Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals



Simple Summary
Abstract

1. Introduction
2. Toxoplasma gondii Antigenic Structure—Antigens and Antigen Complexes
2.1. Surface Antigens (SAG)
2.2. Microneme Antigens (MIC)
2.3. Rhoptry Neck (RON) and Rhoptry Antigens (ROP)
2.4. Dense Granules Antigens (GRA)
3. Animal Toxoplasmosis—Prevalence, Symptoms, and Diagnosis
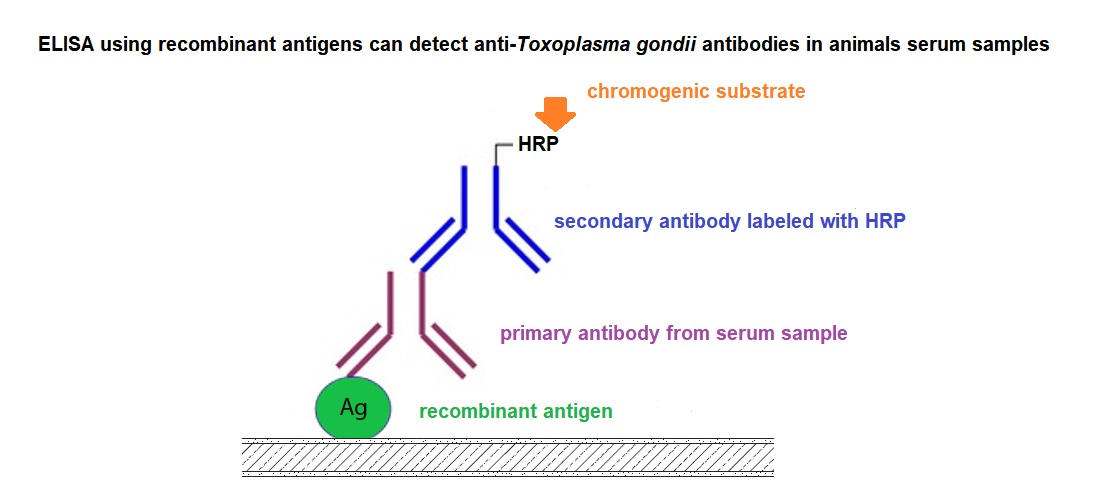
4. Recombinant Antigens in the Diagnosis of Toxoplasmosis
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Buxton, D. Protozoan infections (Toxoplasma gondii, Neospora caninum and Sarcocystis spp.) in sheep and goats: Recent advances. Vet. Res. 1998, 29, 289–310. [Google Scholar]
- Olivier, A.; Herbert, B.; Sava, B.; Pierre, C.; Collins, J.D.; Aline, D.K.; John, G.; Arie, H.; James, H.; Gunter, K.; et al. Surveillance and monitoring of Toxoplasma in humans, food and animals—Scientific Opinion of the Panel on Biological Hazards. EFSA J. 2007, 583, 1–64. [Google Scholar]
- Tassi, P. Toxoplasma gondii infection in horses. A review. Parassitologia 2007, 49, 7–15. [Google Scholar] [PubMed]
- Jones, J.L.; Dubey, J.P. Foodborne Toxoplasmosis. Clin. Infect. Dis. 2012, 55, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.E.; Dubey, J.P. Toxoplasma gondii prevalence in farm animals in the United States. Int. J. Parasitol. 2013, 43, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Sam-Yellowe, T.Y. Rhoptry organelles of the apicomplexa: Their role in host cell invasion and intracellular survival. Parasitol. Today 1996, 12, 308–316. [Google Scholar] [CrossRef]
- Carruthers, V.B.; Sibley, L.D. Sequential protein secretion front three distinct organelles of Toxoplasma gondii accompanies invasion of human fibroblasts. Eur. J. Cell Biol. 1997, 73, 114–123. [Google Scholar]
- Carruthers, V.B.; Giddings, O.K.; Sibley, L.D. Secretion of micronemal proteins is associated with Toxoplasma invasion of host cells. Cell. Microbiol. 1999, 1, 225–235. [Google Scholar] [CrossRef]
- Lekutis, C.; Ferguson, D.J.P.; Grigg, M.E.; Camps, M.; Boothroyd, J.C. Surface antigens of Toxoplasma gondii: Variations on a theme. Int. J. Parasitol. 2001, 31, 1285–1292. [Google Scholar] [CrossRef]
- Joiner, K.A.; Roos, D.S. Secretory traffic in the eukaryotic parasite Toxoplasma gondii: Less is more. J. Cell Biol. 2002, 157, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Mercier, C.; Dubremetz, J.F.; Rauscher, B.; Lecordier, L.; Sibley, L.D.; Cesbron-Delauw, M.F. Biogenesis of nanotubular network in Toxoplasma parasitophorous vacuole induced by parasite proteins. Mol. Biol. Cell 2002, 13, 2397–2409. [Google Scholar] [CrossRef] [PubMed]
- Mercier, C.; Adjogble, K.D.Z.; Däubener, W.; Delauw, M.F.C. Dense granules: Are they key organelles to help understand the parasitophorous vacuole of all apicomplexa parasites? Int. J. Parasitol. 2005, 35, 829–849. [Google Scholar] [CrossRef]
- Nagel, S.D.; Boothroyd, J.C. The major surface antigen, P30, of Toxoplasma gondii is anchored by a glycolipid. J. Biol. Chem. 1989, 264, 5569–5574. [Google Scholar] [PubMed]
- Bülow, R.; Boothroyd, J.C. Protection of mice from fatal Toxoplasma gondii infection by immunization with p30 antigen in liposomes. J. Immunol. 1991, 147, 3496–3500. [Google Scholar] [PubMed]
- Manger, I.D.; Hehl, A.B.; Boothroyd, J.C. The surface of Toxoplasma tachyzoites is dominated by a family of glycosylphosphatidylinositol-anchored antigens related to SAG1. Infect. Immun. 1998, 66, 2237–2244. [Google Scholar] [CrossRef] [PubMed]
- Lekutis, C.; Ferguson, D.J.P.; Boothroyd, J.C. Toxoplasma gondii: Identification of a developmentally regulated family of genes related to SAG2. Exp. Parasitol. 2000, 96, 89–96. [Google Scholar] [CrossRef]
- Jung, C.; Lee, C.Y.F.; Grigg, M.E. The SRS superfamily of Toxoplasma surface proteins. Int. J. Parasitol. 2004, 34, 285–296. [Google Scholar] [CrossRef]
- Cesbron-Delauw, M.F.; Tomavo, S.; Beauchamps, P.; Fourmaux, M.P.; Camus, D.; Capron, A.; Dubremetz, J.F. Similarities between the primary structures of two distinct major surface proteins of Toxoplasma gondii. J. Biol. Chem. 1994, 269, 16217–16222. [Google Scholar]
- Tomley, F.M.; Soldati, D.S. Mix and match modules: Structure and function of microneme proteins in apicomplexan parasites. Trends Parasitol. 2001, 17, 81–88. [Google Scholar] [CrossRef]
- Kappe, S.; Bruderer, T.; Gantt, S.; Fujioka, H.; Nussenzweig, V.; Ménard, R. Conservation of a gliding motility and cell invasion machinery in Apicomplexan parasites. J. Cell Biol. 1999, 147, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, V.B. Host cell invasion by the opportunistic pathogen Toxoplasma gondii. Acta Trop. 2002, 81, 111–122. [Google Scholar] [CrossRef]
- Huynh, M.; Rabenau, K.E.; Harper, J.M.; Beatty, W.L.; Sibley, L.D.; Carruthers, V.B. Rapid invasion of host cells by Toxoplasma requires secretion of the MIC2-M2AP adhesive protein complex. EMBO J. 2003, 22, 2082–2090. [Google Scholar] [CrossRef]
- Jewett, T.J.; Sibley, L.D. The Toxoplasma proteins MIC2 and M2AP form a hexameric complex necessary for intracellular survival. J. Biol. Chem. 2004, 279, 9362–9369. [Google Scholar] [CrossRef] [PubMed]
- Cérède, O.; Dubremetz, J.F.; Soête, M.; Deslée, D.; Vial, H.; Bout, D.; Lebrun, M. Synergistic role of micronemal proteins in Toxoplasma gondii virulence. J. Exp. Med. 2005, 201, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Mital, J.; Meissner, M.; Soldati, D.; Ward, G.E. Conditional expression of Toxoplasma gondii apical membrane antigen-1 (TgAMA1) demonstrates that TgAMA1 plays a critical role in host cell invasion. Mol. Biol. Cell 2005, 16, 4341–4349. [Google Scholar] [CrossRef] [PubMed]
- Sawmynaden, K.; Saouros, S.; Friedrich, N.; Marchant, J.; Simpson, P.; Bleijlevens, B.; Blackman, M.J.; Soldati-Favre, D.; Matthews, S. Structural insights into microneme protein assembly reveal a new mode of EGF domain recognition. EMBO Rep. 2008, 9, 1149–1155. [Google Scholar] [CrossRef]
- Carruthers, V.; Boothroyd, J.C. Pulling together: An integrated model of Toxoplasma cell invasion. Curr. Opin. Microbiol. 2007, 10, 83–89. [Google Scholar] [CrossRef]
- Camejo, A.; Gold, D.A.; Lu, D.; McFetridge, K.; Julien, L.; Yang, N.; Jensen, K.D.C.; Saeij, J.P.J. Identification of three novel Toxoplasma gondii rhoptry proteins. Int. J. Parasitol. 2014, 44, 147–160. [Google Scholar] [CrossRef]
- Besteiro, S.; Michelin, A.; Poncet, J.; Dubremetz, J.-F.; Lebrun, M. Export of a Toxoplasma gondii rhoptry neck protein complex at the host cell membrane to form the moving junction during invasion. PLoS Pathog. 2009, 5, e1000309. [Google Scholar] [CrossRef]
