Dietary Supplementation of Postbiotics Mitigates Adverse Impacts of Heat Stress on Antioxidant Enzyme Activity, Total Antioxidant, Lipid Peroxidation, Physiological Stress Indicators, Lipid Profile and Meat Quality in Broilers

 , , , , and
, , , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Postbiotics Production
2.2. Ethical Note, Birds, Experimental Design and Housing
2.3. Samples Collection
2.4. Plasma Antioxidant Enzymes Biomarkers
2.4.1. Total Antioxidant Capacity
2.4.2. Superoxide Dismutase Activity
2.4.3. Catalase Activity
2.4.4. Glutathione Peroxidase Activity
2.4.5. Glutathione Activity
2.5. Plasma Lipid Profile
2.6. Lipid Peroxidation
2.7. Plasma Corticosterone
2.8. Heat Shock Protein 70 (HSP70)
2.9. Plasma Acute Phase Proteins
2.9.1. α1-Acid Glycoprotein (α1-AGP)
2.9.2. Ceruloplasmin (CPN)
2.10. Meat Quality
2.10.1. Drip Loss
2.10.2. Cooking Loss
2.10.3. Shear Force (Tenderness)
2.10.4. Colour
2.10.5. Meat pH
2.11. Statistical Analysis
3. Results
3.1. Antioxidant Enzyme Activities
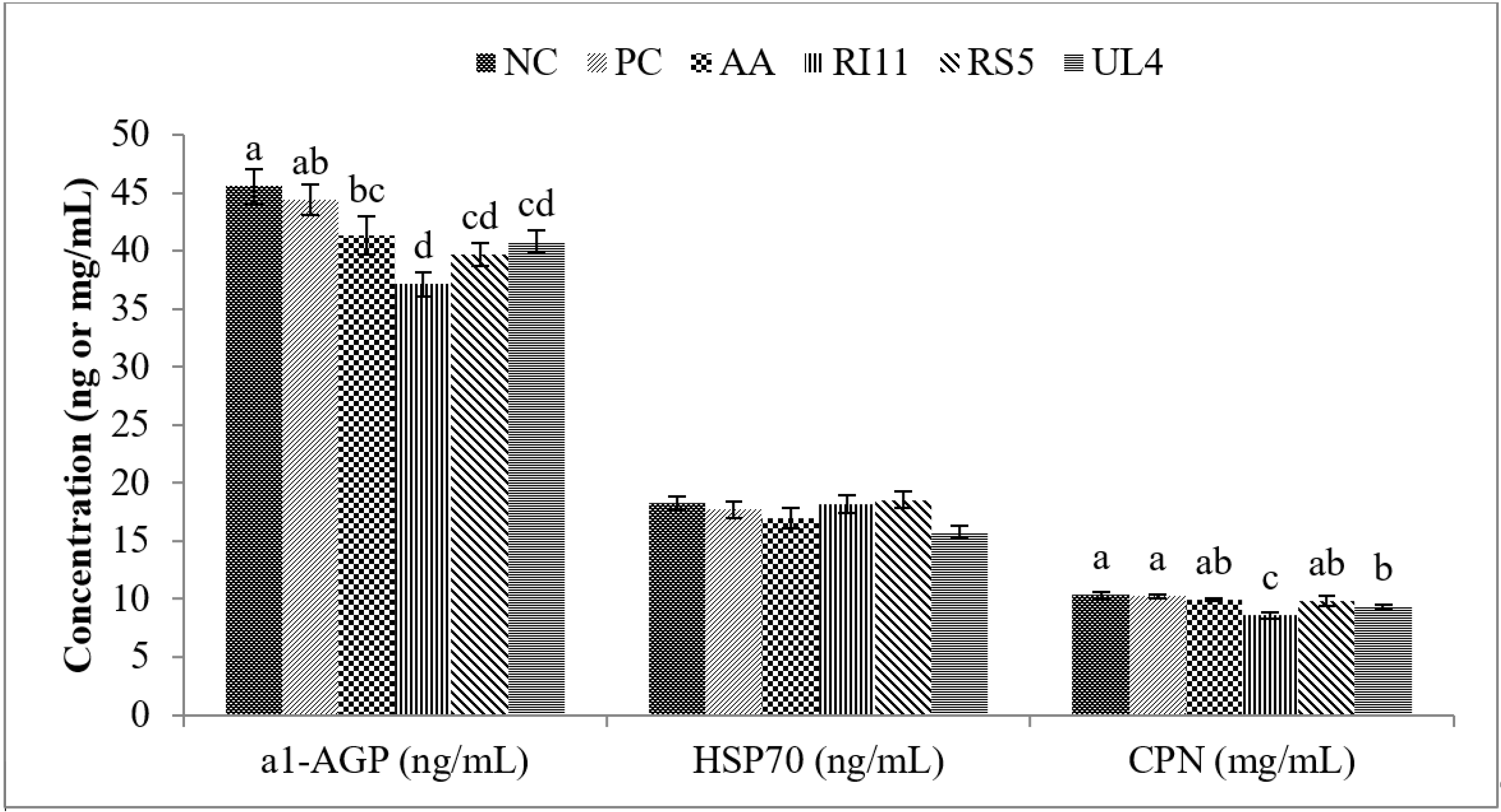
3.2. Acute Phase Proteins (APPs) and Heat Shock Protein 70 (HSP70)
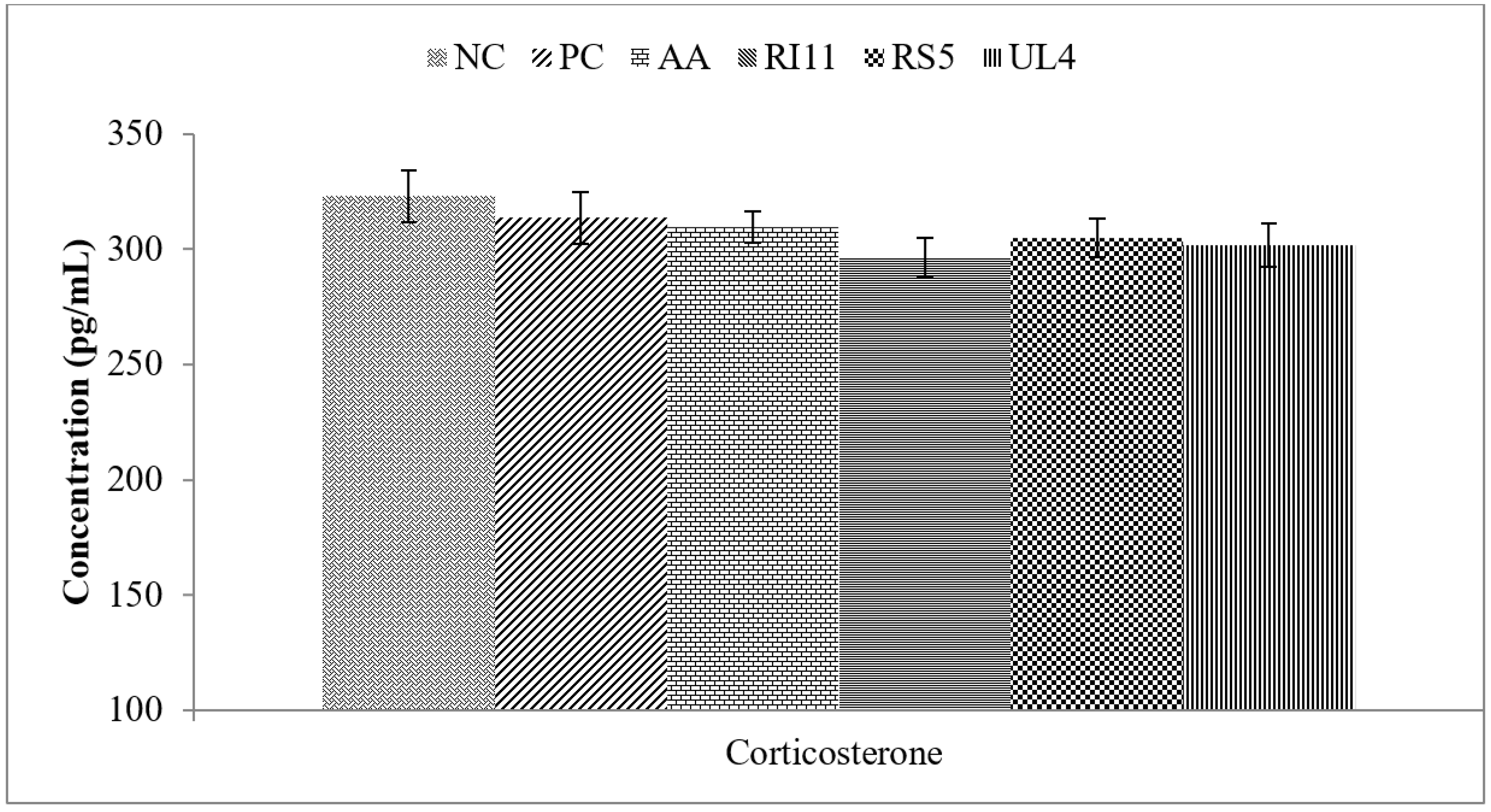
3.3. Plasma Corticosterone
3.4. Plasma Lipid Profile
3.5. Meat Quality (pH, Drip Loss, Cooking Loss, Shear Force and Colour) and Lipid Peroxidation (TBARS)
4. Discussion
4.1. Antioxidant Activities and Lipid Peroxidation
4.2. Acute Phase Proteins, HSP70 and Corticosterone
4.3. Plasma Lipid Profile
4.4. Meat Quality
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mahmoud, K.Z.; Edens, F.W.; Eisen, E.J.; Havenstein, G.B. Effect of ascorbic acid and acute heat exposure on heat shock protein 70 expression by young white Leghorn chickens. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 136, 329–335. [Google Scholar] [CrossRef]
- Mahmoud, K.Z.; Edens, F.W.; Eisen, E.J.; Havenstein, G.B. Ascorbic acid decreases heat shock protein 70 and plasma corticosterone response in broilers (Gallus gallus domesticus) subjected to cyclic heat stress. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 137, 35–42. [Google Scholar] [CrossRef]
- He, X.; Lu, Z.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Effects of chronic heat exposure on growth performance, intestinal epithelial histology, appetite-related hormones and genes expression in broilers. J. Sci. Food Agric. 2018, 98, 4471–4478. [Google Scholar] [CrossRef]
- Najafi, P.; Zulkifli, I.; Jajuli, N.A.; Farjam, A.S.; Ramiah, S.K.; Amir, A.A.; O‘Reily, E.; Eckersall, D. Environmental temperature and stocking density effects on acute phase proteins, heat shock protein 70, circulating corticosterone and performance in broiler chickens. Int. J. Biometeorol. 2015, 59, 1577–1583. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M.; Pérez-Gómez, C.; De Castro, I.N. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- Mager, W.H.; De Kruijff, A. Stress-induced transcriptional activation. Microbiol. Mol. Biol. Rev. 1995, 59, 506–531. [Google Scholar] [CrossRef]
- Iwagami, Y. Changes in the ultrastructure of human cells related to certain biological responses under hyperthermic culture conditions. Hum. Cell 1996, 9, 353–366. [Google Scholar]
- Halliwell, B.; Cross, C.E. Oxygen-derived species: Their relation to human disease and environmental stress. Environ. Health Perspect. 1994, 102, 5–12. [Google Scholar]
- Halliwell, B. Role of free radicals in the neurodegenerative diseases. Drugs Aging 2001, 18, 685–716. [Google Scholar] [CrossRef]
