
- Duranton, C.; Horowitz, A. Let me sniff! Nosework induces positive judgment bias in pet dogs. Appl. Anim. Behav. Sci. 2019, 211, 61–66. [Google Scholar] [CrossRef]
- Gazit, I.; Goldblatt, A.; Terkel, J. Formation of an olfactory search image for explosives odours in sniffer dogs. Ethology 2005, 111, 669–680. [Google Scholar] [CrossRef]
- Bräuer, J.; Belger, J. A ball is not a Kong: Odor representation and search behavior in domestic dogs (Canis familiaris) of different education. J. Comp. Psychol. 2018, 132, 189. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.W.; Hendriks, M.; Aarts, H. Smells like clean spirit: Nonconscious effects of scent on cognition and behavior. Psychol. Sci. 2005, 16, 689–693. [Google Scholar] [CrossRef] [PubMed]
- De Lange, M.A.; Debets, L.W.; Ruitenburg, K.; Holland, R.W. Making less of a mess: Scent exposure as a tool for behavioral change. Soc. Influ. 2012, 7, 90–97. [Google Scholar] [CrossRef]
- Schnall, S.; Haidt, J.; Clore, G.L.; Jordan, A.H. Disgust as embodied moral judgment. Pers. Soc. Psychol. Bull. 2008, 34, 1096–1109. [Google Scholar] [CrossRef]
- Liljenquist, K.; Zhong, C.B.; Galinsky, A.D. The smell of virtue: Clean scents promote reciprocity and charity. Psychol. Sci. 2010, 21, 381–383. [Google Scholar] [CrossRef]
- Fenko, A.; Loock, C. The influence of ambient scent and music on patients’ anxiety in a waiting room of a plastic surgeon. HERD 2014, 7, 38–59. [Google Scholar] [CrossRef] [PubMed]
- Herz, R.S.; Schankler, C.; Beland, S. Olfaction, Emotion and Associative Learning: Effects on Motivated Behavior. Motiv. Emot. 2004, 28, 363–383. [Google Scholar] [CrossRef]
- Proust, M. À la Récherche du Temps Perdu; Grasset, B., Ed.; Editions de la Nouvelle Revue Francaise: Paris, France, 1913. [Google Scholar]
- Herz, R.S. Odor-Evoked Memory. In The Oxford Handbook of Social Neuroscience; Decety, J., Cacioppo, J.T., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 265–276. [Google Scholar] [CrossRef]
- Tulving, E.; Thomson, D.M. Encoding specificity and retrieval processes in episodic memory. Psychol. Rev. 1973, 80, 352. [Google Scholar] [CrossRef]
- Herz, R.S.; Engen, T. Odor memory: Review and analysis. Psychon. Bull. Rev. 1996, 3, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Herz, R.S. Emotion experienced during encoding enhances odor retrieval cue effectiveness. Am. J. Psychol. 1997, 110, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Herz, R.S. A naturalistic analysis of autobiographical memories triggered by olfactory visual and auditory stimuli. Chem. Senses 2004, 29, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Herz, R.S.; Schooler, J.W. A naturalistic study of autobiographical memories evoked by olfactory and visual cues: Testing the Proustian hypothesis. Am. J. Psychol. 2002, 115, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Willander, J.; Larsson, M. Olfaction and emotion: The case of autobiographical memory. Mem. Cognit. 2007, 35, 1659–1663. [Google Scholar] [CrossRef]
- Bonfigli, L.; Kodilja, R.; Zanuttini, L. Verbal versus olfactory cues: Affect in elicited memories. Percept. Mot. Skills 2002, 94, 9–20. [Google Scholar] [CrossRef]
- Herz, R.S.; Eliassen, J.; Beland, S.; Souza, T. Neuroimaging evidence for the emotional potency of odor-evoked memory. Neuropsychologia 2003, 42, 371–378. [Google Scholar] [CrossRef]
- Arshamian, A.; Iannilli, E.; Gerber, J.C.; Willander, J.; Persson, J.; Seo, H.S.; Hummel, T.; Larsson, M. The functional neuroanatomy of odor evoked autobiographical memories cued by odors and words. Neuropsychologia 2013, 51, 123–131. [Google Scholar] [CrossRef]
- Herz, R.S. Are Odors the Best Cues to Memory? A Cross-Modal Comparison of Associative Memory Stimuli. Ann. N. Y. Acad. Sci. 1998, 855, 670–674. [Google Scholar] [CrossRef]
- Toffolo, M.B.; Smeets, M.A.; Van Den Hout, M.A. Proust revisited: Odours as triggers of aversive memories. Cognit. Emot. 2012, 26, 83–92. [Google Scholar] [CrossRef]
- Herz, R.S.; Cupchik, G.C. An experimental characterization of odor-evoked memories in humans. Chem. Senses 1992, 17, 519–528. [Google Scholar] [CrossRef]
- Chu, S.; Downes, J.J. Proust nose best: Odors are better cues of autobiographical memory. Mem. Cognit. 2002, 30, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Maylor, E.A.; Carter, S.M.; Hallett, E.L. Preserved olfactory cuing of autobiographical memories in old age. J. Gerontol. B Psychol. 2002, 57, P41–P46. [Google Scholar] [CrossRef] [PubMed]
- Aggleton, J.P.; Waskett, L. The ability of odours to serve as state-dependent cues for real-world memories: Can Viking smells aid the recall of Viking experiences? Br. J. Psychol. 1999, 90, 1–7. [Google Scholar] [CrossRef] [PubMed]