- Schwarz, J.A.; Fouts, A.E.; Cummings, C.A.; Ferguson, D.J.P.; Boothroyd, J.C. A novel rhoptry protein in Toxoplasma gondii bradyzoites and merozoites. Mol. Biochem. Parasitol. 2005, 144, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.L.; Yap, A.; Gilson, P.R.; Cowman, A.F.; Crabb, B.S. Insights and controversies into the role of the key apicomplexan invasion ligand, Apical Membrane Antigen 1. Int. J. Parasitol. 2014, 44, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Vetrivel, U.; Muralikumar, S.; Mahalakshmi, B.; Lily Therese, K.; Madhavan, H.; Alameen, M.; Thirumudi, I. Multilevel precision-based rational design of chemical inhibitors targeting the hydrophobic cleft of Toxoplasma gondii apical membrane antigen 1 (AMA1). Genom. Inform. 2016, 14, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Cortez, E.; Stumbo, A.C.; Saldanha-Gama, R.; Villela, C.G.; Barja-Fidalgo, C.; Rodrigues, C.A.; Das Graças Henriques, M.; Benchimol, M.; Barbosa, H.S.; Porto, L.C.; et al. Immunolocalization of an osteopontin-like protein in dense granules of Toxoplasma gondii tachyzoites and its association with the parasitophorous vacuole. Micron 2008, 39, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Leriche, M.A.; Dubremetz, J.F. Exocytosis of Toxoplasma gondii dense granules into the parasitophorous vacuole after host cell invasion. Parasitol. Res. 1990, 76, 559–562. [Google Scholar] [CrossRef]
- Leriche, M.A.; Dubremetz, J.F. Characterization of the protein contents of rhoptries and dense granules of Toxoplasma gondii tachyzoites by subcellular fractionation and monoclonal antibodies. Mol. Biochem. Parasitol. 1991, 45, 249–259. [Google Scholar] [CrossRef]
- Dubremetz, J.F.; Achbarou, A.; Bermudes, D.; Joiner, K.A. Kinetics and pattern of organelle exocytosis during Toxoplasma gondii/host-cell interaction. Parasitol. Res. 1993, 79, 402–408. [Google Scholar] [CrossRef]
- Dubey, J.P.; Jones, J.L. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol. 2008, 38, 1257–1278. [Google Scholar] [CrossRef]
- Sroka, J.; Zwoliński, J.; Dutkiewicz, J. Seroprevalence of Toxoplasma gondii In farm and wild animals from the area of lublin province. Bull. Vet. Inst. Pulawy 2007, 51, 535–540. [Google Scholar]
- Davis, S.W.; Dubey, J.P. Mediation of immunity to Toxoplasma gondii oocyst shedding in cats. J. Parasitol. 1995, 81, 882–886. [Google Scholar] [CrossRef]
- Lappin, M.R.; Dawe, D.L.; Lindl, P.; Greene, C.E.; Prestwood, A.K. Mitogen and antigen-specific induction of lymphoblast transformation in cats with subclinical toxoplasmosis. Vet. Immunol. Immunopathol. 1992, 30, 207–220. [Google Scholar] [CrossRef]
- Dubey, J.P. Duration of Immunity to shedding of Toxoplasma gondii oocysts by cats. J. Parasitol. 1995, 81, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Lappin, M.R.; Greene, C.E.; Prestwood, A.K.; Dawe, D.L.; Tarleton, R.L. Diagnosis of recent Toxoplasma gondii infection in cats by use of an enzyme-linked immunosorbent assay for immunoglobulin M. Am. J. Vet. Res. 1989, 50, 1580–1585. [Google Scholar] [PubMed]
- Freyre, A.; Choromanski, L.; Fishback, J.L.; Popiel, I. Immunization of cats with tissue cysts, bradyzoites, and tachyzoites of the T-263 strain of Toxoplasma gondii. J. Parasitol. 1993, 79, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.K.; Pfefferkorn, E.R.; Smith, D.D.; Fishback, J.L. Prospective vaccine prepared from a new mutant of Toxoplasma gondii for use in cats. Am. J. Vet. Res. 1991, 52, 759–763. [Google Scholar] [PubMed]
- Mateus-Pinilla, N.E.; Dubey, J.P.; Choromanski, L.; Weigel, R.M. A field trial of the effectiveness of a feline Toxoplasma gondii vaccine in reducing T. gondii exposure for swine. J. Parasitol. 1999, 85, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Choromanski, L.; Freyre, A.; Brown, K.; Popiel, I.; Shibley, G. Safety aspects of a vaccine for cats containing a Toxoplasma gondii mutant strain. J. Eukaryot. Microbiol. 1994, 41, 8S. [Google Scholar]
- Choromanski, L.; Freyre, A.; Popiel, R.; Brown, K.; Grieve, R.; Shibley, G. Safety and efficacy of modified live feline Toxoplasma gondii vaccine. Dev. Biol. Stand. 1995, 84, 269–281. [Google Scholar]
- Dubey, J.P.; Carpenter, J.L. Histologically confirmed clinical toxoplasmosis in cats: 100 cases (1952–1990). J. Am. Vet. Med. Assoc. 1993, 203, 1556–1566. [Google Scholar]
- Dubey, J.P. Toxoplasmosis in sheep—The last 20 years. Vet. Parasitol. 2009, 163, 1–14. [Google Scholar] [CrossRef]
- Pereira-Bueno, J.; Quintanilla-Gozalo, A.; Pérez-Pérez, V.; Álvarez-García, G.; Collantes-Fernández, E.; Ortega-Mora, L.M. Evaluation of ovine abortion associated with Toxoplasma gondii in Spain by different diagnostic techniques. Vet. Parasitol. 2004, 121, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasmosis in pigs—The last 20 years. Vet. Parasitol. 2009, 164, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kijlstra, A.; Eissen, O.A.; Cornelissen, J.; Munniksma, K.; Eijck, I.; Kortbeek, T. Toxoplasma gondii infection in animal-friendly pig production systems. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3165–3169. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Hill, D.E.; Rozeboom, D.W.; Rajendran, C.; Choudhary, S.; Ferreira, L.R.; Kwok, O.C.H.; Su, C. High prevalence and genotypes of Toxoplasma gondii isolated from organic pigs in northern USA. Vet. Parasitol. 2012, 188, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Sadrebazzaz, A.; Haddadzadeh, H.; Shayan, P. Seroprevalence of Neospora caninum and Toxoplasma gondii in camels (Camelus dromedarius) in Mashhad, Iran. Parasitol. Res. 2006, 98, 600–601. [Google Scholar] [CrossRef]
- Parameswaran, N.; O’Handley, R.M.; Grigg, M.E.; Fenwick, S.G.; Thompson, R.C.A. Seroprevalence of Toxoplasma gondii in wild kangaroos using an ELISA. Parasitol. Int. 2009, 58, 161–165. [Google Scholar] [CrossRef]
- Liu, J.; Cai, J.Z.; Zhang, W.; Liu, Q.; Chen, D.; Han, J.P.; Liu, Q.R. Seroepidemiology of Neospora caninum and Toxoplasma gondii infection in yaks (Bos grunniens) in Qinghai, China. Vet. Parasitol. 2008, 152, 330–332. [Google Scholar] [CrossRef]
- Shapiro, K.; VanWormer, E.; Packham, A.; Dodd, E.; Conrad, P.A.; Miller, M. Type X strains of Toxoplasma gondii are virulent for southern sea otters (Enhydra lutris nereis) and present in felids from nearby watersheds. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191334. [Google Scholar] [CrossRef]
- Burgess, T.L.; Tinker, M.T.; Miller, M.A.; Bodkin, J.L.; Murray, M.J.; Saarinen, J.A.; Nichol, L.M.; Larson, S.; Conrad, P.A.; Johnson, C.K. Defining the risk landscape in the context of pathogen pollution: Toxoplasma gondii in sea otters along the Pacific Rim. R. Soc. Open Sci. 2018, 5, 171178. [Google Scholar] [CrossRef]
- Verma, S.K.; Knowles, S.; Cerqueira-Cézar, C.K.; Kwok, O.C.; Jiang, T.; Su, C.; Dubey, J.P. An update on Toxoplasma gondii infections in northern sea otters (Enhydra lutris kenyoni) from Washington State, USA. Vet. Parasitol. 2018, 258, 133–137. [Google Scholar] [CrossRef]
- Kirmse, P.; Ferguson, H. Toxoplasma-like organisms as the possible causative agents of a proliferative condition in farmed turbot (Scophthalmus maximus). In Wildlife Diseases; Springer: New York, NY, USA, 1976; pp. 561–564. [Google Scholar]
