Optimal Dietary Protein Requirement for Juvenile Sesarmid Crab (Episesarma singaporense)



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Survival, Growth Performance and Feed Utilization
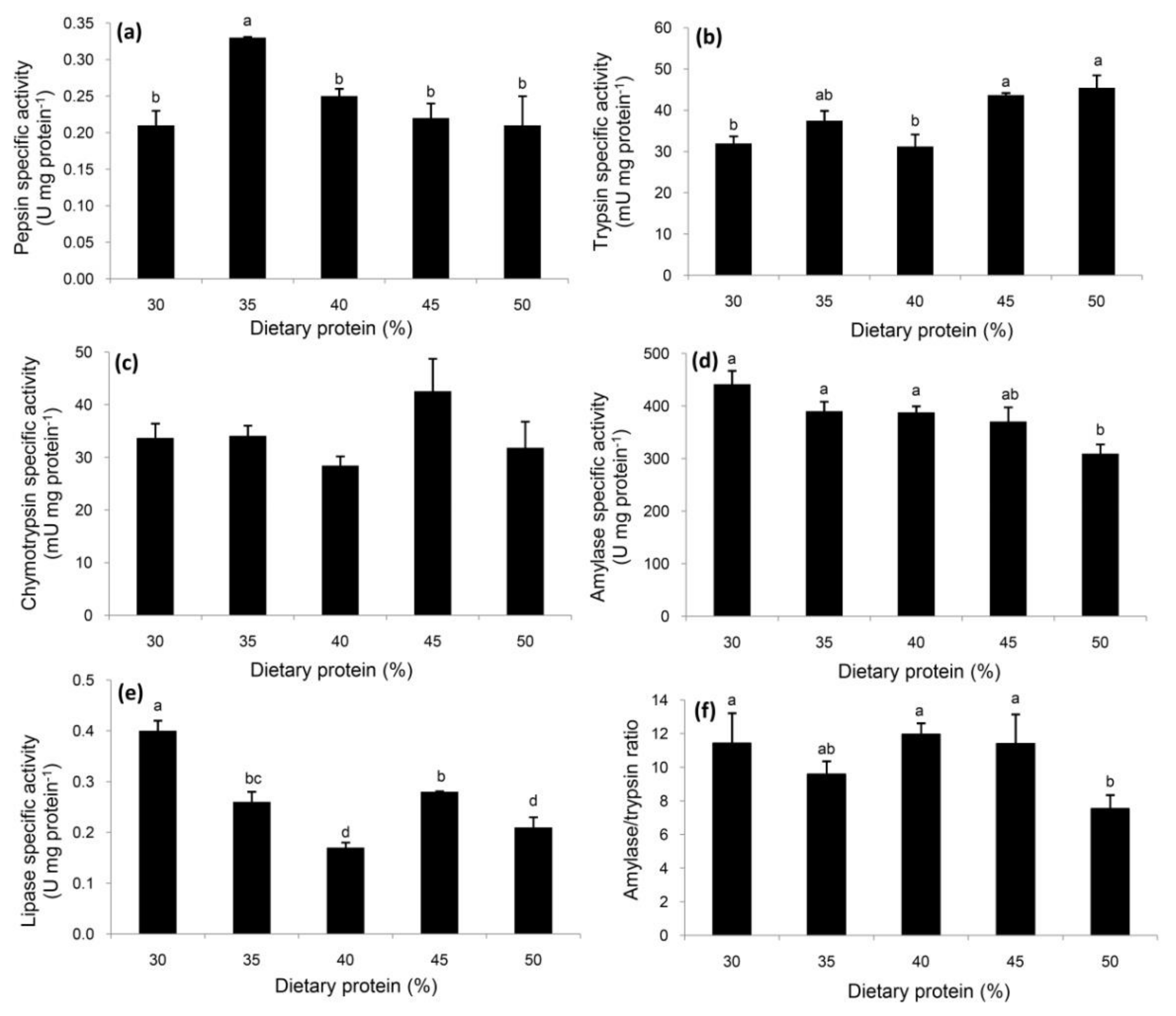
3.2. Specific Activities of Digestive Enzymes
3.3. Muscle Amino Acid Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, B.Y.; Ng, N.K.; Ng, P.K.L. The taxonomy of five species of Episesarma De Man, 1895, in Singapore (Crustacea: Decapoda: Brachyura: Sesarmidae). Raffles Bull. Zool. 2015, 31, 199–215. [Google Scholar]
- Ng, P.K.L. Crabs. In FAO Species Identification Guide for Fishery Purposes: The Living Marine Resources of the Western Central Pacific Volume 2. Cephalopods, Crustaceans, Holothurians and Sharks; Carpenter, K.E., Niem, V.H., Eds.; FAO: Rome, Italy, 1998; pp. 1143–1146. [Google Scholar]
- Tiensongrassamee, B. Vinegar Crabs; The Thailand Research Fund: Bangkok, Thailand, 2009; pp. 20–21. (In Thai) [Google Scholar]
- Carter, C.G.; Mente, E. Protein synthesis in crustaceans: A review focused on feeding and nutrition. Cent. Eur. J. Biol. 2014, 9, 1–10. [Google Scholar] [CrossRef]
- Glencross, B.D.; Booth, M.; Allan, G.L. A feed is only as good as its ingredients—A review of ingredient evaluation strategies for aquaculture feeds. Aquac. Nutr. 2007, 13, 17–34. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011; pp. 59–60. [Google Scholar]
- Wilson, R.P. Amino Acid and Proteins. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: London, UK, 2002; pp. 144–174. [Google Scholar]
- Castine, S.; Southgate, P.C.; Zeng, C. Evaluation of four dietary protein sources for use in microbound diets fed to megalopae of the blue swimmer crab, Portunus pelagicus. Aquaculture 2008, 281, 95–99. [Google Scholar] [CrossRef]
- Mu, Y.Y.; Shim, K.F.; Guo, J.Y. Effects of protein level in isocaloric diets on growth performance of the juvenile Chinese hairy crab, Eriocheir sinensis. Aquaculture 1998, 165, 139–148. [Google Scholar] [CrossRef]
- Jin, M.; Zhou, Q.; Zhang, W.; Xie, F.; Shen, T.J.; Huang, X. Dietary protein requirements of the juvenile swimming crab, Portunus trituberculatus. Aquaculture 2013, 414–415, 303–308. [Google Scholar] [CrossRef]