- De Bruijn, M.J.; Bender, M. Olfactory cues are more effective than visual cues in experimentally triggering autobiographical memories. Memory 2018, 26, 547–558. [Google Scholar] [CrossRef]
- Saive, A.L.; Ravel, N.; Thévenet, M.; Royet, J.P.; Plailly, J. A novel experimental approach to episodic memory in humans based on the privileged access of odors to memories. J. Neurosci. Methods 2013, 213, 22–31. [Google Scholar] [CrossRef]
- Gagnon, S.; Doré, F.Y. Search behavior in various breeds of adult dogs (Canis familiaris): Object permanence and olfactory cues. J. Comp. Psychol. 1992, 106, 58. [Google Scholar] [CrossRef]
- Gagnon, S.; Doré, F.Y. Search behavior of dogs (Canis familiaris) in invisible displacement problems. Anim. Learn. Behav. 1993, 21, 246–254. [Google Scholar] [CrossRef]
- Fiset, S.; Beaulieu, C.; Landry, F. Duration of dogs’ (Canis familiaris) working memory in search for disappearing objects. Anim. Cognit. 2003, 6, 1–10. [Google Scholar] [CrossRef]
- Zentall, T.R.; Pattison, K.F. Now you see it, now you don’t: Object permanence in dogs. Curr. Dir. Psychol. Sci. 2016, 25, 357–362. [Google Scholar] [CrossRef]
- Milgram, N.W.; Adams, B.; Callahan, H.; Head, E.; Mackay, B.; Thirlwell, C.; Cotman, C.W. Landmark discrimination learning in the dog. Learn. Mem. 1999, 6, 54–61. [Google Scholar] [CrossRef]
- Milgram, N.W.; Head, E.; Muggenburg, B.; Holowachuk, D.; Murphey, H.; Estrada, J.; Ikeda-Douglas, C.J.; Zicker, S.C.; Cotman, C.W. Landmark discrimination learning in the dog: Effects of age, an antioxidant fortified food, and cognitive strategy. Neurosci. Biobehav. Rev. 2002, 26, 679–695. [Google Scholar] [CrossRef]
- Macpherson, K.; Roberts, W.A. Spatial memory in dogs (Canis familiaris) on a radial maze. J. Comp. Psychol. 2010, 124, 47. [Google Scholar] [CrossRef]
- Salvin, H.E.; McGreevy, P.D.; Sachdev, P.S.; Valenzuela, M.J. The canine sand maze: An appetitive spatial memory paradigm sensitive to age-related change in dogs. J. Exp. Anal. Behav. 2011, 95, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Herz, R.S. The effects of cue distinctiveness on odor-based context-dependent memory. Mem. Cognit. 1997, 25, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Bentosela, M.; Jakovcevic, A.; Elgier, A.M.; Mustaca, A.E.; Papini, M.R. Incentive contrast in domestic dogs (Canis familiaris). J. Comp. Psychol. 2009, 123, 125. [Google Scholar] [CrossRef]
- Strauch, C.; Manahan-Vaughan, D. Orchestration of Hippocampal Information Encoding by the Piriform Cortex. Cereb. Cortex 2020, 30, 135–147. [Google Scholar] [CrossRef]
- Aqrabawi, A.J.; Kim, J.C. Hippocampal projections to the anterior olfactory nucleus differentially convey spatiotemporal information during episodic odour memory. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Eichenbaum, H. Memory: Organization and control. Annu. Rev. Psychol. 2017, 68, 19–45. [Google Scholar] [CrossRef]
- Kerr, K.M.; Agster, K.L.; Furtak, S.C.; Burwell, R.D. Functional neuroanatomy of the parahippocampal region: The lateral and medial entorhinal areas. Hippocampus 2007, 17, 697–708. [Google Scholar] [CrossRef]
- Agster, K.L.; Burwell, R.D. Cortical efferents of the perirhinal, postrhinal, and entorhinal cortices of the rat. Hippocampus 2009, 19, 1159–1186. [Google Scholar] [CrossRef]
- Truchet, B.; Chaillan, F.A.; Soumireu-Mourat, B.; Roman, F.S. Learning and memory of cue-reward association meaning by modifications of synaptic efficacy in dentate gyrus and piriform cortex. Hippocampus 2002, 12, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Gallistel, C.R. The Organization of Learning; The MIT Press: Cambridge, MA, USA, 1990; ISBN 978-0262570985. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quaranta, A.; d’Ingeo, S.; Siniscalchi, M. Odour-Evoked Memory in Dogs: Do Odours Help to Retrieve Memories of Food Location? Animals 2020, 10, 1249. https://doi.org/10.3390/ani10081249
Quaranta A, d’Ingeo S, Siniscalchi M. Odour-Evoked Memory in Dogs: Do Odours Help to Retrieve Memories of Food Location? Animals. 2020; 10(8):1249. https://doi.org/10.3390/ani10081249
Chicago/Turabian StyleQuaranta, Angelo, Serenella d’Ingeo, and Marcello Siniscalchi. 2020. "Odour-Evoked Memory in Dogs: Do Odours Help to Retrieve Memories of Food Location?" Animals 10, no. 8: 1249. https://doi.org/10.3390/ani10081249
APA StyleQuaranta, A., d’Ingeo, S., & Siniscalchi, M. (2020). Odour-Evoked Memory in Dogs: Do Odours Help to Retrieve Memories of Food Location? Animals, 10(8), 1249. https://doi.org/10.3390/ani10081249