- Rahimi, S.; Khaksefidi, A. A comparison between the effects of a probiotic (Bioplus 2B) and an antibiotic (virginiamycin) on the performance of broiler chickens under heat stress condition. Iran. J. Vet. Res. 2006, 7, 23–28. [Google Scholar]
- Zulkifli, I.; Abdullah, N.; Azrin, N.M.; Ho, Y. Growth performance and immune response of two commercial broiler strains fed diets containing Lactobacillus cultures and oxytetracycline under heat stress conditions. Br. Poult. Sci. 2000, 41, 593–597. [Google Scholar] [CrossRef]
- Ramiah, S.K.; Zulkifli, I.; Rahim, N.A.A.; Ebrahimi, M.; Meng, G.Y. Effects of two herbal extracts and virginiamycin supplementation on growth performance, intestinal microflora population and fatty acid composition in broiler chickens. Asian-Australas. J. Anim. Sci. 2014, 27, 375–382. [Google Scholar] [CrossRef]
- Odore, R.; De Marco, M.; Gasco, L.; Rotolo, L.; Meucci, V.; Palatucci, A.; Rubino, V.; Ruggiero, G.; Canello, S.; Guidetti, G. Cytotoxic effects of oxytetracycline residues in the bones of broiler chickens following therapeutic oral administration of a water formulation. Poult. Sci. 2015, 94, 1979–1985. [Google Scholar] [CrossRef]
- Shazali, N.; Foo, H.L.; Loh, T.C.; Choe, D.W.; Rahim, R.A. Prevalence of antibiotic resistance in lactic acid bacteria isolated from the faeces of broiler chicken in Malaysia. Gut Pathog. 2014, 6, 1. [Google Scholar] [CrossRef]
- Regulation, E. No 1831/2003 of the European Parliament and Council of 22 September 2003 on additives for use in animal nutrition. Off. J. Eur. Commun. 2003, 268, 29–43. [Google Scholar]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Tajkarimi, M.; Ibrahim, S.A. Antimicrobial activity of ascorbic acid alone or in combination with lactic acid on Escherichia coli O157: H7 in laboratory medium and carrot juice. Food Control 2011, 22, 801–804. [Google Scholar] [CrossRef]
- Verghese, R.J.; Ramya, S.; Kanungo, R. In vitro Antibacterial Activity of Vitamin C and in Combination with Ciprofloxacin against Uropathogenic Escherichia coli. J. Clin. Diagn. Res. 2017, 11, 1–5. [Google Scholar] [CrossRef]
- Njoku, P. Effect of dietary ascorbic acid (vitamin C) supplementation on the performance of broiler chickens in a tropical environment. Anim. Feed Sci. Technol. 1986, 16, 17–24. [Google Scholar] [CrossRef]
- Kadim, I.; Al-Qamshui, B.; Mahgoub, O.; Al-Marzooqi, W.; Johnson, E. Effect of seasonal temperatures and ascorbic acid supplementation on performance of broiler chickens maintained in closed and open-sided houses. Int. J. Poult. Sci. 2008, 7, 655–660. [Google Scholar] [CrossRef]
- Kutlu, H.; Forbes, J. Changes in growth and blood parameters in heat-stressed broiler chicks in response to dietary ascorbic acid. Livest. Prod. Sci. 1993, 36, 335–350. [Google Scholar] [CrossRef]
- Sahin, K.; Sahin, N.; Kucuk, O. Effects of chromium, and ascorbic acid supplementation on growth, carcass traits, serum metabolites, and antioxidant status of broiler chickens reared at a high ambient temperature (32 °C). Nutr. Res. 2003, 23, 225–238. [Google Scholar] [CrossRef]
- Ferreira, I.; Matos Junior, J.; Sgavioli, S.; Vicentini, T.; Morita, V.; Boleli, I. Vitamin C prevents the effects of high rearing temperatures on the quality of broiler thigh meat. Poult. Sci. 2015, 94, 841–851. [Google Scholar] [CrossRef]