- Holme, M.; Zeng, C.; Southgate, P.C. A review of recent progress toward development of a formulated microbound diet for mud crab, Scylla serrata, larvae and their nutritional requirements. Aquaculture 2009, 286, 164–175. [Google Scholar] [CrossRef]
- Wang, J.; Qi, Z.; Yang, Z. Evaluation of the protein requirement of juvenile Chinese soft-shelled turtle (Pelodiscus sinensis, Wiegmann) fed with practical diets. Aquaculture 2014, 433, 252–255. [Google Scholar] [CrossRef]
- Zhou, J.B.; Zhou, Q.C.; Chi, S.Y.; Yang, Q.H.; Liu, C.W. Optimal dietary protein requirement for juvenile ivory shell, Babylonia areolate. Aquaculture 2007, 270, 186–192. [Google Scholar] [CrossRef]
- Perera, E.; Fraga, I.; Carrillo, O.; Diaz-Iglesias, E.; Cruz, R.; Baez, M.; Galich, G.S. Evaluation of practical diets for the Caribbean spiny lobster Panulirus argus (Latreille, 1804): Effects of protein sources on substrate metabolism and digestive proteases. Aquaculture 2005, 244, 251–262. [Google Scholar] [CrossRef]
- Sheen, S.S. Dietary cholesterol requirements of juvenile mud crab Scylla serrata. Aquaculture 2000, 189, 277–285. [Google Scholar] [CrossRef]
- Thongrod, S.; Boonyaratpulin, M. Cholesterol and lecithin requirement of juvenile banana shrimp, Penaeus merguiensis. Aquaculture 1988, 161, 315–321. [Google Scholar] [CrossRef]
- Unnikrishnan, U.; Paulraj, R. Dietary protein requirement of giant mud crab Scylla serrata juveniles fed iso-energetic formulated diets having graded protein levels. Aquac. Res. 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Holme, M.; Zeng, C.; Southgate, P.C. Use of microbound diets for larval culture of the mud crab, Scylla serrata. Aquaculture 2006, 257, 482–490. [Google Scholar] [CrossRef]
- AOAC. Official Method of Analysis of AOAC International; Association of Official Analytical Chemists: Arlington, VA, USA, 1990; pp. 69–84. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fisheries Research Board of Canada Bulletin: Ottawa, ON, Canada, 1972; pp. 71–80. [Google Scholar]
- Sudtongkong, C.; Thongprajukaew, K.; Hahor, W.; Saekhow, S. Ontogenetic development of digestive enzymes and elemental composition of sesarmid crab Episesarma singaporense. Fish. Sci. 2019, 85, 991–1000. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Andrés, M.; Gisbert, E.; Díaz, M.; Moyano, F.J.; Estévez, A.; Rotllant, G. Ontogenetic changes in digestive enzymatic capacities of the spider crab, Maja brachydactyla (Decapoda: Majidae). J. Exp. Mar. Biol. Ecol. 2010, 389, 75–84. [Google Scholar] [CrossRef]
