Estimating Abundance of Siberian Roe Deer Using Fecal-DNA Capture-Mark-Recapture in Northeast China

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
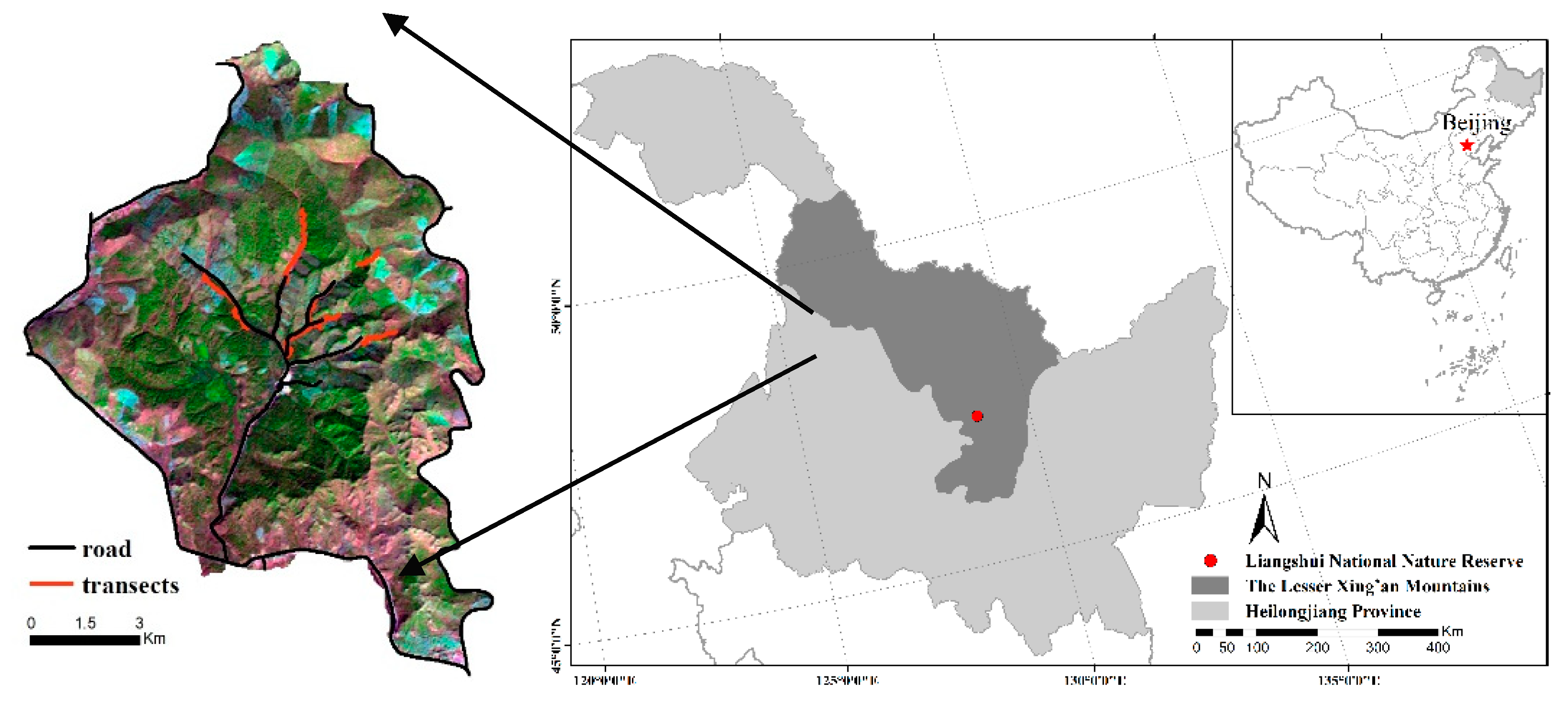
2.1. Study Area
2.2. Sampling Design
2.3. Genetic Analysis
2.3.1. DNA Extraction
2.3.2. Microsatellite Marker Selection
2.3.3. Genotyping and Individual Identification
2.4. Population Abundance Estimation
2.5. Population Density Estimation
3. Results
3.1. Fecal Sampling and Genotyping
3.2. Population Abundance Estimation
3.3. Population Density Estimation
4. Discussion
4.1. Sampling Design and Genetic Analysis
4.2. Population Abundance Model
4.3. Population Abundance and Density
5. Management Implications
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Locus | Sequences (5′-3′) | Length (bp) | A * | References |
|---|---|---|---|---|
| Roe1 | F AAATTTGGCTCTGCAATCGG R ACACAAAAGCCACCCAATAC | 112–132 | 7 | [68] |
| Roe6 | F GTTCCTAGCCCAGTGCTC R TGCAGACCTGGCAGAC | 89–109 | 3 | [68] |
| Roe8 | F AAGCCGCGCTTGAAGGAG R ATCAAGCTCCCCTCTTCG | 69–89 | 7 | [68] |
| Roe9 | F TTGGCGTCATTCCAACAGAG R TCACAGCAGAATGTCATCTG | 175–179 | 3 | [68] |
| IDVGA8 | F CTCTTGGGGGCGTGTTGTCT R TAGCAGAAAGCACAGGAGTC | 209–225 | 4 | [69] |
| BM757 | F TGGAAACAATGTAAACCTGGG R TTGAGCCACCAAGGAACC | 172–204 | 10 | [69] |
| MB25 | F GGACACGTTCTGCAGATACAACTAC R GAACTCTCCTTAAGCATACTTGCTC | 198–200 | 2 | [70] |
| OarFCB304 | F CCCTAGGAGCTTTCAATAAAGAATCGG R CGCTGCTGTCAACTGGGTCAGGG | 158–177 | 15 | [69] |
| BL42 | F CAAGGTCAAGTCCAAATGCC R GCATTTTTGTGTTAATTTCATGC | 225–231 | 3 | [70] |
| BM1818 | F AGCTGGGAATATAACCAAAGG R AGTGCTTTCAAGGTCCATGC | 249–267 | 3 | [70] |
| BMC1009 | F GCTTGGATGGACCATGTTG R CACTTGAGGGGCAAATGATT | 282–290 | 3 | [70] |
| BMS119 | F TTAAGCAGGGACGAACGTG R AATTGCCAGGAAGATTGTGG | 99–105 | 4 | [70] |
| ETH225 | F GATCACCTTGCCACTATTTCCT R ACATGACAGCCAGCTGCTACT | 139–145 | 4 | [70] |
| HUJ1177 | F TCCATCAAGTATTTGAGTGCAA R ATAGCCCTACCCACTGTTTCTG | 199–219 | 6 | [69] |
| ILST030Q | F CTGCAGTTCTGCATATGTGG R GTTTCTTCTTAGACAACAGGGGTTTGG | 157–165 | 3 | [70] |
| KCNA4 | F CTGGAAGAGATGTTAAAAGTA R CACTGAATAAACAACTGCTCA | 220–250 | 3 | [70] |
| MAF70 | F CACGGAGTCACAAAGAGTCAGACC R GCAGGACTCTACGGGGCCTTTGC | 139–161 | 5 | [70] |
| SR-CRSP1 | F TGCAAGAAGTTTTTCCAGAGC R TCATTCTCAGGAAACTCTGAAAC | 127–151 | 5 | [70] |
References