- Marino, A.M.F.; Giunta, R.P.; Salvaggio, A.; Castello, A.; Alfonzetti, T.; Barbagallo, A.; Aparo, A.; Scalzo, F.; Reale, S.; Buffolano, W.; et al. Toxoplasma gondii in edible fishes captured in the Mediterranean basin. Zoonoses Public Health 2019, 66, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Sroka, J.; Szymanska, J.; Wojcik-Fatla, A. The occurrence of Toxoplasma gondii and Borrelia burgdorferi sensu lato in Ixodes ricinus ticks from east Poland with the use of pcr. Ann. Agric. Environ. Med. 2009, 16, 313–319. [Google Scholar] [PubMed]
- Kijlstra, A.; Jongert, E. Toxoplasma-safe meat: Close to reality? Trends Parasitol. 2009, 25, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Sabin, A.B.; Feldman, H.A. Dyes as microchemical indicators of a new immunity phenomenon affecting a protozoon parasite (Toxoplasma). Science 1948, 108, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gasior, L. Toxoplasma gondii recombinant antigens as tools for serodiagnosis of human toxoplasmosis: Current status of studies. Clin. Vaccine Immunol. 2013, 20, 1343–1351. [Google Scholar] [CrossRef]
- Rostami, A.; Karanis, P.; Fallahi, S. Advances in serological, imaging techniques and molecular diagnosis of Toxoplasma gondii infection. Infection 2018, 46, 303–315. [Google Scholar] [CrossRef]
- Tenter, A.M.; Vietmeyer, C.; Johnson, A.M. Development of ELISAs based on recombinant antigens for the detection of Toxoplasma gondii-specific antibodies in sheep and cats. Vet. Parasitol. 1992, 43, 189–201. [Google Scholar] [CrossRef]
- Tenter, A.M.; Vietmeyer, C.; Johnson, A.M.; Janitschke, K.; Rommel, M.; Lehmacher, W. ELISAs based on recombinant antigens for seroepidemiological studies on Toxoplasma gondii infections in cats. Parasitology 1994, 109, 29–36. [Google Scholar] [CrossRef]
- Kimbita, E.N.; Xuan, X.; Huang, X.; Miyazawa, T.; Fukumoto, S.; Mishima, M.; Suzuki, H.; Sugimoto, C.; Nagasawa, H.; Fujisaki, K.; et al. Serodiagnosis of Toxoplasma gondii infection in cats by enzyme-linked immunosorbent assay using recombinant SAG1. Vet. Parasitol. 2001, 102, 35–44. [Google Scholar] [CrossRef]
- Huang, X.; Xuan, X.; Kimbita, E.N.; Battur, B.; Miyazawa, T.; Fukumoto, S.; Mishima, M.; Makala, L.H.; Suzuki, H.; Sugimoto, C.; et al. Development and evaluation of an enzyme-linked immunosorbent assay with recombinant SAG2 for diagnosis of Toxoplasma gondii infection in cats. J. Parasitol. 2002, 88, 804–807. [Google Scholar] [CrossRef]
- Huang, X.; Xuan, X.; Suzuki, H.; Sugimoto, C.; Nagasawa, H.; Fujisaki, K.; Mikami, T.; Igarashi, I. Characterization of Toxoplasma gondii SAG2 expressed in insect cells by recombinant baculovirus and evaluation of its diagnostic potential in an enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol. 2002, 9, 1343–1347. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xuan, X.; Hirata, H.; Yokoyama, N.; Xu, L.; Suzuki, N.; Igarashi, I. Rapid Immunochromatographic Test Using Recombinant SAG2 for Detection of Antibodies against Toxoplasma gondii in Cats. J. Clin. Microbiol. 2004, 42, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.K.; Jeong, W.; Kim, H.Y.; An, D.J.; Jeoung, H.Y.; Ryu, J.E.; Ko, A.R.; Kim, Y.J.; Hong, S.J.; Yang, Z.; et al. Development and clinical evaluation of a rapid serodiagnostic test for toxoplasmosis of cats using recombinant SAG1 antigen. Korean J. Parasitol. 2011, 49, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, Z.; Li, J.; Li, N.; Wei, F.; Liu, Q. Evaluation of an indirect elisa using recombinant granule antigen GRA7 for serodiagnosis of Toxoplasma gondii infection in cats. J. Parasitol. 2015, 101, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Liu, Y.; Chen, Y.; Yang, Q.; Chun, P.; Yao, K.; Han, X.; Wang, S.; Yu, S.; Liu, Y.; et al. A novel dynamic flow immunochromatographic test (DFICT) using gold nanoparticles for the serological detection of Toxoplasma gondii infection in dogs and cats. Biosens. Bioelectron. 2015, 72, 133–139. [Google Scholar] [CrossRef]
- Abdelbaset, A.E.; Alhasan, H.; Salman, D.; Karram, M.H.; Ellah Rushdi, M.A.; Xuenan, X.; Igarashi, M. Evaluation of recombinant antigens in combination and single formula for diagnosis of feline toxoplasmosis. Exp. Parasitol. 2017, 172, 1–4. [Google Scholar] [CrossRef]
- Ybañez, R.H.D.; Kyan, H.; Nishikawa, Y. Detection of antibodies against Toxoplasma gondii in cats using an immunochromatographic test based on GRA7 antigen. J. Vet. Med. Sci. 2020, 82, 441–445. [Google Scholar] [CrossRef]
- Wang, Z.; Ge, W.; Huang, S.Y.; Li, J.; Zhu, X.Q.; Liu, Q. Evaluation of recombinant granule antigens GRA1 and GRA7 for serodiagnosis of Toxoplasma gondii infection in dogs. BMC Vet. Res. 2014, 10, 158. [Google Scholar] [CrossRef]
- Zhou, M.; Cao, S.; Sevinc, F.; Sevinc, M.; Ceylan, O.; Liu, M.; Wang, G.; Moumouni, P.F.A.; Jirapattharasate, C.; Suzuki, H.; et al. Enzyme-linked immunosorbent assays using recombinant TgSAG2 and NcSAG1 to detect Toxoplasma gondii and Neospora caninum-specific antibodies in domestic animals in Turkey. J. Vet. Med. Sci. 2016, 78, 1877–1881. [Google Scholar] [CrossRef]
- Zhuo, X.; Sun, H.; Zhang, Z.; Luo, J.; Shan, Y.; Du, A. Development and application of an indirect enzyme-linked immunosorbent assay using recombinant MAG1 for serodiagnosis of Toxoplasma gondii in dogs. J. Parasitol. 2017, 103, 237–242. [Google Scholar] [CrossRef]
- Sager, H.; Gloor, M.; Tenter, A.; Maley, S.; Hässig, M.; Gottstein, B. Immunodiagnosis of primary Toxoplasma gondii infection in sheep by the use of a P30 IgG avidity ELISA. Parasitol. Res. 2003, 91, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Tumurjav, B.; Terkawi, M.A.; Zhang, H.; Zhang, G.; Jia, H.; Goo, Y.-K.; Yamagishi, J.; Nishikawa, Y.; Igarashi, I.; Sugimoto, C.; et al. Serodiagnosis of ovine toxoplasmosis in Mongolia by an enzyme-linked immunosorbent assay with recombinant Toxoplasma gondii matrix antigen 1. Jpn. J. Vet. Res. 2010, 58, 111–119. [Google Scholar] [PubMed]
- Holec-Gasior, L.; Ferra, B.; Hiszczyńska-Sawicka, E.; Kur, J. The optimal mixture of Toxoplasma gondii recombinant antigens (GRA1, P22, ROP1) for diagnosis of ovine toxoplasmosis. Vet. Parasitol. 2014, 206, 146–152. [Google Scholar] [CrossRef]
- Ferra, B.; Holec-Gąsior, L.; Kur, J. Serodiagnosis of Toxoplasma gondii infection in farm animals (horses, swine, and sheep) by enzyme-linked immunosorbent assay using chimeric antigens. Parasitol. Int. 2015, 64, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Tewari, A.K.; Mishra, A.K.; Maharana, B.; Sudan, V.; Raina, O.K.; Rao, J.R. Detection of antibodies to Toxoplasma gondii in domesticated ruminants by recombinant truncated SAG2 enzyme-linked immunosorbent assay. Trop. Anim. Health Prod. 2014, 47, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Tumurjav, B.; Nishikawa, Y.; Xuan, X. Seroprevalence of ovine toxoplasmosis in mongolian samples using an ELISA with recombinant Toxoplasma gondii matrix antigen 1. J. Agric. Sci. 2015, 15, 33–37. [Google Scholar] [CrossRef]