- Chamchuen, P.; Pratoomchat, B.; Engkakul, A.; Kovitvadhi, U.; Rungruangsak-Torrissen, K. Development of enzymes and in vitro digestibility during metamorphosis and molting of blue swimming crab (Portunus pelagicus). J. Mar. Biol. 2014. [Google Scholar] [CrossRef]
- Cherif, S.; Gargouri, Y. Thermoactivity and effects of organic solvents on digestive lipase from hepatopancreas of the green crab. Food Chem. 2009, 116, 82–86. [Google Scholar] [CrossRef]
- Worthington. Worthington Enzyme Manual. Enzymes and Related Biochemicals; Worthington Chemical: New Jersey, NJ, USA, 1993; p. 399. [Google Scholar]
- Rungruangsak-Torrissen, K.; Moss, R.; Andresen, L.H.; Berg, A.; Waagbo, R. Different expressions of trypsin and chymotrypsin in relation to growth in Atlantic salmon (Salmo salar L.). Fish Physiol. Biochem. 2006, 32, 7–23. [Google Scholar] [CrossRef]
- Areekijseree, M.; Engkagul, A.; Kovitvadhi, U.; Thongpan, A.; Mingmuang, M.; Pakkong, P.; Rungruangsak-Torrissen, K. Temperature and pH characteristics of amylase and proteinase of adult freshwater pearl mussel, Hyriopsis (Hyriopsis) bialatus Simpson 1900. Aquaculture 2004, 234, 575–587. [Google Scholar] [CrossRef]
- Winkler, U.K.; Stuckmann, M. Glycogen, hyaluronate and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J. Bacteriol. 1979, 138, 663–670. [Google Scholar] [CrossRef] [PubMed]
- González-Félix, M.L.; Gómez-Jiménez, S.; Perez-Velazquez, M.; Davis, D.A.; Velazco-Rameños, J.G. Nitrogen budget for a low salinity, zero-water exchange culture system: I. Effect of dietary protein level on the performance of Litopenaeus vannamei (Boone). Aquac. Nutr. 2007, 38, 798–808. [Google Scholar] [CrossRef]
- Huo, Y.W.; Jin, M.; Zhou, P.P.; Li, M.; Mai, K.S.; Zhou, Q.C. Effects of dietary protein and lipid levels on growth, feed utilization and body composition of juvenile swimming crab, Portunus trituberculatus. Aquaculture 2014, 434, 151–158. [Google Scholar] [CrossRef]
- Jones, D.A.; Kamarudin, M.S.; LeVay, L.L. The potential for the replacement of live feeds in larval culture. J. World Aquac. Soc. 1993, 24, 199–210. [Google Scholar] [CrossRef]
- Lee, D.J. Studies on protein utilization related to growth in Penaeus monodon. Agriculture 1971, 1, 1–13. [Google Scholar]