- Coulson, T.; Albon, S.; Guinness, F.; Pemberton, J.; CluttonBrock, T. Population substructure, local density, and calf winter survival in red deer (Cervus elaphus). Ecology 1997, 78, 852–863. [Google Scholar] [CrossRef]
- Roseberry, J.L.; Woolf, A. Habitat-population density relationships for white-tailed deer in Illinois. Wildl. Soc. Bull. 1998, 26, 252–258. [Google Scholar]
- Lounsberry, Z.T.; Forrester, T.D.; Olegario, M.T.; Brazeal, J.L.; Wittmer, H.U.; Sacks, B.N. Estimating Sex-Specific Abundance in Fawning Areas of a High-Density Columbian Black-Tailed Deer Population Using Fecal DNA. J. Wildl. Manag. 2015, 79, 39–49. [Google Scholar] [CrossRef]
- DeCalesta, D.S. Achieving and maintaining sustainable white-tailed deer density with adaptive management. Hum. Wildl. Interact. 2017, 11, 99–111. [Google Scholar]
- Kjellander, P.; Hewison, A.J.M.; Liberg, O.; Angibault, J.-M.; Bideau, E.; Cargnelutti, B. Experimental evidence for density-dependence of home-range size in roe deer (Capreolus capreolus L.): A comparison of two long-term studies. Oecologia 2004, 139, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Massé, A.; Côté, S.D. Habitat Selection of a Large Herbivore at High Density and Without Predation: Trade-Off Between Forage and Cover? J. Mammal. 2009, 90, 961–970. [Google Scholar] [CrossRef]
- Russell, F.L.; Zippin, D.B.; Fowler, N.L. Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: A review. Am. Midl. Nat. 2001, 146, 1–26. [Google Scholar] [CrossRef]
- Walter, W.D.; Vercauteren, K.C.; Iii, H.C.; Clark, W.R.; Fischer, J.W.; Hygnstrom, S.E.; Mathews, N.E.; Nielsen, C.K.; Schauber, E.M.; Deelen, T.R.V. Regional assessment on influence of landscape configuration and connectivity on range size of white-tailed deer. Landsc. Ecol. 2009, 24, 1405–1420. [Google Scholar] [CrossRef]
- Gibbs, J.P. Monitoring populations. In Research Techniques in Animal Ecology; Columbia University Press: New York, NY, USA, 2000; pp. 213–252. [Google Scholar]
- Yao, G.; Wang, B.-B.; Zhu, Y.; Wan, Q.-H.; Fang, S.-G. Low Population Density of the Endangered Forest Musk Deer, Moschus berezovskii, in China. Pak. J. Zool. 2015, 47, 325–333. [Google Scholar]
- Wu, Y. Ungulates Survey in Hunchun Nature Reserve, China, in the Winters of 2003/5. Master’s Thesis, East China Normal University, Shanghai, China, 2005. [Google Scholar]