- Thanh, N.T.; Loh, T.C.; Foo, H.L.; Hair-Bejo, M.; Azhar, B.K. Effects of feeding metabolite combinations produced by Lactobacillus plantarum on growth performance, faecal microbial population, small intestine villus height and faecal volatile fatty acids in broilers. Br. Poult. Sci. 2009, 50, 298–306. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.; Garcia-Varela, R.; Garcia, H.; Mata-Haro, V.; González-Córdova, A.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Kareem, K.Y.; Ling, F.H.; Chwen, L.T.; Foong, O.M.; Asmara, S.A. Inhibitory activity of postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin. Gut Pathog. 2014, 6, 23. [Google Scholar] [CrossRef]
- Choe, D.W.; Foo, H.L.; Loh, T.C.; Hair-Bejo, M.; Awis, Q.S. Inhibitory property of metabolite combinations produced from Lactobacillus plantarum strains. Pertanika J. Trop. Agric. Sci. 2013, 36, 79–88. [Google Scholar]
- Thanh, N.T.; Chwen, L.T.; Foo, H.L.; Hair-Bejo, M.; Kasim, A.B. Inhibitory activity of metabolites produced by strains of Lactobacillus plantarum isolated from Malaysian fermented food. Int. J. Probiot. Prebiot. 2010, 5, 37–44. [Google Scholar]
- Van Thu, T.; Foo, H.L.; Loh, T.C.; Bejo, M.H. Inhibitory activity and organic acid concentrations of metabolite combinations produced by various strains of Lactobacillus plantarum. Afr. J. Biotechnol. 2011, 10, 1359–1363. [Google Scholar]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Akit, H.; Samsudin, A.A. Effects of dietary postbiotic and inulin on growth performance, IGF1 and GHR mRNA expression, faecal microbiota and volatile fatty acids in broilers. BMC Vet. Res. 2016, 12, 163. [Google Scholar] [CrossRef]
- Loh, T.C.; Thanh, N.T.; Foo, H.L.; Hair-Bejo, M.; Azhar, B.K. Feeding of different levels of metabolite combinations produced by Lactobacillus plantarum on growth performance, fecal microflora, volatile fatty acids and villi height in broilers. Anim. Sci. J. 2010, 81, 205–214. [Google Scholar] [CrossRef]
- Rosyidah, M.; Loh, T.; Foo, H.; Cheng, X.; Bejo, M. Effect of feeding metabolites and acidifier on growth performance, faecal characteristics and microflora in broiler chickens. J. Anim. Vet. Adv. 2011, 10, 2758–2764. [Google Scholar]
- Loh, T.C.; Choe, D.W.; Foo, H.L.; Sazili, A.Q.; Bejo, M.H. Effects of feeding different postbiotic metabolite combinations produced by Lactobacillus plantarum strains on egg quality and production performance, faecal parameters and plasma cholesterol in laying hens. BMC Vet. Res. 2014, 10, 149. [Google Scholar] [CrossRef]
- Choe, D.W.; Loh, T.C.; Foo, H.L.; Hair-Bejo, M.; Awis, Q.S. Egg production, faecal pH and microbial population, small intestine morphology, and plasma and yolk cholesterol in laying hens given liquid metabolites produced by Lactobacillus plantarum strains. Br. Poult. Sci. 2012, 53, 106–115. [Google Scholar] [CrossRef]
- Loh, T.C.; Thu, T.V.; Foo, H.L.; Bejo, M.H. Effects of different levels of metabolite combination produced by Lactobacillus plantarum on growth performance, diarrhoea, gut environment and digestibility of postweaning piglets. J. Appl. Anim. Res. 2013, 41, 200–207. [Google Scholar] [CrossRef]
- Thu, T.V.; Loh, T.C.; Foo, H.L.; Yaakub, H.; Bejo, M.H. Effects of liquid metabolite combinations produced by Lactobacillus plantarum on growth performance, faeces characteristics, intestinal morphology and diarrhoea incidence in postweaning piglets. Trop. Anim. Health Prod. 2011, 43, 69–75. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Loh, T.C.; Samsudin, A.A.; Foo, H.L. In vitro study of postbiotics from Lactobacillus plantarum RG14 on rumen fermentation and microbial population. Rev. Bras. Zootec. 2018, 47. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Humam, A.M. Postbiotic L. plantarum RG14 improves ruminal epithelium growth, immune status and upregulates the intestinal barrier function in post-weaning lambs. Sci. Rep. 2019, 9, 9938. [Google Scholar] [CrossRef]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Asmara, S.A.; Akit, H. Influence of postbiotic RG14 and inulin combination on cecal microbiota, organic acid concentration, and cytokine expression in broiler chickens. Poult. Sci. 2016, 96, 966–975. [Google Scholar] [CrossRef]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Asmara, S.A.; Akit, H.; Abdulla, N.R.; Ooi, M.F. Carcass, meat and bone quality of broiler chickens fed with postbiotic and prebiotic combinations. Int. J. Probiot. Prebiot. 2015, 10, 23. [Google Scholar]
- Loh, T.; Thanh, N.; Foo, H.; Hair-Bejo, M. Effects of feeding metabolite combinations from Lactobacillus plantarum on plasma and breast meat lipids in Broiler Chickens. Rev. Bras. Cienc. Avic. 2013, 15, 307–316. [Google Scholar] [CrossRef]
- Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Mustapha, N.M.; Zulkifli, I.; Izuddin, W.I. Effects of Feeding Different Postbiotics Produced by Lactobacillus plantarum on Growth Performance, Carcass Yield, Intestinal Morphology, Gut Microbiota Composition, Immune Status, and Growth Gene Expression in Broilers under Heat Stress. Animals 2019, 9, 644. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary Postbiotic Lactobacillus plantarum Improves Serum and Ruminal Antioxidant Activity and Upregulates Hepatic Antioxidant Enzymes and Ruminal Barrier Function in Post-Weaning Lambs. Antioxidants 2020, 9, 250. [Google Scholar] [CrossRef]
- He, Z.; Wang, X.; Li, G.; Zhao, Y.; Zhang, J.; Niu, C.; Zhang, L.; Zhang, X.; Ying, D.; Li, S. Antioxidant activity of prebiotic ginseng polysaccharides combined with potential probiotic Lactobacillus plantarum C88. Int. J. Food Sci. Technol. 2015, 50, 1673–1682. [Google Scholar] [CrossRef]
- Ji, K.; Jang, N.Y.; Kim, Y.T. Isolation of Lactic Acid Bacteria Showing Antioxidative and Probiotic Activities from Kimchi and Infant Feces. J. Microbiol. Biotechnol. 2015, 25, 1568–1577. [Google Scholar] [CrossRef]
- Moghadam, M.S.; Foo, H.L.; Leow, T.C.; Rahim, R.A.; Loh, T.C. Novel Bacteriocinogenic Lactobacillus plantarum Strains and Their Differentiation by Sequence Analysis of 16 S rDNA, 16 S-23 S and 23 S-5 S Intergenic Spacer Regions and Randomly Amplified Polymorphic DNA Analysis. Food Technol. Biotechnol. 2010, 48, 476–483. [Google Scholar]
- Lim, Y.H.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdullah, N. Comparative studies of versatile extracellular proteolytic activities of lactic acid bacteria and their potential for extracellular amino acid productions as feed supplements. J. Anim. Sci. Biotechnol. 2019, 10, 15. [Google Scholar] [CrossRef]
- Foo, H.; Loh, T.; Law, F.; Lim, Y.; Kuflin, C.; Rusul, G. Effect of feeding L. plantarum I-UL4 isolated from Malaysian Tempeh on growth performance, fecla flora and lactic acid bacteria and plasma cholesterol concentrations in post weaning rats. J. Food Sci. Biotechnol. 2003, 12, 403–408. [Google Scholar]
- Loh, T.; Chong, S.; Foo, H.; Law, F. Effects on growth performance, faecal microflora and plasma cholesterol after supplementation of spray-dried metabolite to postweaning rats. Czech J. Anim. Sci. 2009, 54, 10–16. [Google Scholar] [CrossRef]
- Malaysian Standard, M. Halal Food—Production, Preparation, Handling and Storage—General Guideline; Department of Standards Malaysia: Cyberjaya, Malaysia, 2009; pp. 1–26. Available online: http://www.standardsmalaysia.gov.my (accessed on 26 July 2019).