- Fereig, R.M.; Mahmoud, H.Y.A.H.; Mohamed, S.G.A.; AbouLaila, M.R.; Abdel-Wahab, A.; Osman, S.A.; Zidan, S.A.; El-Khodary, S.A.; Mohamed, A.E.A.; Nishikawa, Y. Seroprevalence and epidemiology of Toxoplasma gondii in farm animals in different regions of Egypt. Vet. Parasitol. Reg. Stud. Rep. 2016, 3–4, 1–6. [Google Scholar] [CrossRef]
- Holec-Gąsior, L.; Ferra, B.; Grąźlewska, W. Toxoplasma gondii tetravalent chimeric proteins as novel antigens for detection of specific immunoglobulin g in sera of small ruminants. Animals 2019, 9, 1146. [Google Scholar] [CrossRef]
- Ichikawa-Seki, M.; Guswanto, A.; Allamanda, P.; Mariamah, E.S.; Wibowo, P.E.; Igarashi, I.; Nishikawa, Y. Seroprevalence of antibody to TgGRA7 antigen of Toxoplasma gondii in livestock animals from Western Java, Indonesia. Parasitol. Int. 2015, 64, 484–486. [Google Scholar] [CrossRef]
- Pagmadulam, B.; Myagmarsuren, P.; Fereig, R.M.; Igarashi, M.; Yokoyama, N.; Battsetseg, B.; Nishikawa, Y. Seroprevalence of Toxoplasma gondii and Neospora caninum infections in cattle in Mongolia. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 11–17. [Google Scholar] [CrossRef]
- Sudan, V.; Tewari, A.K.; Singh, H. Detection of antibodies against Toxoplasma gondii in Indian Cattle by recombinant SAG2 enzyme-linked immunosorbent assay. Acta Parasitol. 2019, 64, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Bachan, M.; Deb, A.R.; Maharana, B.R.; Sudhakar, N.R.; Sudan, V.; Saravanan, B.C.; Tewari, A.K. High seroprevalence of Toxoplasma gondii in goats in Jharkhand state of India. Vet. Parasitol. Reg. Stud. Rep. 2018, 12, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Andrews, C.D.; Dubey, J.P.; Tenter, A.M.; Webert, D.W. Toxoplasma gondii recombinant antigens H4 and H11: Use in ELISAs for detection of toxoplasmosis in swine. Vet. Parasitol. 1997, 70, 1–11. [Google Scholar] [CrossRef]
- Gamble, H.R.; Andrews, C.D.; Dubey, J.P.; Webert, D.W.; Parmley, S.F. Use of recombinant antigens for detection of Toxoplasma gondii infection in swine. J. Parasitol. 2000, 86, 459–462. [Google Scholar] [CrossRef]
- Jiang, T.; Gong, D.; Ma, L.-A.; Nie, H.; Zhou, Y.; Yao, B.; Zhao, J. Evaluation of a recombinant MIC3 based latex agglutination test for the rapid serodiagnosis of Toxoplasma gondii infection in swines. Vet. Parasitol. 2008, 158, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gasior, L.; Kur, J.; Hiszczyńska-Sawicka, E.; Drapała, D.; Dominiak-Górski, B.; Pejsak, Z. Application of recombinant antigens in serodiagnosis of swine toxoplasmosis and prevalence of Toxoplasma gondii infection among pigs in Poland. Pol. J. Vet. Sci. 2010, 13, 457–464. [Google Scholar]
- Terkawi, M.A.; Kameyama, K.; Rasul, N.H.; Xuan, X.; Nishikawa, Y. Development of an immunochromatographic assay based on dense granule protein 7 for serological detection of Toxoplasma gondii infection. Clin. Vaccine Immunol. 2013, 20, 596–601. [Google Scholar] [CrossRef]
- Santana, S.S.; Gebrim, L.C.; Carvalho, F.R.; Barros, H.S.; Barros, P.C.; Pajuaba, A.C.A.M.; Messina, V.; Possenti, A.; Cherchi, S.; Reiche, E.M.V.; et al. CCp5A protein from Toxoplasma gondii as a serological marker of oocyst-driven infections in humans and domestic animals. Front. Microbiol. 2015, 6, 1305. [Google Scholar] [CrossRef]
- Liu, X.Y.; Wang, Z.D.; El-Ashram, S.; Liu, Q. Toxoplasma gondii oocyst-driven infection in pigs, chickens and humans in northeastern China. BMC Vet. Res. 2019, 15, 366. [Google Scholar] [CrossRef]
- Sun, X.; Wang, Z.; Li, J.; Wei, F.; Liu, Q. Evaluation of an indirect ELISA using recombinant granule antigen GRA1, GRA7 and soluble antigens for serodiagnosis of Toxoplasma gondii infection in chickens. Res. Vet. Sci. 2015, 100, 161–164. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; Abdel-Ghaffar, F.; Osman, G.Y.; El-Shourbagy, S.H.; Nishikawa, Y.; Khattab, R.A. Prevalence of Toxoplasma gondii in Chicken samples from delta of Egypt using ELISA, histopathology and immunohistochemistry. J. Parasit. Dis. 2016, 40, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Duong, H.D.; Appiah-Kwarteng, C.; Takashima, Y.; Aye, K.M.; Nagayasu, E.; Yoshida, A. A novel luciferase-linked antibody capture assay (LACA) for the diagnosis of Toxoplasma gondii infection in chickens. Parasitol. Int. 2020, 77, 102125. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.J.C.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of Toxoplasma infection in pregnant women: European multicentre case-control study. Br. Med. J. 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.M.; Pandit, A.; Golwilkar, A.; Williams, T.S. How do jains get Toxoplasma infection? Lancet 1999, 354, 486–487. [Google Scholar] [CrossRef]
- Kijlstra, A.; Jongert, E. Control of the risk of human toxoplasmosis transmitted by meat. Int. J. Parasitol. 2008, 38, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Ciaramella, P.; Corona, M.; Cortese, L.; Piantedosi, D.; Santoro, D.; Di Loria, A.; Rigato, R. Seroprevalence of Neospora spp. in asymptomatic horses in Italy. Vet. Parasitol. 2004, 123, 11–15. [Google Scholar] [CrossRef]
- Pitel, P.H.; Romand, S.; Pronost, S.; Foucher, N.; Gargala, G.; Maillard, K.; Thulliez, P.; Collobert-Laugier, C.; Tainturier, D.; Fortier, G.; et al. Investigation of Neospora sp. antibodies in aborted mares from Normandy, France. Vet. Parasitol. 2003, 118, 1–6. [Google Scholar] [CrossRef]
- Innes, E.A. Toxoplasmosis: Comparative species susceptibility and host immune response. Comp. Immunol. Microbiol. Infect. Dis. 1997, 20, 131–138. [Google Scholar] [CrossRef]
- Bekpen, C.; Hunn, J.P.; Rohde, C.; Parvanova, I.; Guethlein, L.; Dunn, D.M.; Glowalla, E.; Leptin, M.; Howard, J.C. The interferon-inducible p47 (IRG) GTPases in vertebrates: Loss of the cell autonomous resistance mechanism in the human lineage. Genome Biol. 2005, 6, R92. [Google Scholar] [CrossRef]
- Bernstein-Hanley, I.; Coers, J.; Balsara, Z.R.; Taylor, G.A.; Starnbach, M.N.; Dietrich, W.F. The p47 GTPases Igtp and Irgb10 map to the Chlamydia trachomatis susceptibility locus Ctrq-3 and mediate cellular resistance in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 14092–14097. [Google Scholar] [CrossRef]
- Collazo, C.M.; Yap, G.S.; Sempowski, G.D.; Lusby, K.C.; Tessarollo, L.; Vande Woude, G.F.; Sher, A.; Taylor, G.A. Inactivation of LRG-47 and IRG-47 reveals a family of interferon γ-inducible genes with essential, pathogen-specific roles in resistance to infection. J. Exp. Med. 2001, 194, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Howard, J. The IRG proteins: A function in search of a mechanism. Immunobiology 2008, 213, 367–375. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D.; Taylor, G.A.; McKinney, J.D. Immune control of tuberculosis by IFN-γ-inducible LRG-47. Science 2003, 302, 654–659. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D. Immune control of phagosomal bacteria by p47 GTPases. Curr. Opin. Microbiol. 2005, 8, 74–82. [Google Scholar] [CrossRef]