- Briggs, M.; Jauncey, K.; Brown, J. The cholesterol and lecithin requirement of juvenile prawn (Macrobrachium rosenbergii) fed semi-purified diets. Aquaculture 1988, 70, 121–129. [Google Scholar] [CrossRef]
- Celis-Guerrero, L.E.; Garcia-Carreno, F.L.; Navarrete del Toro, A.N. Characterization of proteases in the digestive system of spiny lobster (Panulirus interruptus). Mar. Biotechnol. 2004, 6, 262–269. [Google Scholar] [CrossRef]
- Navarrete del Toro, M.; Garcia-Carreño, F.; Lopez, M.D.; Celis-Guerrero, L.; Saborowski, R. Aspartic proteinases in the digestive tract of marine decapod crustaceans. J. Exp. Zool. 2006, 305A, 645–654. [Google Scholar] [CrossRef]
- Khoa, T.N.D.; Mai, N.T.; Linh, N.K.; Mi, L.T.Y.; Shaharom-Harrison, F. Ontogenic development of digestive enzymes of mud crab (Scylla paramamosain) during larval stages. Thalass. Int. J. Mar. Sci. 2019, 35, 655–661. [Google Scholar] [CrossRef]
- Dunn, B.M. Overview of pepsin-like aspartic peptidases. Curr. Protoc. Protein Sci. 2001, 25, 21.3.1–21.3.6. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Alarcón, D.; Saborowski, R.; Rojo-Arreola, L.; García-Carreño, F. Is digestive cathepsin D the rule in decapod crustaceans? Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 215, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Rotllant, G.; Guerao, G.; Sastre, M.; Anger, K. Developmental and moult cycle related biochemical changes in larvae of the spider crab, Maja brachydactyla (Brachyura: Majidae). Sci. Mar. 2012, 76, 483–488. [Google Scholar] [CrossRef]
- Péres, A.; Zambonino-Infante, J.L.; Cahu, C. Dietary regulation of activities and mRNA levels of trypsin and amylase in sea bass (Dicentrarchus labrax) larvae. Fish Physiol. Biochem. 1998, 19, 145–152. [Google Scholar] [CrossRef]
- Krogdahl, A.; Bakke-McKellep, A.M.; Baeverfjord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Hofer, R.; Schiemer, F. Proteolytic activity in the digestive tract of several species of fish with different feeding habits. Oecologia 1981, 48, 342–345. [Google Scholar] [CrossRef]
- Johnston, D. Ontogenetic changes in digestive enzyme activity of the spiny lobster, Jasus edwardsii (Decapoda; Palinuridae). Mar. Biol. 2003, 143, 1071–1082. [Google Scholar] [CrossRef]