- DeLong, D.M.; DeLong, E.R.; Wood, P.D.; Lippel, K.; Rifkind, B.M. A comparison of methods for the estimation of plasma low-and very low-density lipoprotein cholesterol: The Lipid Research Clinics Prevalence Study. JAMA 1986, 256, 2372–2377. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.M.; Frei, B. Mechanisms of copper-and iron-dependent oxidative modification of human low density lipoprotein. J. Lipid Res. 1993, 34, 1745–1753. [Google Scholar]
- Mercier, Y.; Gatellier, P.; Viau, M.; Remignon, H.; Renerre, M. Effect of dietary fat and vitamin E on colour stability and on lipid and protein oxidation in turkey meat during storage. Meat Sci. 1998, 48, 301–318. [Google Scholar] [CrossRef]
- Martinez-Subiela, S.; Tecles, F.; Ceron, J. Comparison of two automated spectrophotometric methods for ceruloplasmin measurement in pigs. Res. Vet. Sci. 2007, 83, 12–19. [Google Scholar] [CrossRef]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Sazili, A.Q.; Parr, T.; Sensky, P.L.; Jones, S.W.; Bardsley, R.G.; Buttery, P.J. The relationship between slow and fast myosin heavy chain content, calpastatin and meat tenderness in different ovine skeletal muscles. Meat Sci. 2005, 69, 17–25. [Google Scholar] [CrossRef]
- American Meat Science Association. AMSA Meat Color Measurement Guidelines; American Meat Science Association: Savoy, IL, USA, 2012; pp. 1–136. Available online: http://www.meatscience.org (accessed on 5 March 2020).
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef]
- Altan, O.; Pabuccuoglu, A.; Altan, A.; Konyalioglu, S.; Bayraktar, H. Effect of heat stress on oxidative stress, lipid peroxidation and some stress parameters in broilers. Br. Poult. Sci. 2003, 44, 545–550. [Google Scholar] [CrossRef]
- Bai, K.; Huang, Q.; Zhang, J.; He, J.; Zhang, L.; Wang, T. Supplemental effects of probiotic Bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poult. Sci. 2017, 96, 74–82. [Google Scholar] [CrossRef]
- Surai, P.F. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef]
- Ko, Y.; Yang, H.; Jang, I. Effect of conjugated linoleic acid on intestinal and hepatic antioxidant enzyme activity and lipid peroxidation in broiler chickens. Asian-Australas. J. Anim. Sci. 2004, 17, 1162–1167. [Google Scholar] [CrossRef]
- Yang, P.C.; Tu, Y.H.; Perdue, M.H.; Oluwole, C.; Struiksma, S. Regulatory effect of heat shock protein 70 in stress-induced rat intestinal epithelial barrier dysfunction. N. Am. J. Med. Sci. 2009, 1, 9–15. [Google Scholar] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Xin, J.; Luo, M.; Khalique, A.; Dan, Y.; Pan, K.; Jing, B. Probiotic Lactobacillus johnsonii BS15 improves blood parameters related to immunity in broilers experimentally infected with subclinical necrotic enteritis. Front. Microbiol. 2018, 9, 49. [Google Scholar] [CrossRef]
- Akbarian, A.; Michiels, J.; Golian, A.; De Smet, S. Fourteen days cyclic heat challenge and feeding oreganum compactum and curcuma xanthorrhiza essential oils: Effects on antioxidant system of broilers. Commun. Agric. Appl. Biol. Sci. 2014, 79, 147–151. [Google Scholar]
- Foo, H.L.; Loh, T.C.; Abdul Mutalib, N.E.; Rahim, R.A. The myth and therapeutic potentials of postbiotics. In Microbiome and Metabolome in Diagnosis, Therapy, and Other Strategic Applications: Academic Press, 1st ed.; Faintuch, J., Faintuch, S., Eds.; John Fedor: Washington, WA, USA, 2019; Volume 1, pp. 201–208. [Google Scholar]
- Gao, J.; Li, Y.; Wan, Y.; Hu, T.; Liu, L.; Yang, S.; Gong, Z.; Zeng, Q.; Wei, Y.; Yang, W. A novel postbiotic from Lactobacillus rhamnosus GG with a beneficial effect on intestinal barrier function. Front. Microbiol. 2019, 10, 477. [Google Scholar] [CrossRef]
- Deng, W.; Dong, X.; Tong, J.; Zhang, Q. The probiotic Bacillus licheniformis ameliorates heat stress-induced impairment of egg production, gut morphology, and intestinal mucosal immunity in laying hens. Poult. Sci. 2012, 91, 575–582. [Google Scholar] [CrossRef]
- Havenaar, R.; Ten Brink, B.; Huis Veld, J.H.J. Selection of strains for probiotic use. In Probiotics: The Scientific Basis, 1st ed.; Fuller, R., Ed.; Chapman and Hall: London, UK, 1992; pp. 209–224. [Google Scholar]