- Taylor, G.A.; Collazo, C.M.; Yap, G.S.; Nguyen, K.; Gregorio, T.A.; Taylor, L.S.; Eagleson, B.; Secrest, L.; Southon, E.A.; Reid, S.W.; et al. Pathogen-specific loss of host resistance in mice lacking the IFN-γ-inducible gene IGTP. Proc. Natl. Acad. Sci. USA 2000, 97, 751–755. [Google Scholar] [CrossRef]
- Taylor, G.A. IRG proteins: Key mediators of interferon-regulated host resistance to intracellular pathogens. Cell. Microbiol. 2007, 9, 1099–1107. [Google Scholar] [CrossRef]
- Zhao, Y.O.; Rohde, C.; Lilue, J.T.; Könen-Waisman, S.; Khaminets, A.; Hunn, J.P.; Howard, J.C. Toxoplasma gondii and the immunity-related GTPase (IRG) resistance system in mice—A review. Mem. Inst. Oswaldo Cruz 2009, 104, 234–240. [Google Scholar] [CrossRef]
- Araujo, F.G.; Williams, D.M.; Grumet, F.C.; Remington, J.S. Strain-dependent differences in murine susceptibility to Toxoplasma. Infect. Immun. 1976, 13, 1528–1530. [Google Scholar] [CrossRef]
- McLeod, R.; Eisenhauer, P.; Mack, D.; Brown, C.; Filice, G.; Spitalny, G. Immune responses associated with early survival after peroral infection with Toxoplasma gondii. J. Immunol. 1989, 142, 3247–3255. [Google Scholar]
- Suzuki, Y.; Yang, Q.; Remington, J.S. Genetic resistance against acute toxoplasmosis depends on the strain of Toxoplasma gondii. J. Parasitol. 1995, 81, 1032–1034. [Google Scholar] [CrossRef]

{kind=link}
| Method | Advantages | Disadvantages |
|---|---|---|
| Western blotting (WB) | Reference method Detection of IgG and IgM class antibodies Repeatability of results | Time-consuming |
| Specialised equipment needed | ||
| High-level of operator experience required | ||
| Immunofluorescence antibody test (IFAT) | Reference method | Microscope required Visual reading of results Possibility of cross-reactions Test based on T. gondii tachyzoites |
| Detection of specific antibodies | ||
| The ability to determine antibody titres | ||
| Detection of antibody classes | ||
| Repeatability of results | ||
| Direct agglutination test (DAT) | High sensitivity and specificity Detection of IgG and IgM class antibodies Simplicity of implementation | High IgG titres may mask the presence of IgM |
| 2-mercaptoethanol needed to bind natural and IgM antibodies | ||
| Test based on T. gondii tachyzoites | ||
| Latex agglutination test (LAT) | Simplicity of implementation | No possibility to differentiate classes of antibodies |
| Modified agglutination test (MAT) | High sensitivity and specificity Detection of IgG and IgM class antibodies Simplicity of implementation | High IgG titres may mask the presence of IgM |
| 2-mercaptoethanol needed to bind natural and IgM antibodies | ||
| Test based on T. gondii tachyzoites | ||
| ELISA | Quantitative and qualitative determination of antibodies and antigens in biological material | Specialised equipment needed Quality of the test dependent on antigen preparation |
| High sensitivity and specificity | ||
| Detection of all class antibodies | ||
| Automation |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| H4; H11; M: H4 + H11 | E. coli | IgG ELISA | In-house TEA-ELISA | 32 serum samples were tested. | [68] Germany |
| H4-ELISA: sensitivity 100%; specificity 100%. | |||||
| H11-ELISA: sensitivity 50%; specificity 100%. | |||||
| M-ELISA: sensitivity 100%; specificity 100%. | |||||
| TEA-ELISA: sensitivity 100%; specificity 100%. | |||||
| H4; H11; M: H4 + H11 | E. coli | IgG ELISA | In-house TEA-ELISA IFAT | 306 serum samples were tested. | [69] Germany |
| H4-ELISA: sensitivity 93%; specificity 100%. | |||||
| H11-ELISA: sensitivity 64%; specificity 100%. | |||||
| M-ELISA: sensitivity 95%; specificity 100%. | |||||
| TEA-ELISA: sensitivity 98%, specificity 99%. | |||||
| IFAT: sensitivity 94%; specificity 92%. | |||||
| SAG1 | E. coli | IgG ELISA | LAT WB | 193 serum samples were tested. | [70] Japan |
| Using the commercial ELISA and LAT tests the serum samples were divided into positive (40) and negative (153). | |||||
| SAG1-ELISA: positive results for 40 (20.7%) serum samples. | |||||
| 1 serum sample was positive in LAT, negative in WB and SAG1-ELISA. | |||||
| 1 serum sample was in LAT, positive in WB and SAG1-ELISA. | |||||
| SAG1; SAG2 | E. coli | IgG ELISA | LAT ELISA | 192 serum samples were tested. | [71] Japan |
| Using the commercial LAT test the serum samples were divided into positive (39) and negative (153). | |||||
| 40 (20.8%) of serum samples were positive in SAG1-ELISA. | |||||
| 42 (21.9%) of serum samples were positive in SAG2-ELISA. | |||||
| 38 (19.8%) of serum samples were positive in both SAG1-ELISA and SAG2-ELISA. | |||||
| 39 (20.3%) of serum samples were positive in LAT. | |||||
| 41 (21.4%) serum samples were positive in both SAG2-ELISA and WB. | |||||
| SAG2 | E. coli (E) Baculovirus (B) | IgG ELISA | LAT | 187 serum samples were tested. | [72] Japan |
| Using the commercial LAT test the serum samples were divided into positive (35) and negative (152). | |||||
| 53 (28.3%) of serum samples were positive in S-SAG2-ELISA. | |||||
| 36 (19.7%) of serum samples were positive in E-SAG2-ELISA (tested group of serum samples 183). | |||||
| 35 (18.7%) of serum samples were positive results in LAT. | |||||
| The authors suggest that the B-SAG2 antigen is more reactive, but in view of the large discrepancy with the commercial LAT test, it is difficult to agree with this statement. | |||||
| SAG2 | E. coli | IgG ELISA IgG ICT | LAT | 179 serum samples were tested. | [73] Japan |
| 41 (22.9%) of serum samples were positive in SAG2-ICT. | |||||
| 33 (18.4%) of serum samples were positive in LAT, constituting 95.5% compliance with SAG2-ICT. | |||||
| 35 (19.6%) of serum samples were positive in SAG2-ELISA, constituting 96.1% compliance with SAG2-ICT. | |||||
| SAG1; SAG2; GRA6 | E. coli | IgG RDT | ELISA | 182 serum samples were tested. Preliminary tests showed that SAG2 and GRA6 recombinant proteins are characterised by low reactivity. SAG1-RDT: sensitivity 100%; specificity 99.2%. Commercial ELISA: specificity 91.2%; specificity 98.7%. | [74] Korea |
| GRA7 | E. coli | IgG ELISA | MAT IFAT In-house TLA-ELISA | 185 serum samples were tested. Using the commercial IFAT and MAT tests the serum samples were divided into positive (39) and negative (146). TLA-ELISA: sensitivity 84.6%; specificity 99.3%. GRA7-ELISA: sensitivity 89.7%; specificity 92.5%. | [75] China |
| M: SAG1 + SAG2 | E. coli | DFICT | ELISA | 97 serum samples were tested. | [76] China |
| Using the commercial ID Screen® Toxoplasmosis indirect multi-species ELISA test, the serum samples were divided into positive (28) and negative (68). | |||||