- Zhang, C. Studies on Current Status of Amur Tiger’s Prey Population Resources and Habitat Use of Amur Tiger in Wanda Mountains of Heilongjiang Province. Master’s Thesis, Northeast Forestry University, Harbin, China, 2006. [Google Scholar]
- Foran, D.R.; Minta, S.C.; Heinemeyer, K.S. DNA-based analysis of hair to identify species and individuals for population research and monitoring. Wildl. Soc. Bull. (1973–2006) 1997, 25, 840–847. [Google Scholar]
- Wilson, G.; Frantz, A.; Pope, L.; Roper, T.; Burke, T.; Cheeseman, C.; Delahay, R. Estimation of badger abundance using faecal DNA typing. J. Appl. Ecol. 2003, 40, 658–666. [Google Scholar] [CrossRef]
- Flagstad, Ø.; Hedmark, E.; Landa, A.; Brøseth, H.; Persson, J.; Andersen, R.; Segerström, P.; Ellegren, H. Colonization history and noninvasive monitoring of a reestablished wolverine population. Conserv. Biol. 2004, 18, 676–688. [Google Scholar] [CrossRef]
- Hedmark, E.; Flagstad, Ø.; Segerström, P.; Persson, J.; Landa, A.; Ellegren, H. DNA-based individual and sex identification from wolverine (Gulo gulo) faeces and urine. Conserv. Genet. 2004, 5, 405–410. [Google Scholar] [CrossRef]
- Maudet, C.; Luikart, G.; Dubray, D.; Von Hardenberg, A.; Taberlet, P. Low genotyping error rates in wild ungulate faeces sampled in winter. Mol. Ecol. Resour. 2004, 4, 772–775. [Google Scholar] [CrossRef]
- Bellemain, E.; Swenson, J.E.; Tallmon, D.; Brunberg, S.; Taberlet, P. Estimating population size of elusive animals with DNA from hunter-collected feces: Four methods for brown bears. Conserv. Biol. 2005, 19, 150–161. [Google Scholar] [CrossRef]
- Brinkman, T.J.; Person, D.K.; Chapin, F.S., III; Smith, W.; Hundertmark, K.J. Estimating Abundance of Sitka Black-Tailed Deer Using DNA From Fecal Pellets. J. Wildl. Manag. 2011, 75, 232–242. [Google Scholar] [CrossRef]
- Marucco, F.; Boitani, L.; Pletscher, D.H.; Schwartz, M.K. Bridging the gaps between non-invasive genetic sampling and population parameter estimation. Eur. J. Wildl. Res. 2011, 57, 1–13. [Google Scholar] [CrossRef]
- Valiere, N.; Bonenfant, C.; Toïgo, C.; Luikart, G.; Gaillard, J.-M.; Klein, F. Importance of a pilot study for non-invasive genetic sampling: Genotyping errors and population size estimation in red deer. Conserv. Genet. 2007, 8, 69–78. [Google Scholar] [CrossRef]