- Li, N.; Yu, H.; Liu, H.; Wang, Y.; Zhou, J.; Ma, X.; Wang, Z.; Sun, C.; Qiao, S. Horizontal transfer of vanA between probiotic Enterococcus faecium and Enterococcus faecalis in fermented soybean meal and in digestive tract of growing pigs. J. Anim. Sci. Biotechnol. 2019, 10, 36. [Google Scholar] [CrossRef]
- Shen, X.; Yi, D.; Ni, X.; Zeng, D.; Jing, B.; Lei, M.; Bian, Z.; Zeng, Y.; Li, T.; Xin, J. Effects of Lactobacillus plantarum on production performance, immune characteristics, antioxidant status, and intestinal microflora of bursin-immunized broilers. Can. J. Microbiol. 2014, 60, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Cramer, T.A.; Kim, H.W.; Chao, Y.; Wang, W.; Cheng, H.W.; Kim, Y.H.B. Effects of probiotic (Bacillus subtilis) supplementation on meat quality characteristics of breast muscle from broilers exposed to chronic heat stress. Poult. Sci. 2018, 97, 3358–3368. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.L.; Li, W.; Shi, Y.H.; Le, G.W. Lipoic acid prevents high-fat diet-induced dyslipidemia and oxidative stress: A microarray analysis. Nutrition 2008, 24, 582–588. [Google Scholar] [CrossRef]
- Bai, W.K.; Zhang, F.J.; He, T.J.; Su, P.W.; Ying, X.Z.; Zhang, L.L.; Wang, T. Dietary Probiotic Bacillus subtilis Strain fmbj Increases Antioxidant Capacity and Oxidative Stability of Chicken Breast Meat during Storage. PLoS ONE 2016, 11, e0167339. [Google Scholar] [CrossRef] [PubMed]
- Najafi, P.; Zulkifli, I.; Soleimani, A.F.; Goh, Y.M. Acute phase proteins response to feed deprivation in broiler chickens. Poult. Sci. 2016, 95, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Najafi, P.; Nurfarahin, A.J.; Soleimani, A.F.; Kumari, S.; Aryani, A.A.; O‘Reilly, E.L.; Eckersall, P.D. Acute phase proteins, interleukin 6, and heat shock protein 70 in broiler chickens administered with corticosterone. Poult. Sci. 2014, 93, 3112–3118. [Google Scholar] [CrossRef]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Ijaz, A.; Sohail, A.; Shabbir, M.Z.; Rehman, H. Effect of supplementation of prebiotic mannan-oligosaccharides and probiotic mixture on growth performance of broilers subjected to chronic heat stress. Poult. Sci. 2012, 91, 2235–2240. [Google Scholar] [CrossRef] [PubMed]
- Al-Aqil, A.; Zulkifli, I. Changes in heat shock protein 70 expression and blood characteristics in transported broiler chickens as affected by housing and early age feed restriction. Poult. Sci. 2009, 88, 1358–1364. [Google Scholar] [CrossRef]
- Zulkifli, I.; Al-Aqil, A.; Omar, A.R.; Sazili, A.Q.; Rajion, M.A. Crating and heat stress influence blood parameters and heat shock protein 70 expression in broiler chickens showing short or long tonic immobility reactions. Poult. Sci. 2009, 88, 471–476. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. Neonatal feed restriction modulates circulating levels of corticosterone and expression of glucocorticoid receptor and heat shock protein 70 in aged Japanese quail exposed to acute heat stress. Poult. Sci. 2011, 90, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Malago, J.J.; Koninkx, J.F.; van Dijk, J.E. The heat shock response and cytoprotection of the intestinal epithelium. Cell Stress Chaperones 2002, 7, 191–199. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Hair-Bejo, M.; Omar, A.R.; Raha, A.R. The role of heat shock protein 70 in resistance to Salmonella enteritidis in broiler chickens subjected to neonatal feed restriction and thermal stress. Poult. Sci. 2012, 91, 340–345. [Google Scholar] [CrossRef]
- Heydari, A.R.; Wu, B.; Takahashi, R.; Strong, R.; Richardson, A. Expression of heat shock protein 70 is altered by age and diet at the level of transcription. Mol. Cell. Biol. 1993, 13, 2909–2918. [Google Scholar] [CrossRef]
- Yu, J.; Bao, E. Effect of acute heat stress on heat shock protein 70 and its corresponding mRNA expression in the heart, liver, and kidney of broilers. Asian-Australas. J. Anim. Sci. 2008, 21, 1116–1126. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. The relationship between adrenocortical function and Hsp70 expression in socially isolated Japanese quail. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 161, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Bao, E.; Yu, J. Heat shock protein 60 expression in heart, liver and kidney of broilers exposed to high temperature. Res. Vet. Sci. 2009, 86, 533–538. [Google Scholar] [CrossRef]