| DFICT-M: sensitivity 92%, specificity 93.1%. | |||||
| SAG2, GRA2, GRA6; GRA7; GRA15; MIC10; M1: GRA6 + GRA7; M2: GRA2 + GRA7; M3: SAG2 + GRA7; M4: SAG2 + GRA6; M5: GRA2 + GRA6 + GRA7 + GRA15; M6: SAG2 + GRA2 + GRA6 + GRA7 + GRA15 | E. coli | IgG ELISA | LAT In-house TLA-ELISA | 419 serum samples were tested. Using the commercial LAT test the serum samples were divided into positive (73) and negative (346). TLA-ELISA: sensitivity 97.3%; specificity 93.6%. SAG2-ELISA: sensitivity 91.9%; specificity 88.1%. GRA2-ELISA: sensitivity 27.0%; specificity 96.5%. GRA6-ELISA: sensitivity 82.4%; specificity 88.7%. GRA7-ELISA: sensitivity 35.1%; specificity 89.9%. GRA15-ELISA: sensitivity 17.6%; specificity 86.4%. MIC10-ELISA: sensitivity 16.2%; specificity 85.8%. M1-ELISA: sensitivity 74.3%; specificity 89.0%. M2-ELISA: sensitivity 44.6%; specificity 89.3%. M3-ELISA: sensitivity 90.5%; specificity 85.5%. M4-ELISA: sensitivity 94.6%; specificity 89.6%. M5-ELISA: sensitivity 70.3%; specificity 86.1%. M6-ELISA: sensitivity 89.2%; specificity 95.4%. | [77] Egypt |
| GRA7 | E. coli | IgG ICT IgG ELISA | LAT In-house TLA-ELISA | 100 serum samples were tested. | [78] Japan |
| Using the commercial LAT test the serum samples were divided into positive (76) and negative (24). | |||||
| 74 of serum samples were positive in GRA7-ELISA. | |||||
| 74 of serum samples were positive in GRA7-ICT. | |||||
| 71 of serum samples were positive in TLA RH-ELISA. | |||||
| 70 of serum samples were positive in TLA PLK-ELISA. | |||||
| 70 of serum samples were positive in TLA VEG-ELISA. | |||||
| The authors suggest that two serum samples produced a false positive result in the LAT test., but their tests were based on the GRA7 protein are characterised by 100% sensitivity and specificity. |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| GRA1; GRA7 | E. coli | IgG ELISA | MAT IFAT In-house TLA-ELISA | 259 serum samples were tested. | [79] China |
| Using the commercial MAT and IFAT tests, the serum samples were divided into positive (44) and negative (215). | |||||
| GRA1-ELISA: sensitivity 81%, specificity 95.4%. | |||||
| GRA7-ELISA: sensitivity 91%, specificity 97.7%. | |||||
| TLA-ELISA: sensitivity 88.1%, specificity 96.8%. | |||||
| M: SAG1 + SAG2 | E. coli | DFICT | ELISA | 241 serum samples were tested. Using the commercial ID Screen® Toxoplasmosis indirect multi-species ELISA test. the serum samples were divided into positive (57) and negative (184). DFICT-M: sensitivity 96.2%, specificity 96.8%. | [76] China |
| SAG2 | E. coli | IgG ELISA | - | 187 serum samples were tested. | [80] Japan Turkey |
| 37 (24.1%) serum samples were positive for the presence of anti-T. gondii antibodies. | |||||
| Serum samples were not tested by any commercial assay. | |||||
| MAG1 | E. coli | IgG ELISA | WB In-house TLA-ELISA | 93 serum samples were tested. | [81] China |
| Using the Western blotting test with TLA, the serum samples were divided into positive (33) and negative (60). | |||||
| MAG1-ELISA: sensitivity 93.9%, specificity 98.3%. | |||||
| TLA-ELISA: sensitivity 87.8%, specificity 96.7%. |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| H4; H11; M: H4 + H11 | E. coli | IgG ELISA | In-house TEA-ELISA | 26 serum samples were tested. | [68] Germany |
| H4-ELISA: sensitivity 79%; specificity 100%. | |||||
| H11-ELISA: sensitivity 43%; specificity 100%. | |||||
| M-ELISA: sensitivity 79%; specificity 100%. | |||||
| TEA-ELISA: sensitivity100%; specificity 100%. | |||||
| H11 | E. coli | IgG ELISA | In-house TEA-ELISA In-house P30-ELISA In-house SA-ELISA | 92 serum samples were tested. H11-ELISA: sensitivity 34%; specificity 89%. P30-ELISA: sensitivity 96%; specificity 100%. TEA-ELISA: sensitivity 96%; specificity 100%. | [82] Switzerland |
| MAG1 | E. coli | IgG ELISA | LAT | 175 serum samples were tested. | [83] Japan |
| 29 (16.6%) serum samples were positive in LAT test. | |||||
| 42 (24%) serum samples were positive in MAG1-ELISA. | |||||
| 27 (15.4%) serum samples had the same results in LAT test and MAG1-ELISA. | |||||
| No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of MAG1 protein. | |||||
| GRA1; GRA2ex2; GRA4; GRA5; GRA6; GRA9; SAG1; SAG2; SAG4; BSR4; ROP1; ROP9; MIC1ex2; MIC1ex34; MIC3; MAG1; BAG1; LDH1; LDH2; M1: GRA1 + ROP1; M2: GRA1 + SAG2; M3: SAG2 + ROP1; M4: GRA1 + SAG2 + ROP1 | E. coli | IgG ELISA | IFAT MAT In-house TLA-ELISA | Preliminary evaluation of single recombinant antigens in IgG ELISA with 2 positive and 2 negative serum samples. 8 antigens (GRA1, GRA9, SAG1, SAG2, SAG4, MIC1ex2, MIC3, and ROP1) were selected for further analysis with pool of 108 serum samples. 3 antigens (GRA1, SAG2, ROP1) were selected with the sensitivity (98.9–100%) and specificity (100%). Mixtures of these antigens were tested with the same pool of 108 serum samples. M1-ELISA: sensitivity 100%, specificity 100%. M2-ELISA: sensitivity 100%, specificity 95%. M3-ELISA: sensitivity 100%, specificity 95%. M4-ELISA: sensitivity 100%, specificity 100%. M4 was characterised by the highest reactivity. To confirm this thesis, a test with a new 128 serum samples was carried out, again sensitivity and specificity were 100%. All serum samples was tested in TLA-ELISA: sensitivity 100%; specificity 100%. Serum samples previously tested with MAT and IFAT. | [84] Poland |
| M1: SAG1 + MIC1 + MAG1; M2: SAG2 + GRA1 + ROP1; M3: GRA1 + GRA2 + GRA6 Chimeric antigens: MIC1-MAG1-SAG1S; SAG1L-MIC1-MAG1; SAG2-GRA1-ROP1S; SAG2-GRA1-ROP1L; GRA1-GRA2-GRA6 | E. coli | IgG ELISA | IFAT MAT In-house TLA-ELISA | 191 serum samples were tested. M1-ELISA: sensitivity 77.9%; specificity 92.2%. M2-ELISA: sensitivity 100%; specificity 100%. M3-ELISA: sensitivity 92.1%; specificity 100%. MIC1-MAG1-SAG1S-ELISA: sensitivity 97.9%. SAG1L-MIC1-MAG1-ELISA: sensitivity 100%. SAG2-GRA1-ROP1S-ELISA: sensitivity 100%. SAG2-GRA1-ROP1L-ELISA: sensitivity 100%. GRA1-GRA2-GRA6-ELISA: sensitivity 92.1%. Specificity of all IgG ELISA with recombinant chimeric antigens were 100%. TLA-ELISA: sensitivity 100%; specificity 100%. Serum samples previously tested with MAT and IFAT. | [85] Poland |
| SAG2 | E. coli | IgG ELISA | IFAT | 60 serum samples were tested. | [86] India |
| Compared to IFAT, the SAG2-ELISA had 81.3% sensitivity and 85.7% specificity. | |||||
| MAG1 | E. coli | IgG ELISA | LAT | 175 serum samples were tested. 29 (16.6%) serum samples were positive in LAT test. 42 (24%) serum samples positive in MAG1-ELISA. 27 (15.4%) serum samples produced the same results in LAT test and MAG1-ELISA. No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of MAG1 protein. | [87] Japan |
| SAG2 | E. coli | IgG ELISA | - | 610 serum samples were tested. 122 (20%) serum samples were positive for the presence of anti-T. gondii antibodies. Serum samples were not tested by any commercial assay. | [80] Japan Turkey |