- McCoy, K.R.; Pendleton, G.W.; Flynn, R.W. Assessing Population Estimation Protocols for Sitka Black-tailed Deer Using DNA from Fecal Pellets; Alaska Department of Fish and Game, Division of Wildlife Conservation: Juneau, AK, USA, 2014. [Google Scholar]
- Ebert, C.; Sandrini, J.; Spielberger, B.; Thiele, B.; Hohmann, U. Non-invasive genetic approaches for estimation of ungulate population size: A study on roe deer (Capreolus capreolus) based on faeces. Anim. Biodivers. Conserv. 2012, 35, 267–275. [Google Scholar]
- Hettinga, P.N.; Arnason, A.N.; Manseau, M.; Cross, D.; Whaley, K.; Wilson, P.J. Estimating size and trend of the North Interlake woodland caribou population using fecal-DNA and capture–recapture models. J. Wildl. Manag. 2012, 76, 1153–1164. [Google Scholar] [CrossRef]
- Goode, M.J.; Beaver, J.T.; Muller, L.I.; Clark, J.D.; van Manen, F.T.; Harper, C.A.; Basinger, P.S. Capture—Recapture of white-tailed deer using DNA from fecal pellet groups. Wildl. Biol. 2014, 20, 270–278. [Google Scholar] [CrossRef]
- Brazeal, J.L.; Weist, T.; Sacks, B.N. Noninvasive genetic spatial capture-recapture for estimating deer population abundance. J. Wildl. Manag. 2017, 81, 629–640. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, M. Population status and dynamic trends of Amur tiger’s prey in Eastern Wandashan Mountain, Heilongjiang Province. Acta Ecol. Sin. 2011, 31, 6481–6487. [Google Scholar]
- Xiao, W.; Feng, L.; Zhao, X.; Yang, H.; Dou, H.; Cheng, Y.; Mou, P.; Wang, T.; Ge, J. Distribution and abundance of Amur tiger, Amur leopard and their ungulate prey in Hunchun National Nature Reserve, Jilin. Biodivers. Sci. 2014, 22, 717–724. [Google Scholar]
- Anderson, D.R. The need to get the basics right in wildlife field studies. Wildl. Soc. Bull. 2001, 29, 1294–1297. [Google Scholar]
- Collier, B.A.; Ditchkoff, S.S.; Ruth, C.R., Jr.; Raglin, J.B. Spotlight surveys for white-tailed deer: Monitoring panacea or exercise in futility? J. Wildl. Manag. 2013, 77, 165–171. [Google Scholar] [CrossRef]
- Tian, X.; Zhang, M. Population size and sex ration of wapiti (Cervus elephus xanthopygus) as revealed by fecal DNA. Acta Ecol. Sin. 2010, 30, 6249–6254. [Google Scholar]
- Insisting on education for students and aiding the forest industry modernization_Liangshui Experimental Forest Farm of Northeast Forestry University (Liangshui National Natural Reserve). In Chinese Forest and Grassland Industry Annuals; China Forestry Publishing House: Beijing, China, 2019; p. 93.