- Kareem, K.Y. Effect of Postbiotic and Inulin Supplements on Growth Performance, Gut Morthology, Gene Expression and Fecal Characteristics of Broiler Chickens. Ph.D. Thesis, University Putra Malaysia, Selangor, Malaysia, 2016. [Google Scholar]
- Thu, T.; Chwen, L.; Foo, H.; Halimatun, Y.; Bejo, M. Effects of metabolite combinations produced by Lactobacillus plantarum on plasma cholesterol and fatty acids in piglets. Am. J. Anim. Vet. Sci. 2010, 5, 233–236. [Google Scholar] [CrossRef]
- Pereira, D.I.; Gibson, G.R. Cholesterol assimilation by Lactic acid bacteria and Bifidobacteria isolated from the human gut. Appl. Environ. Microbiol. 2002, 68, 4689–4693. [Google Scholar] [CrossRef]
- Noh, D.O.; Kim, S.H.; Gilliland, S.E. Incorporation of cholesterol into the cellular membrane of Lactobacillus acidophilus ATCC 43121. J. Dairy Sci. 1997, 80, 3107–3113. [Google Scholar] [CrossRef]
- Den Hertog-Meischke, M.J.; van Laack, R.J.; Smulders, F.J. The water-holding capacity of fresh meat. Vet. Q. 1997, 19, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Sandercock, D.A.; Hunter, R.R.; Nute, G.R.; Mitchell, M.A.; Hocking, P.M. Acute heat stress-induced alterations in blood acid-base status and skeletal muscle membrane integrity in broiler chickens at two ages: Implications for meat quality. Poult. Sci. 2001, 80, 418–425. [Google Scholar] [CrossRef]
- Liu, H.W.; Li, K.; Zhao, J.S.; Deng, W. Effects of chestnut tannins on intestinal morphology, barrier function, pro-inflammatory cytokine expression, microflora and antioxidant capacity in heat-stressed broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.H.; Liang, R.R.; Lin, H.; Zhu, L.X.; Zhang, Y.M.; Mao, Y.W.; Dong, P.C.; Niu, L.B.; Zhang, M.H.; Luo, X. Effect of acute heat stress and slaughter processing on poultry meat quality and postmortem carbohydrate metabolism. Poult. Sci. 2017, 96, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Jia, G.Q.; Zuo, J.J.; Zhang, Y.; Lei, J.; Ren, L.; Feng, D.Y. Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat. Poult. Sci. 2012, 91, 2931–2937. [Google Scholar] [CrossRef] [PubMed]
- Zaboli, G.; Huang, X.; Feng, X.; Ahn, D.U. How can heat stress affect chicken meat quality—A review? Poult. Sci. 2019, 98, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Y.; Gu, Q.; Li, W. Effect of dietary probiotic, Bacillus coagulans, on growth performance, chemical composition, and meat quality of Guangxi Yellow chicken. Poult. Sci. 2010, 89, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.H. Probiotics feed supplement to improve quality of broiler chicken carcasses. World J. Dairy Food Sci. 2010, 5, 93–99. [Google Scholar]
- Zheng, A.; Luo, J.; Meng, K.; Li, J.; Zhang, S.; Li, K.; Liu, G.; Cai, H.; Bryden, W.L.; Yao, B. Proteome changes underpin improved meat quality and yield of chickens (Gallus gallus) fed the probiotic Enterococcus faecium. BMC Genom. 2014, 15, 1167. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, C.Z.; Richards, M.P.; O‘Keefe, S.F.; Wang, H. The effect of blood removal on oxidation and shelf life of broiler breast meat. Poult. Sci. 2007, 86, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Betti, M.; Bianchi, M.; Cavani, C. Color variation and characterization of broiler breast meat during processing in Italy. Poult. Sci. 2004, 83, 2086–2092. [Google Scholar] [CrossRef]
- Castellini, C.; Mugnai, C.; Dal Bosco, A. Effect of organic production system on broiler carcass and meat quality. Meat Sci. 2002, 60, 219–225. [Google Scholar] [CrossRef]
- Karaoglu, M.; Aksu, M.; Esenbuga, N.; Kaya, M.; Macit, M.; Durdag, H. Effect of dietary probiotic on the pH and colour characteristics of carcasses, breast fillets and drumsticks of broilers. Anim. Sci. 2004, 78, 253–259. [Google Scholar] [CrossRef]
- Aksu, M.; Esenbuga, N.; Macit, M. pH and colour characteristics of carcasses of broilers fed with dietary probiotics and slaughtered at different ages. Asian-Australas. J. Anim. Sci. 2006, 19, 605–610. [Google Scholar]
- Shimokomakillal, A.O.M. The effects of biotic additives on growth performance and meat qualities in broiler chickens. Int. J. Poult. Sci. 2012, 11, 599–604. [Google Scholar] [CrossRef]