| GRA7 | E. coli | IgG ELISA | LAT | 111 serum samples were tested. | [88] Egypt |
| T. gondii infection was detected in 53 (47.8%) serum samples using the LAT test. | |||||
| T. gondii infection was detected in 57 (51.4%) serum samples using the GRA7-ELISA. | |||||
| The same result were obtained for both LAT and GRA7 ELISA for 43 (38.7%) serum samples. | |||||
| No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of GRA7 protein. | |||||
| Chimeric antigens: AMA1N-SAG2-GRA1-ROP1; AMA1C-SAG2-GRA1-ROP1; AMA1-SAG2-GRA1-ROP1; SAG2-GRA1-ROP1-GRA2 | E. coli | IgG ELISA | IFAT MAT In-house TLA-ELISA | 90 serum samples were tested. AMA1N-SAG2-GRA1-ROP1-ELISA: sensitivity 97.9%, specificity 97.6%. AMA1C-SAG2-GRA1-ROP1-ELISA: sensitivity 95.8%, specificity 95.2%. AMA1-SAG2-GRA1-ROP1-ELISA: sensitivity 97.9%, specificity 100%. SAG2-GRA1-ROP1-GRA2-ELISA: sensitivity 97.9%, specificity 97.6%. TLA-ELISA: sensitivity 97.9%, specificity 100%. Serum samples previously tested with MAT and IFAT. | [89] Poland |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| GRA7 | E. coli | IgG ELISA | IFA WB In house TLA-ELISA | 101 serum samples were tested. GRA7-ELISA identified 28 positive and 73 negative samples. TLA-ELISA identified 22 positive and 79 negative samples. Results were compared with those obtained with IFA and WB, which showed that the GRA7-ELISA is the best method. GRA7-ELISA: sensitivity 96.4%; specificity 98.6%. TLA-ELISA: sensitivity 95.5%; specificity 91.1%. | [79] China |
| SAG2 | E. coli | IgG ELISA | IFAT | 45 serum samples were tested. | [86] India |
| Compared to IFAT, the SAG2-ELISA had 87.1% sensitivity and 85.7% specificity. | |||||
| GRA7 | E. coli | IgG ELISA | - | 598 serum samples were tested. | [90] Japan |
| 44 (7.4%) serum samples were considered positive. | |||||
| No reference tests were performed (neither commercial nor in-house TLA based). | |||||
| SAG2 | E. coli | IgG ELISA | - | 377 serum samples were tested. | [80] Japan Turkey |
| 15 (4%) serum samples were positive for the presence of anti-T. gondii antibodies. | |||||
| Serum samples were not tested by any commercial assay. | |||||
| GRA7 | E. coli | IgG ELISA | LAT | 301 serum samples were tested. | [88] Egypt |
| T. gondii infection was detected in 88 (29.2%) serum samples using the LAT test. | |||||
| T. gondii infection was detected in 85 (28.2%) serum samples using the GRA7-ELISA. | |||||
| The same result were obtained for both LAT and GRA7 ELISA for 71 (23.6%) serum samples. | |||||
| No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of GRA7 protein. | |||||
| GRA7 | E. coli | IgG ELISA | - | 1438 serum samples were tested. | [91] Japan |
| 269 (18.7%) serum samples were considered positive. | |||||
| No reference tests were performed (neither commercial nor in-house TLA based). | |||||
| SAG2 | E. coli | IgG ELISA | IFAT | 258 serum samples were tested. | [92] India |
| 115 (44.6%) serum samples give positive results for the presence of anti-T. gondii antibodies. | |||||
| Compared to IFAT, the SAG2-ELISA had 80% sensitivity and 88.6% specificity. |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| SAG2 | E. coli | IgG ELISA | IFAT | 63 serum samples were tested. | [86] India |
| Compared to IFAT, the SAG2-ELISA had 82.1% sensitivity and 91.4% specificity. | |||||
| SAG2 | E. coli | IgG ELISA | - | 249 serum samples were tested. | [80] Japan Turkey |
| 32 (12.9%) serum samples were positive for the presence of anti-T. gondii antibodies. | |||||
| Serum samples were not tested by any commercial assay. | |||||
| GRA7 | E. coli | IgG ELISA | LAT | 94 serum samples were tested. T. gondii infection was detected in 33 (35.1%) serum samples using the LAT test. T. gondii infection was detected in 37 (39.4%) serum samples using the GRA7-ELISA. The same result was obtained for both LAT and GRA7 ELISA for 27 (28.7%) serum samples. No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of GRA7 protein. | [88] Egypt |
| SAG1 | E. coli | IgG ELISA | IFAT | 445 serum samples were tested. | [93] India |
| 189 (42.5%) serum samples were positive for the presence of anti-T. gondii antibodies in SAG1-ELISA. | |||||
| Relative to the results obtained in the IFAT, the sensitivity and specificity of the SAG1-ELISA was 92.7% and 90.7%, respectively. | |||||
| Chimeric antigens: AMA1N-SAG2-GRA1-ROP1; AMA1C-SAG2-GRA1-ROP1; AMA1-SAG2-GRA1-ROP1; SAG2-GRA1-ROP1-GRA2 | E. coli | IgG ELISA | IFAT MAT In-house TLA-ELISA | 86 serum samples were tested. AMA1N-SAG2-GRA1-ROP1-ELISA: sensitivity 88.9%, specificity 100%. AMA1C-SAG2-GRA1-ROP1-ELISA: sensitivity 95.6%, specificity 97.6%. AMA1-SAG2-GRA1-ROP1-ELISA: sensitivity 95.6%, specificity 100%. SAG2-GRA1-ROP1-GRA2-ELISA: sensitivity 57.8%, specificity 95.1%. TLA-ELISA: sensitivity 97.8%, specificity 100%. Serum samples were previously tested with MAT and IFAT. | [89] Poland |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| H4; H11 | E. coli | IgG ELISA | MAT In-house NAE-ELISA | Recombinant antigens are better recognised by specific antibodies contained in some samples of sera from pigs up to 9 weeks after infection with T. gondii. | [94] USA |
| Recombinant antigens are generally less reactive than the native antigen extract, in particular, the reactivity with sera takes longer to develop after T. gondii infection. | |||||
| B427 (MAG1); C55 (GRA2); V22 (SAG2); MBP30 (SAG1); C51 | E. coli | IgG ELISA | In-house NTA-ELISA | Antigens with the best properties are C51, V22 and MBP30 based on the difference between the values obtained for negative and positive sera. | [95] USA |
| C51 and MBP30 antigens were correctly recognised by specific antibodies in sera of pigs infected with T. gondii 52 weeks after infection. | |||||
| MIC3; SAG1 | E. coli | IgG LAT IgG ELISA | - | MIC3-LAT test correctly identified all 50 seropositive sera from experimentally infected pigs; the test specificity was 100%. | [96] China |
| SAG1-ELISA test correctly identified 48 seropositive sera; the test specificity was 100%. | |||||
| Examining 256 undefined samples of pig sera using the MIC3-LAT test were identified 103 as positive; specific anti-T. gondii antibodies were detected in 94 sera in the SAG1-ELISA. | |||||
| Positive agreement between MIC3-LAT and SAG1-ELISA was 91.3% for undefined serum samples, and 92.8% for all sera used. | |||||
| MAG1; SAG1; GRA7; M1: MAG1 + SAG1; M2: MAG1 + GRA7; M3: SAG1 + GRA7; M4: MAG1 + SAG1 + GRA7 | E. coli | IgG ELISA | MAT In-house TLA-ELISA | 120 serum samples were tested. | [97] Poland |
| MAG1-ELISA: sensitivity 64%. SAG1-ELISA: sensitivity 85.3%. | |||||
| GRA7-ELISA: sensitivity 81.3%. | |||||
| M1-ELISA: sensitivity 86.7%. | |||||
| M2-ELISA: sensitivity 89.2%. | |||||
| M3-ELISA: sensitivity 92%. | |||||
| M4-ELISA: sensitivity 97.3%. | |||||
| Specificity of all IgG ELISA with recombinant antigens was 100%. | |||||
| TLA-ELISA: sensitivity 100%; specificity 100%. | |||||