- Xu, L.; Jin, G. Species composition and community structure of a typical mixed broad-leaved-Korean pine (Pinus koraiensis) forest plot in Liangshui Nature Reserve, Northeast China. Biodivers. Sci. 2012, 20, 470–481. [Google Scholar]
- Sun, X.; Tan, B.-X. Estimating forest leaf area index based on CASI remotesens ingdata. Guangdong Agric. Sci. 2012, 39, 189–193. [Google Scholar]
- Zhao, X. Response of Forest Seedlings and Shrubs Regeneration to Topographic Heterogeneity in a Mixed Broadleaved-Korean pine(Pinus Koraiensis) Forest in Xioaxing’an Mountains. Master’s Thesis, Northeast Forestry University, Harbin, China, 2013. [Google Scholar]
- Bao, X.; Liu, B. Avian and mammalian biodiversity of Heilongjiang Liangshui and Fenglin Reserve. Acta Ecol. Sin. 2018, 38, 502–510. [Google Scholar]
- Said, S.; Servanty, S. The influence of landscape structure on female roe deer home-range size. Landsc. Ecol. 2005, 20, 1003–1012. [Google Scholar] [CrossRef]
- Said, S.; Gaillard, J.M.; Duncan, P.; Guillon, N.; Guillon, N.; Servanty, S.; Pellerin, M.; Lefeuvre, K.; Martin, C.; Van Laere, G. Ecological correlates of home-range size in spring-summer for female roe deer (Capreolus capreolus) in a deciduous woodland. J. Zool. 2005, 267, 301–308. [Google Scholar] [CrossRef]
- Said, S.; Gaillard, J.M.; Widmer, O.; Debias, F.; Bourgoin, G.; Delorme, D.; Roux, C. What shapes intra-specific variation in home range size? A case study of female roe deer. Oikos 2009, 118, 1299–1306. [Google Scholar] [CrossRef]
- Morellet, N.; Bonenfant, C.; Borger, L.; Ossi, F.; Cagnacci, F.; Heurich, M.; Kjellander, P.; Linnell, J.D.; Nicoloso, S.; Sustr, P.; et al. Seasonality, weather and climate affect home range size in roe deer across a wide latitudinal gradient within Europe. J. Anim. Ecol. 2013, 82, 1326–1339. [Google Scholar] [CrossRef] [PubMed]
- Dou, H.L.; Yang, H.T.; Feng, L.M.; Mou, P.; Wang, T.M.; Ge, J.P. Estimating the Population Size and Genetic Diversity of Amur Tigers in Northeast China. PLoS ONE 2016, 11, 14. [Google Scholar] [CrossRef]
- Hu, H.; Xing, B.; Yang, M.; Mpemba, H.; Lv, Z.; Zhang, M. Population and genetic diversity of Tibetan red deer based on fecal DNA. J. For. Res. 2018, 29, 227–232. [Google Scholar] [CrossRef]
- Lee, Y.S.; Markov, N.; Voloshina, I.; Argunov, A.; Bayarlkhagva, D.; Oh, J.G.; Park, Y.-S.; Min, M.-S.; Lee, H.; Kim, K.S. Genetic diversity and genetic structure of the Siberian roe deer (Capreolus pygargus) populations from Asia. BMC Genet. 2015, 16, 100. [Google Scholar] [CrossRef]
- Waits, L.P.; Luikart, G.; Taberlet, P. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol. 2001, 10, 249–256. [Google Scholar] [CrossRef]
- Paetkau, D.; Strobeck, C. Microsatellite Analysis of Genetic-Variation in Black Bear Populations. Mol. Ecol. 1994, 3, 489–495. [Google Scholar] [CrossRef]
- Valiere, N. GIMLET: A computer program for analysing genetic individual identification data. Mol. Ecol. Notes 2002, 2, 377–379. [Google Scholar] [CrossRef]
- Taberlet, P.; Luikart, G. Non-invasive genetic sampling and individual identification. Biol. J. Linn. Soc. 1999, 68, 41–55. [Google Scholar] [CrossRef]
- Waits, L.P.; Paetkau, D. Noninvasive genetic sampling tools for wildlife biologists: A review of applications and recommendations for accurate data collection. J. Wildl. Manag. 2005, 69, 1419–1433. [Google Scholar] [CrossRef]