- Perera, E.; Moyano, F.J.; Díaz, M.; Perdomo-Morales, R.; Montero-Alejo, V.; Rodriguez-Viera, L.; Alonso, E.; Carrillo, O.; Galich, G.S. Changes in digestive enzymes through developmental and molt stages in the spiny lobster, Panulirus argus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 250–256. [Google Scholar] [CrossRef]
- Hewitt, D.R. Response of protein turnover in the brown tiger prawn Penaeus esculentus to variation in dietary protein content. Comp. Biochem. Physiol. Part A Physiol. 1992, 103, 183–187. [Google Scholar] [CrossRef]
- Andrés, M.; Estévez, A.; Hontoria, F.; Rotllant, G. Differential utilization of biochemical components during larval development of the spider crab Maja brachydactyla (Decapoda: Majidae). Mar. Biol. 2010, 157, 2329–2340. [Google Scholar] [CrossRef]
- Akiyama, T.; Oohara, I.; Yamamoto, T. Comparison of essential amino acid requirements with A/E ratio among fish species. Fish. Sci. 1997, 63, 963–970. [Google Scholar] [CrossRef]
- Catacutan, M.R. Growth and body composition of juvenile mud crab, Scylla serrata, fed different dietary protein and lipid levels and protein to energy ratios. Aquaculture 2002, 208, 113–123. [Google Scholar] [CrossRef]
- Guzman, C.; Gaxiola, G.; Rosa, C.; Torre-Blanco, A. The effect of dietary protein and total energy content on digestive enzyme activities, growth and survival of Litopenaeus setiferus (Linnaeus 1767) postlarvae. Aquac. Nutr. 2001, 7, 113–122. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient and Composition | Experimental Diet (% CP) | ||||
|---|---|---|---|---|---|
| 30 | 35 | 40 | 45 | 50 | |
| Ingredient | |||||
| Fish meal | 16.75 | 21.76 | 26.76 | 31.76 | 36.76 |
| Shrimp meal | 5 | 5 | 5 | 5 | 5 |
| Soybean meal | 20 | 20 | 20 | 20 | 20 |
| Wheat gluten meal | 12.09 | 14.92 | 17.77 | 20.61 | 23.46 |
| Fish oil | 2.56 | 2.33 | 2.11 | 1.88 | 1.66 |
| Soybean oil | 2.56 | 2.33 | 2.11 | 1.88 | 1.66 |
| Dextrin | 36.24 | 28.08 | 19.89 | 11.74 | 3.54 |
| Soy lecithin | 1 | 1 | 1 | 1 | 1 |
| Vitamin premix a | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| Mineral premix b | 3 | 3 | 3 | 3 | 3 |
| Cellulose | 0 | 0.78 | 1.56 | 2.33 | 3.12 |
| Composition | |||||
| Moisture | 10.47 ± 0.21 | 10.06 ± 0.10 | 12.61 ± 0.13 | 8.62 ± 0.20 | 10.06 ± 0.19 |
| CP | 30.71 ± 0.19 | 35.84 ± 0.21 | 38.64 ± 0.24 | 46.34 ± 0.23 | 50.49 ± 0.19 |