| Serum samples were previously tested with MAT and IFAT. | |||||
| SAG2; GRA7; GRA14 | E. coli | IgG ELISA IgM ELISA IgG ICT | LAT | From based on recombinant antigens, the GRA7 antigen is best recognised by anti-T. gondii antibodies. | [98] Japan |
| Results of the commercial LAT test indicated that the sensitivity of the GRA7-ELISA was 90.6%, the specificity was 85.2%, and the compliance was 88.1%. | |||||
| Results of the commercial LAT test, indicated that the sensitivity of the GRA7-ICT test was 71.9%, the specificity was 96.3%, and the compliance was 83.1%. | |||||
| M1: SAG1 + MIC1 + MAG1; M2: SAG2 + GRA1 + ROP1; M3: GRA1 + GRA2 + GRA6; Chimeric antigens: MIC1-MAG1-SAG1S; SAG1L-MIC1-MAG1; SAG2-GRA1-ROP1S; SAG2-GRA1-ROP1L; GRA1-GRA2-GRA6 | E. coli | IgG ELISA | IFAT MAT In-house TLA-ELISA | 168 serum samples were tested. M1-ELISA: sensitivity 88.9%; specificity 100%. M2-ELISA: sensitivity 81.5%; specificity 100%. M3-ELISA: sensitivity 54.3%; specificity 100%. MIC1-MAG1-SAG1S-ELISA: sensitivity 45.7%. SAG1L-MIC1-MAG1-ELISA: sensitivity 90.1%. SAG2-GRA1-ROP1S-ELISA: sensitivity 28.4%. SAG2-GRA1-ROP1L-ELISA: sensitivity 93.8%. GRA1-GRA2-GRA6-ELISA: sensitivity 96.3%. Specificity of all IgG ELISA with recombinant chimeric antigens was 100%. TLA-ELISA: sensitivity 100%; specificity 100%. Serum samples previously tested with MAT and IFAT. | [85] Poland |
| GRA7 | E. coli | IgG ELISA | - | 205 serum samples were tested. | [90] Japan |
| 30 (14.6%) serum samples were considered positive. | |||||
| No reference tests were performed (neither commercial nor in-house TLA based). | |||||
| CCp5A; OWP1 | E. coli | IgG WB IgG ELISA | WB In-house STAg-ELISA | 44 serum samples were tested. | [99] Brazil |
| CCp5A-WB: detection of infection. | |||||
| OWP1-WB: no detection of infection. | |||||
| STAg-WB: detection of infection. | |||||
| CCp5A-ELISA: sensitivity 100%, specificity undefined. | |||||
| STAg-ELISA: sensitivity 100%, specificity undefined. | |||||
| No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of recombinant proteins. | |||||
| GRA7; OWP8; CCp5A | E. coli | IgG ELISA | - | 90 serum samples were tested. | [100] China |
| GRA7-ELISA: sensitivity 16.7%, specificity undefined. | |||||
| OWP8-ELISA: sensitivity 12,2%, specificity undefined. | |||||
| CCp5A-ELISA: sensitivity 12.2%, specificity undefined. | |||||
| No tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of recombinant proteins. |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| M1: SAG1 + MIC1 + MAG1; M2: SAG2 + GRA1 + ROP1; M3: GRA1 + GRA2 + GRA6; Chimeric antigens: MIC1-MAG1-SAG1S; SAG1L-MIC1-MAG1; SAG2-GRA1-ROP1S; SAG2-GRA1-ROP1L; GRA1-GRA2-GRA6 | E. coli | IgG ELISA | IFAT MAT In-house TLA-ELISA | 86 serum samples were tested. M1-ELISA: sensitivity 88.9%; specificity 100%. M2-ELISA: sensitivity 77.8%; specificity 100%. M3-ELISA: sensitivity 66.7%; specificity 100%. MIC1-MAG1-SAG1S-ELISA: sensitivity 75%. SAG1L-MIC1-MAG1-ELISA: sensitivity 77.8%. SAG2-GRA1-ROP1S-ELISA: sensitivity 50%. SAG2-GRA1-ROP1L-ELISA: sensitivity 100%. GRA1-GRA2-GRA6-ELISA: sensitivity 86.1%. Specificity of all IgG ELISA with recombinant chimeric antigens was 100%. TLA-ELISA: sensitivity 100%; specificity 100%. | [85] Poland |
| SAG2 | E. coli | IgG ELISA | - | 616 serum samples were tested. 285 (46.3%) serum samples were positive for the presence of anti-T. gondii antibodies. Serum samples were not tested by any commercial assay. | [80] Japan Turkey |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| CCp5A; OWP1 | E. coli | IgY ELISA | WB In-house STAg-ELISA | 113 serum samples were tested. | [99] Brazil |
| CCp5A-ELISA: sensitivity 70%, specificity undefined. | |||||
| OWP1-ELISA: sensitivity 74% specificity undefined. | |||||
| STAg-ELISA: sensitivity 80%, specificity undefined. | |||||
| No tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of recombinant proteins. | |||||
| GRA1; GRA7 | E. coli | IgY ELISA | In-house TSA-ELISA | 110 serum samples were tested. | [101] China |
| GRA1-ELISA: sensitivity 81.3%, specificity 94.7%. | |||||
| GRA7-ELISA: sensitivity 100%, specificity 98.9%. | |||||
| TSA-ELISA: sensitivity 93.8%, specificity 97.9%. | |||||
| Serum samples were previously tested in WB to confirm the presence of anti-T. gondii antibodies. | |||||
| SAG2 | E. coli | IgY ELISA | Histopathology IHC | 304 serum samples were tested. | [102] Egypt |
| SAG2-ELISA: sensitivity 11.2%, specificity undefined. | |||||
| 6.9% cases confirmed by other testes—histopathology and IHC. | |||||
| Test compliance was 95.7%. | |||||
| GRA7; OWP8; CCp5A | E. coli | IgY ELISA | - | 96 serum samples were tested. | [100] China |
| GRA7-ELISA: Sensitivity 10.4%, specificity undefined. | |||||
| OWP8-ELISA: 13,5%, specificity undefined. | |||||
| CCp5A-ELISA: 9.4%, specificity undefined. | |||||
| No tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of recombinant proteins. | |||||
| GRA8 | E. coli | IgY LACA IgY ELISA | - | 86 serum samples were tested (21 from experimentally infected chicken and 65 from chicken without anti-T. gondii antibodies). Purified Nluc-GRA8-LACA: sensitivity 90.5%, specificity 95.4%. Unpurified Nluc-GRA8-LACA: sensitivity 85.7%, specificity 96.9%. GRA8-ELISA: sensitivity 85.7%, specificity 84.6%%. Screening Nluc-GRA8-LACA was performed using 267 sera from free-range chicken and 100 sera from closed-house broiler chicken. Presence of anti-T. gondii antibodies was found in 10.9% of free-range chickens. | [103] Japan |
| Antigen | Expression System | Assay | Reference Test | Results | Reference Country |
|---|---|---|---|---|---|
| GRA7 | E. coli | IgG ELISA | LAT | 146 serum samples were tested. T. gondii infection was detected in 39 (26.7%) serum samples using the LAT test. T. gondii infection was detected in 42 (28.8%) serum samples using the GRA7-ELISA. The same result were obtained for both LAT and GRA7 ELISA for 27 (22.6%) serum samples. No additional tests were performed to confirm the presence of anti-T. gondii antibodies, so it is difficult to determine the diagnostic utility of GRA7 protein. | [88] Egypt |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferra, B.; Holec-Gąsior, L.; Grąźlewska, W. Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals. Animals 2020, 10, 1245. https://doi.org/10.3390/ani10081245
Ferra B, Holec-Gąsior L, Grąźlewska W. Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals. Animals. 2020; 10(8):1245. https://doi.org/10.3390/ani10081245
Chicago/Turabian StyleFerra, Bartłomiej, Lucyna Holec-Gąsior, and Weronika Grąźlewska. 2020. "Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals" Animals 10, no. 8: 1245. https://doi.org/10.3390/ani10081245
APA StyleFerra, B., Holec-Gąsior, L., & Grąźlewska, W. (2020). Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals. Animals, 10(8), 1245. https://doi.org/10.3390/ani10081245