- Perez, T.; Vazquez, F.; Naves, J.; Fernandez, A.; Corao, A.; Albornoz, J.; Dominguez, A. Non-invasive genetic study of the endangered Cantabrian brown bear (Ursus arctos). Conserv. Genet. 2009, 10, 291–301. [Google Scholar] [CrossRef]
- Li, Y.M.; Bai, C.Y.; Niu, W.P.; Yu, H.; Yang, R.J.; Yan, S.Q.; Zhang, J.Y.; Zhang, M.J.; Zhao, Z.H. Isolation and characterization of novel microsatellite markers from the sika deer (Cervus nippon) genome. Genet. Mol. Res. 2015, 14, 11524–11534. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS-pc, Numerical Taxonomy and Multivariate Analysis System, Version 2.11; Exeter Software: Setauket, NY, USA, 2000. [Google Scholar]
- Otis, D.L.; Burnham, K.P.; White, G.C.; Anderson, D.R. Statistical-Inference from Capture Data on Closed Animal Populations. Wildl. Monogr. 1978, 62, 1–135. [Google Scholar]
- White, G.C.; Burnham, K.P. Program MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46, 120–139. [Google Scholar] [CrossRef]
- White, G.C. Closed population estimation models and their extensions in Program MARK. Environ. Ecol. Stat. 2008, 15, 89–99. [Google Scholar] [CrossRef]
- Brinkman, T.J. Resilience of a Deer Hunting System in Southeast Alaska: Integrating Social, Ecological, and Genetic Dimensions. Ph.D. Thesis, University of Alaska Fairbanks, Fairbanks, AK, USA, 2009. [Google Scholar]
- Kendall, K.C.; Stetz, J.B.; Roon, D.A.; Waits, L.P.; Boulanger, J.B.; Paetkau, D. Grizzly Bear Density in Glacier National Park, Montana. J. Wildl. Manag. 2008, 72, 1693–1705. [Google Scholar] [CrossRef]
- Brinkman, T.J.; Schwartz, M.K.; Person, D.K.; Pilgrim, K.L.; Hundertmark, K.J. Effects of time and rainfall on PCR success using DNA extracted from deer fecal pellets. Conserv. Genet. 2010, 11, 1547–1552. [Google Scholar] [CrossRef]
- Koike, S.; Soga, M.; Enari, H.; Kozakai, C.; Nemoto, Y. Seasonal changes and altitudinal variation in deer fecal pellet decay. Eur. J. Wildl. Res. 2013, 59, 765–768. [Google Scholar] [CrossRef]
- Rea, R.V.; Johnson, C.J.; Murray, B.W.; Hodder, D.P.; Crowley, S.M. Timing Moose Pellet Collections to Increase Genotyping Success of Fecal DNA. J. Fish Wildl. Manag. 2016, 7, 461–466. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M. Population Genetic Diversity of Roe Deer (Capreolus pygargus) in Mountains of Heilongjiang Province. Zool. Res. 2009, 30, 113–120. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, G.; Li, H. A comparative study on four survey methods used in ungulate population size estimation in winter in North China. Acta Ecol. Sin. 2015, 35, 3076–3086. [Google Scholar]
- Gilbert, B.A.; Raedeke, K.J.; Skalski, J.R.; Stringer, A.B. Modeling black-tailed deer population dynamics using structured and unstructured approaches. J. Wildl. Manag. 2007, 71, 144–154. [Google Scholar] [CrossRef]
- Melis, C.; Jędrzejewska, B.; Apollonio, M.; Bartoń, K.A.; Jędrzejewski, W.; Linnell, J.D.; Kojola, I.; Kusak, J.; Adamic, M.; Ciuti, S. Predation has a greater impact in less productive environments: Variation in roe deer, Capreolus capreolus, population density across Europe. Glob. Ecol. Biogeogr. 2009, 18, 724–734. [Google Scholar] [CrossRef]
- Wilson, K.R.; Anderson, D.R. Evaluation of Two Density Estimators of Small Mammal Population Size. J. Mammal. 1985, 66, 13–21. [Google Scholar] [CrossRef]