| Crude lipid | 6.95 ± 0.18 | 6.81 ± 0.25 | 6.41 ± 0.19 | 7.49 ± 0.17 | 7.12 ± 0.20 |
| Ash | 8.00 ± 0.11 | 9.74 ± 0.21 | 10.76 ± 0.18 | 12.71 ± 0.12 | 13.99 ± 0.17 |
| Crude fiber | 2.20 ± 0.12 | 2.00 ± 0.15 | 2.09 ± 0.13 | 2.11 ± 0.17 | 2.66 ± 0.10 |
| NFE | 41.67 ± 0.15 | 35.55 ± 0.56 | 29.49 ± 0.37 | 22.73 ± 0.41 | 15.68 ± 0.16 |
| GE (kcal kg−1) | 4175 ± 10 | 4189 ± 13 | 4064 ± 13 | 4321 ± 5 | 4252 ± 28 |
| Amino Acid | Experimental Diet (% CP) | A/E Ratio | ||||
|---|---|---|---|---|---|---|
| 30 | 35 | 40 | 45 | 50 | ||
| EAA | ||||||
| Arginine | 5.75 ± 0.02 | 6.25 ± 0.08 | 4.78 ± 0.06 | 5.84 ± 0.09 | 4.20 ± 0.04 | 0.20 |
| Histidine | 0.79 ± 0.07 | 0.96 ± 0.09 | 0.76 ± 0.05 | 1.08 ± 0.04 | 0.84 ± 0.06 | 0.03 |
| Isoleucine | 1.44 ± 0.06 | 1.52 ± 0.08 | 1.27 ± 0.03 | 1.98 ± 0.04 | 1.46 ± 0.06 | 0.06 |
| Leucine | 2.77 ± 0.02 | 3.07 ± 0.04 | 2.74 ± 0.08 | 3.42 ± 0.10 | 3.24 ± 0.10 | 0.12 |
| Lysine | 4.77 ± 0.04 | 4.83 ± 0.09 | 4.02 ± 0.10 | 5.20 ± 0.12 | 4.29 ± 0.08 | 0.18 |
| Methionine | 5.57 ± 0.04 | 5.61 ± 0.02 | 4.90 ± 0.12 | 5.41 ± 0.05 | 4.41 ± 0.09 | 0.20 |
| Phenylalanine | 1.34 ± 0.08 | 1.56 ± 0.07 | 1.34 ± 0.09 | 1.67 ± 0.05 | 1.68 ± 0.12 | 0.06 |
| Threonine | 1.97 ± 0.09 | 2.09 ± 0.07 | 1.59 ± 0.06 | 2.25 ± 0.07 | 1.69 ± 0.06 | 0.07 |
| Tryptophan | 0.60 ± 0.02 | 0.63 ± 0.02 | 0.60 ± 0.02 | 0.40 ± 0.01 | 0.77 ± 0.02 | 0.02 |
| Valine | 1.65 ± 0.04 | 1.65 ± 0.07 | 1.46 ± 0.03 | 2.19 ± 0.05 | 1.73 ± 0.04 | 0.07 |
| ΣEAA | 26.65 ± 0.17 | 28.17 ± 0.13 | 23.46 ± 0.14 | 29.44 ± 0.09 | 24.3 ± 0.12 | |
| NEAA | ||||||
| Alanine | 2.57 ± 0.06 | 2.28 ± 0.09 | 2.28 ± 0.04 | 2.96 ± 0.06 | 2.83 ± 0.08 | |
| Aspartic acid | 2.20 ± 0.09 | 2.15 ± 0.06 | 1.54 ± 0.10 | 2.66 ± 0.12 | 1.52 ± 0.12 | |
| Asparagine | nd | nd | nd | nd | nd | |
| Cysteine | nd | nd | nd | nd | nd | |
| Cystine | 0.03 ± 0.01 | 0.41 ± 0.02 | 0.13 ± 0.01 | 0.31 ± 0.02 | 0.27 ± 0.02 | |
| Glutamic acid | 4.85 ± 0.08 | 4.50 ± 0.09 | 3.84 ± 0.07 | 5.42 ± 0.10 | 3.07 ± 0.11 | |
| Glutamine | 0.98 ± 0.07 | 1.41 ± 0.06 | 0.80 ± 0.06 | 1.23 ± 0.10 | 1.49 ± 0.05 | |
| Glycine | 2.80 ± 0.08 | 2.85 ± 0.07 | 2.55 ± 0.08 | 3.50 ± 0.09 | 3.05 ± 0.09 | |
| Proline | 4.11 ± 0.03 | 4.50 ± 0.02 | 3.72 ± 0.04 | 5.52 ± 0.02 | 3.95 ± 0.03 | |
| Hydroxyproline | 0.33 ± 0.05 | 0.29 ± 0.04 | 0.23 ± 0.02 | 0.46 ± 0.03 | 0.30 ± 0.01 | |