- Hao, Y.; Gu, X. Effects of heat shock protein 90 expression on pectoralis major oxidation in broilers exposed to acute heat stress. Poult. Sci. 2014, 93, 2709–2717. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wen, J.; Zhang, H. Effect of chronic heat exposure on fat deposition and meat quality in two genetic types of chicken. Poult. Sci. 2007, 86, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, M.; Zheng, S.; Xie, P.; Ma, A. Effects of high temperature on multiple parameters of broilers in vitro and in vivo. Poult. Sci. 2008, 87, 2133–2139. [Google Scholar] [CrossRef]
- Sato, H.; Takahashi, T.; Sumitani, K.; Takatsu, H.; Urano, S. Glucocorticoid Generates ROS to Induce Oxidative Injury in the Hippocampus, Leading to Impairment of Cognitive Function of Rats. J. Clin. Biochem. Nutr. 2010, 47, 224–232. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | Diets 1 | SEM | p-Values | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| T-AOC (mM) | 0.63 c | 0.67 b,c | 0.68 b,c | 0.79 a | 0.76 a,b | 0.69 b,c | 0.017 | 0.028 |
| GPx (µmol/L) | 529.65 | 569.23 | 530.2 | 678.22 | 569.01 | 672.9 | 24.64 | 0.286 |
| CAT (U/L) | 3.95 b | 4.27 b | 4.46 a,b | 5.01 a | 4.39 a,b | 4.48 a,b | 0.096 | 0.046 |
| GSH (µM) | 42.29 b | 42.20 b | 45.30 a,b | 46.72 a | 45.71 a,b | 46.22 a | 0.534 | 0.026 |
| SOD (U/mL) | 0.955 | 0.959 | 0.981 | 1.190 | 0.944 | 1.069 | 0.028 | 0.129 |
| Parameter | Diets 1 | SEM | p-Values | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| Cholesterol | 3.46 a | 2.816 a,b | 2.46 b | 1.78 c | 1.916 b,c | 2.033 b,c | 0.149 | 0.003 |
| Triglyceride | 1.016 a | 0.77 a,b | 0.64 a,b | 0.416 b | 0.456 b | 0.34 b | 0.070 | 0.043 |
| VLDL | 0.203 a | 0.154 a,b | 0.128 a,b | 0.083 b | 0.091 b | 0.068 b | 0.014 | 0.043 |
| LDL | 0.783 a | 0.638 a | 0.446 b | 0.403 b | 0.386 b | 0.408 b | 0.033 | <0.0001 |
| HDL | 1.1883 | 1.44 | 1.415 | 2.19 | 1.8783 | 1.74 | 0.104 | 0.126 |
| Parameter | Diets 1 | SEM | p-Values | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| pH | 5.72 c | 5.77 c | 0.0007 | 6.01 a | 5.85 b,c | 5.95 a,b | 0.02 | 0.0007 |
| Drip loss % | 3.90 a | 2.90 a,b | 0.024 | 2.32 b | 2.87 a,b | 2.20 b | 0.15 | 0.024 |
| Cooking loss % | 21.64 a,b | 23.13 a | 0.009 | 17.89 c | 19.20 c | 19.38 b,c | 0.39 | 0.009 |
| Shear force (g) | 1113.8 a | 1021.6 a,b | 0.042 | 922.6 b | 949.1 b | 965.8 b | 18.25 | 0.042 |
| Colour | ||||||||
| L* (Lightness) | 49.02 a | 48.47 a | 45.82 b,c | 44.84 c | 47.32 ab | 45.96 b,c | 0.34 | 0.001 |
| a* (Redness) | 4.51 | 4.58 | 4.81 | 5.27 | 5.01 | 4.92 | 0.18 | 0.877 |
| b* (Yellowness) | 14.54 a,b | 15.60 a | 15.41 a | 12.63 c | 14.10 a,b,c | 13.18 b,c | 0.23 | 0.0003 |
| TBARS (µg MDA/g thigh meat) | 9.85 a | 8.91 a,b | 9.04 a,b | 7.01 c | 8.22 b,c | 7.37 c | 0.213 | <0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humam, A.M.; Loh, T.C.; Foo, H.L.; Izuddin, W.I.; Awad, E.A.; Idrus, Z.; Samsudin, A.A.; Mustapha, N.M. Dietary Supplementation of Postbiotics Mitigates Adverse Impacts of Heat Stress on Antioxidant Enzyme Activity, Total Antioxidant, Lipid Peroxidation, Physiological Stress Indicators, Lipid Profile and Meat Quality in Broilers. Animals 2020, 10, 982. https://doi.org/10.3390/ani10060982
Humam AM, Loh TC, Foo HL, Izuddin WI, Awad EA, Idrus Z, Samsudin AA, Mustapha NM. Dietary Supplementation of Postbiotics Mitigates Adverse Impacts of Heat Stress on Antioxidant Enzyme Activity, Total Antioxidant, Lipid Peroxidation, Physiological Stress Indicators, Lipid Profile and Meat Quality in Broilers. Animals. 2020; 10(6):982. https://doi.org/10.3390/ani10060982
Chicago/Turabian StyleHumam, Ali Merzza, Teck Chwen Loh, Hooi Ling Foo, Wan Ibrahim Izuddin, Elmutaz Atta Awad, Zulkifli Idrus, Anjas Asmara Samsudin, and Noordin Mohamed Mustapha. 2020. "Dietary Supplementation of Postbiotics Mitigates Adverse Impacts of Heat Stress on Antioxidant Enzyme Activity, Total Antioxidant, Lipid Peroxidation, Physiological Stress Indicators, Lipid Profile and Meat Quality in Broilers" Animals 10, no. 6: 982. https://doi.org/10.3390/ani10060982
APA StyleHumam, A. M., Loh, T. C., Foo, H. L., Izuddin, W. I., Awad, E. A., Idrus, Z., Samsudin, A. A., & Mustapha, N. M. (2020). Dietary Supplementation of Postbiotics Mitigates Adverse Impacts of Heat Stress on Antioxidant Enzyme Activity, Total Antioxidant, Lipid Peroxidation, Physiological Stress Indicators, Lipid Profile and Meat Quality in Broilers. Animals, 10(6), 982. https://doi.org/10.3390/ani10060982