- Parmenter, R.R.; Yates, T.L.; Anderson, D.R.; Burnham, K.P.; Dunnum, J.L.; Franklin, A.B.; Friggens, M.T.; Lubow, B.C.; Miller, M.; Olson, G.S. Small-mammal density estimation: A field comparison of grid-based vs. web-based density estimators. Ecol. Monogr. 2003, 73, 1–26. [Google Scholar] [CrossRef]
- Ivan, J.S.; White, G.C.; Shenk, T.M. Using simulation to compare methods for estimating density from capture–recapture data. Ecology 2013, 94, 817–826. [Google Scholar] [CrossRef]
- Li, X. Research on the Patterns of Management System Reform in Daxing’anling and Xiaoxing’anling State-owned Forest Region. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Fickel, J.; Reinsch, A. Microsatellite markers for the European Roe deer (Capreolus capreolus). Mol. Ecol. 2000, 9, 994–995. [Google Scholar] [CrossRef]
- Galan, M.; Cosson, J.F.; Aulagnier, S.; Maillard, J.-C.; Thevenon, S.; Hewison, A. Cross-amplification tests of ungulate primers in roe deer (Capreolus capreolus) to develop a multiplex panel of 12 microsatellite loci. Mol. Ecol. Notes 2003, 3, 142–146. [Google Scholar] [CrossRef]
- Vial, L.; Maudet, C.; Luikart, G. Thirty-four polymorphic microsatellites for European roe deer. Mol. Ecol. Resour. 2003, 3, 523–527. [Google Scholar] [CrossRef]


| Locus | A | AE | HE | HO | PIC | PID | PIDsibs |
|---|---|---|---|---|---|---|---|
| Roe1 | 5 | 5 | 0.192 | 0.183 | 0.175 | 7.785 × 10−7 | 8.276 × 10−3 |
| Roe8 | 16 | 4 | 0.909 | 0.879 | 0.866 | 3.790 × 10−4 | 9.560 × 10−1 |
| Roe9 | 4 | 3 | 0.483 | 0.551 | 0.462 | 3.140 × 10−6 | 1.693 × 10−2 |
| BM757 | 17 | 7 | 0.796 | 0.908 | 0.899 | 1.474 × 10−2 | 3.008 × 10−1 |
| MB25 | 3 | 3 | 0.498 | 0.505 | 0.381 | 1.157 × 10−6 | 9.998 × 10−3 |
| OarFCB304 | 19 | 6 | 0.868 | 0.870 | 0.855 | 1.088 × 10−5 | 3.092 × 10−2 |
| Mean | 10.7 | 4.167 | 0.624 | 0.649 | 0.606 | -- | -- |
| Date | No. of Pellet Groups Collected | Successfully Genotyped | Success Rate in Genotyping |
|---|---|---|---|
| December 2016 | 165 | 130 | 78.8% |
| March 2016 | 257 | 135 | 52.5% |
| Model (Fitting Value) | Roe Deer Abundance (Within 95% CI) (Deer) | SE |
|---|---|---|
| Mtbh (1.00) | 87 (80–112) | 7 |
| Mo (0.73) | 201 (134–350) | 52 |
| Mt (0.33) | 182 (125–311) | 44 |
| Mtb (0.11) | 78 (78–92) | 23 |
| Mt- Chao | 176 (122–295) | 42 |
| Mb- Chao | 334 (195–640) | 107 |
| Mh (0.00) | 110 (99–127) | 7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, N.; Chen, L.; Li, Y.; Xiong, Z.; Hu, Y. Estimating Abundance of Siberian Roe Deer Using Fecal-DNA Capture-Mark-Recapture in Northeast China. Animals 2020, 10, 1135. https://doi.org/10.3390/ani10071135
Li Y, Li N, Chen L, Li Y, Xiong Z, Hu Y. Estimating Abundance of Siberian Roe Deer Using Fecal-DNA Capture-Mark-Recapture in Northeast China. Animals. 2020; 10(7):1135. https://doi.org/10.3390/ani10071135
Chicago/Turabian StyleLi, Yuehui, Nana Li, Long Chen, Yueyuan Li, Zaiping Xiong, and Yuanman Hu. 2020. "Estimating Abundance of Siberian Roe Deer Using Fecal-DNA Capture-Mark-Recapture in Northeast China" Animals 10, no. 7: 1135. https://doi.org/10.3390/ani10071135
APA StyleLi, Y., Li, N., Chen, L., Li, Y., Xiong, Z., & Hu, Y. (2020). Estimating Abundance of Siberian Roe Deer Using Fecal-DNA Capture-Mark-Recapture in Northeast China. Animals, 10(7), 1135. https://doi.org/10.3390/ani10071135