| Serine | 2.25 ± 0.07 | 2.21 ± 0.06 | 1.69 ± 0.04 | 2.45 ± 0.09 | 1.80 ± 0.07 | |
| Tyrosine | 0.57 ± 0.01 | 1.00 ± 0.03 | 0.69 ± 0.02 | 1.02 ± 0.03 | 0.90 ± 0.03 | |
| ΣNEAA | 20.69 ± 0.14 | 21.6 ± 0.21 | 17.47 ± 0.17 | 25.53 ± 0.27 | 19.18 ± 0.25 | |
| ΣEAA/ΣNEAA | 1.29 ± 0.01 | 1.31 ± 0.02 | 1.34 ± 0.02 | 1.15 ± 0.01 | 1.27 ± 0.01 | |
| Enzyme | Condition * | Substrate ** | Product | Absorbance |
|---|---|---|---|---|
| Pepsin-like | pH 2 at 37 °C [23] | Hemoglobin | - | 280 nm |
| Trypsin | pH 9 at 60 °C [24] | BAPNA | p-Nitroanilide | 410 nm |
| Chymotrypsin | pH 9 at 40 °C [24] | SAPNA | p-Nitroanilide | 410 nm |
| Amylase | pH 7 at 50 °C [24] | Starch | Maltose | 540 nm |
| Lipase | pH 8 at 60 °C [25] | p-NPP | p-Nitrophenol | 410 nm |
| Parameter | Experimental Diet (% CP) | ||||
|---|---|---|---|---|---|
| 30 | 35 | 40 | 45 | 50 | |
| Survival (%) | 34.43 ± 14.90 b | 75.03 ± 5.91 a | 78.12 ± 10.87 a | 87.5 ± 3.34 a | 78.13 ± 6.95 a |
| WG (%) | 29.58 ± 10.22 | 31.15 ± 10.20 | 32.42 ± 9.04 | 46.67 ± 7.70 | 33.33 ± 8.33 |
| CWG (%) | 10.05 ± 2.94 | 12.69 ± 4.83 | 10.49 ± 3.19 | 21.74 ± 4.06 | 11.69 ± 3.70 |
| SGR (% BW day−1) | 0.41 ± 0.14 | 0.43 ± 0.10 | 0.47 ± 0.21 | 0.68 ± 0.09 | 0.36 ± 0.12 |
| FCR (g feed g gain−1) | 1.94 ± 0.33 | 1.67 ± 0.31 | 1.66 ± 0.32 | 1.17 ± 0.06 | 2.05 ± 0.53 |
| PER (g gain g protein−1) | 0.17 ± 0.03 b | 0.23 ± 0.08 b | 0.26 ± 0.06 b | 0.54 ± 0.09 a | 0.29 ± 0.04 b |
| Amino Acid | Experimental Diet (% CP) | A/E Ratio | ||||
|---|---|---|---|---|---|---|
| 30 | 35 | 40 | 45 | 50 | ||
| EAA | ||||||
| Arginine | 1.41 ± 0.01 c | 1.79 ± 0.01 b | 1.91 ± 0.01 a | 1.92 ± 0.01 a | 1.34 ± 0.01 d | 0.24 |
| Histidine | 0.24 ± 0.01 c | 0.26 ± 0.01 d | 0.35 ± 0.01 b | 0.37 ± 0.01 a | 0.28 ± 0.01 c | 0.04 |
| Isoleucine | 0.38 ± 0.01 e | 0.48 ± 0.01 c | 0.56 ± 0.01 a | 0.55 ± 0.01 b | 0.44 ± 0.01 d | 0.07 |
| Leucine | 0.68 ± 0.01 d | 0.83 ± 0.01 c | 1.07 ± 0.01 a | 1.04 ± 0.01 b | 0.83 ± 0.01 c | 0.13 |
| Lysine | 1.11 ± 0.01 d | 1.30 ± 0.01 c | 1.80 ± 0.01 a | 1.81 ± 0.01 a | 1.59 ± 0.01 b | 0.22 |
| Methionine | 0.35 ± 0.01 d | 0.39 ± 0.01 b | 0.36 ± 0.01 c | 0.47 ± 0.01 a | 0.36 ± 0.01 c | 0.06 |
| Phenylalanine | 0.40 ± 0.01 e | 0.46 ± 0.01 d | 0.60 ± 0.01 b | 0.64 ± 0.01 a | 0.49 ± 0.01 c | 0.08 |
| Threonine | 0.36 ± 0.01 d | 0.45 ± 0.01 b | 0.55 ± 0.01 a | 0.55 ± 0.01 a | 0.43 ± 0.01 c | 0.07 |
| Tryptophan | 0.15 ± 0.01 d | 0.17 ± 0.01 c | 0.19 ± 0.01 a | 0.19 ± 0.01 a | 0.18 ± 0.01 b | 0.03 |
| Valine | 0.39 ± 0.01 d | 0.50 ± 0.01 b | 0.58 ± 0.01 a | 0.49 ± 0.01 c | 0.39 ± 0.01 d | 0.07 |
| ΣEAA | 5.48 ± 0.01 e | 6.62 ± 0.01 c | 7.97 ± 0.02 b | 8.03 ± 0.01 a | 6.33 ± 0.01 d | |
| NEAA | ||||||
| Alanine | 0.57 ± 0.01 d | 0.75 ± 0.01 b | 0.86 ± 0.01 a | 0.62 ± 0.01 c | 0.55 ± 0.01 e | |
| Aspartic acid | 1.01 ± 0.01 e | 1.51 ± 0.01 c | 1.74 ± 0.01 a | 1.70 ± 0.01 b | 1.43 ± 0.01 d | |
| Cysteine | 0.03 ± 0.01 | 0.02 ± 0.01 | nd | nd | nd | |
| Cystine | 0.08 ± 0.01 d | 0.10 ± 0.01 c | 0.11 ± 0.01 b | 0.15 ± 0.01 a | 0.11 ± 0.01 b | |
| Glutamic acid | 1.11 ± 0.01 e | 1.56 ± 0.01 d | 1.77 ± 0.01 b | 1.86 ± 0.10 a | 1.59 ± 0.01 c | |
| Glutamine | 1.17 ± 0.01 b | 0.91 ± 0.01 c | 1.47 ± 0.01 a | 1.19 ± 0.01 b | 0.99 ± 0.01 c | |
| Glycine | 0.99 ± 0.01 e | 1.29 ± 0.01 c | 1.60 ± 0.01 a | 1.36 ± 0.01 b | 1.09 ± 0.01 d | |
| Proline | 0.65 ± 0.01 c | 0.65 ± 0.01 c | 0.91 ± 0.01 a | 0.71 ± 0.01 b | 0.55 ± 0.01 d | |
| Serine | 0.32 ± 0.01 e | 0.42 ± 0.01 c | 0.52 ± 0.01 a | 0.48 ± 0.01 b | 0.38 ± 0.01 d | |
| Tyrosine | 0.39 ± 0.01 d | 0.44 ± 0.01 c | 0.53 ± 0.01 b | 0.60 ± 0.01 a | 0.45 ± 0.01 c | |
| ΣNEAA | 6.33 ± 0.01 e | 7.66 ± 0.01 c | 9.52 ± 0.01 a | 8.68 ± 0.10 b | 7.13 ± 0.01 d | |
| ΣEAA/ΣNEAA | 0.87 ± 0.01 c | 0.86 ± 0.01 c | 0.84 ± 0.01 d | 0.92 ± 0.01 a | 0.89 ± 0.01 b | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudtongkong, C.; Thongprajukaew, K.; Saekhow, S. Optimal Dietary Protein Requirement for Juvenile Sesarmid Crab (Episesarma singaporense). Animals 2020, 10, 998. https://doi.org/10.3390/ani10060998
Sudtongkong C, Thongprajukaew K, Saekhow S. Optimal Dietary Protein Requirement for Juvenile Sesarmid Crab (Episesarma singaporense). Animals. 2020; 10(6):998. https://doi.org/10.3390/ani10060998
Chicago/Turabian StyleSudtongkong, Chanyut, Karun Thongprajukaew, and Suktianchai Saekhow. 2020. "Optimal Dietary Protein Requirement for Juvenile Sesarmid Crab (Episesarma singaporense)" Animals 10, no. 6: 998. https://doi.org/10.3390/ani10060998
APA StyleSudtongkong, C., Thongprajukaew, K., & Saekhow, S. (2020). Optimal Dietary Protein Requirement for Juvenile Sesarmid Crab (Episesarma singaporense). Animals, 10(6), 998. https://doi.org/10.3390/ani10060998