Changes in Metabolites from Bovine Milk with β-Casein Variants Revealed by Metabolomics

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dairy Cow Selection and Milk Sample Collection
2.2. Nuclear Magnetic Resonance (NMR) Spectroscopy Analysis
2.3. Liquid Chromatography Mass Spectrometry (LC–MS) Analysis
2.4. Multivariate Statistical Analysis
3. Results
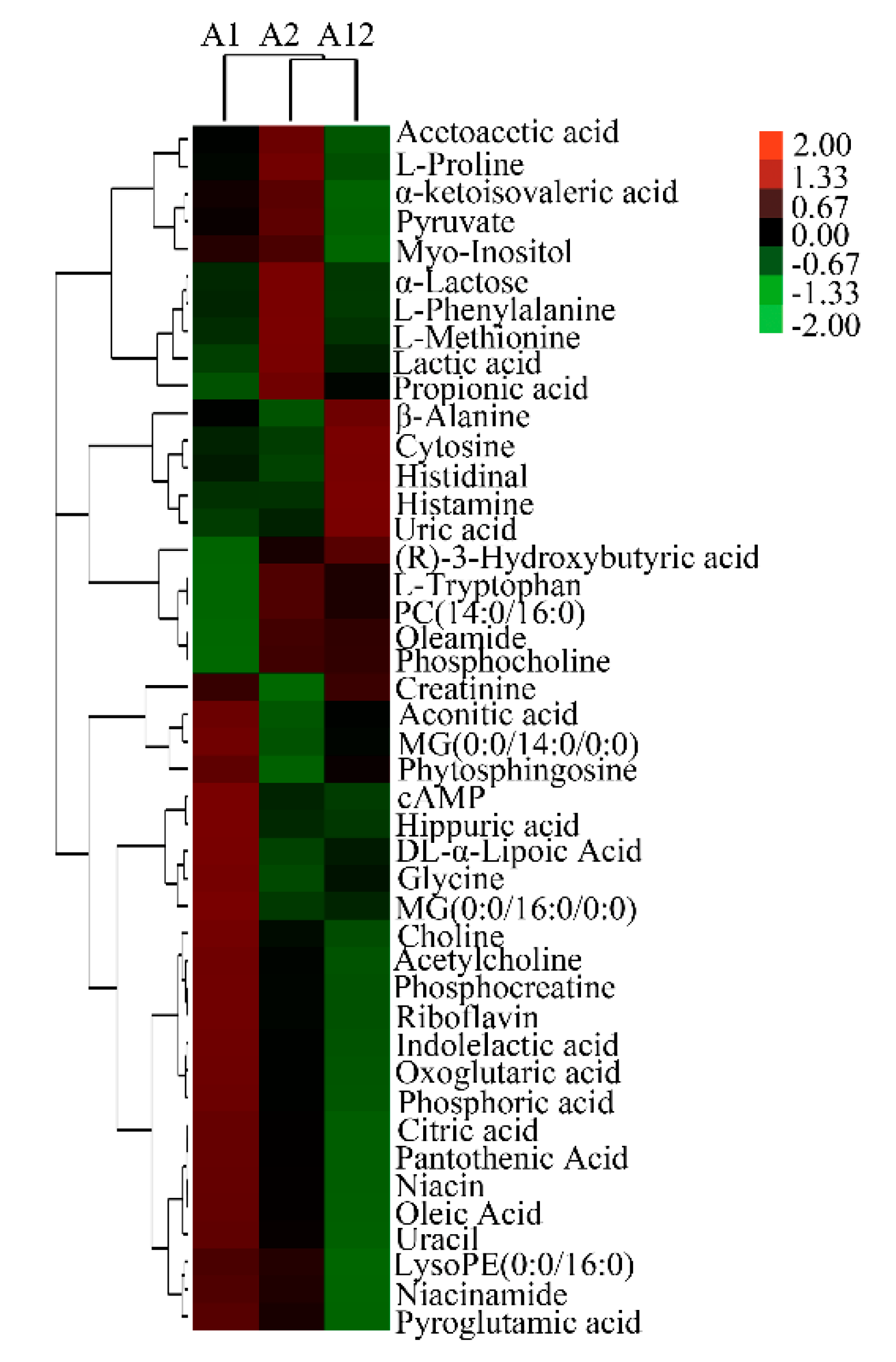
3.1. Nuclear Magnetic Resonance (NMR) Identification of Differential Milk Metabolites among β-Casein Variants
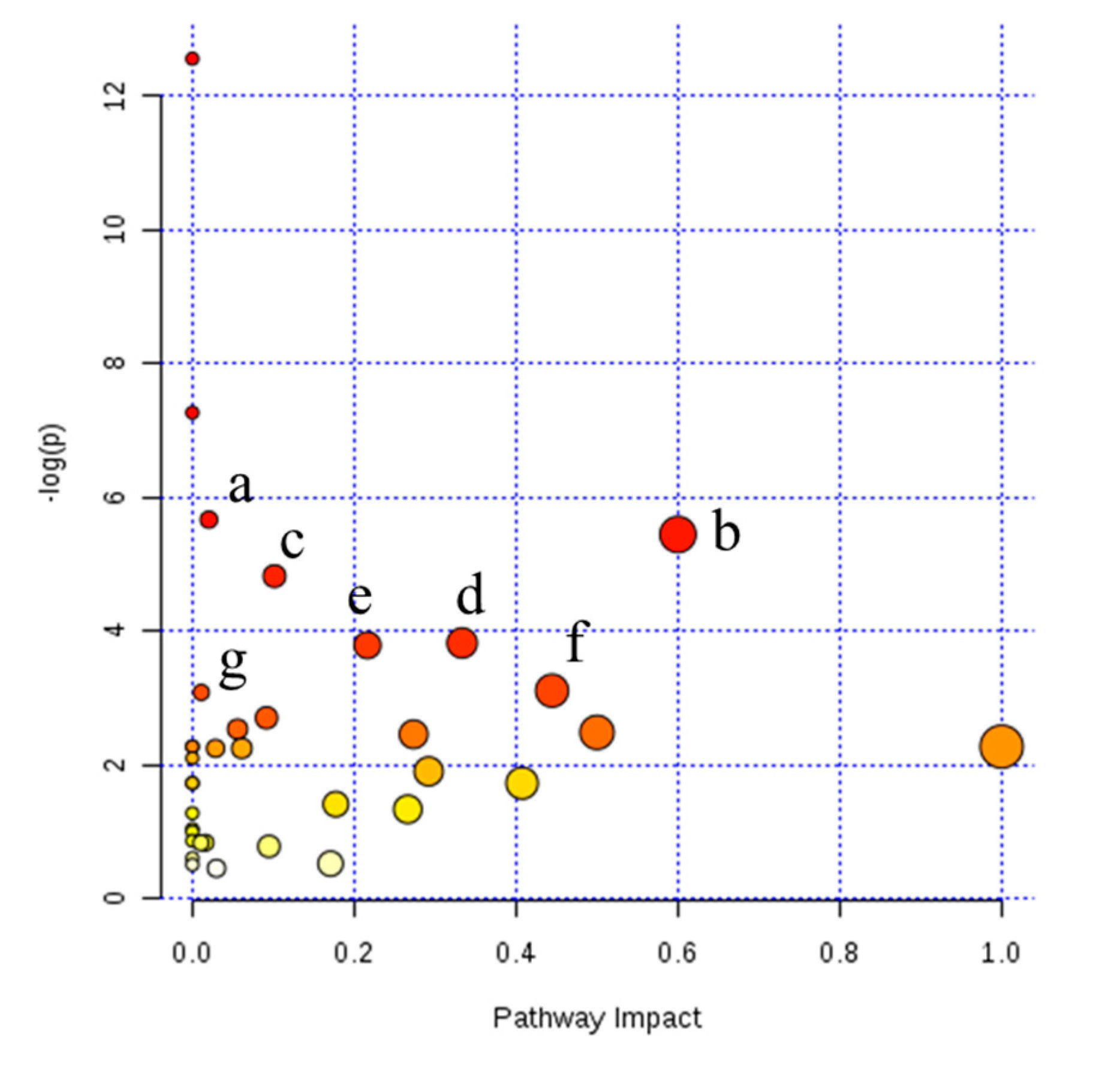
3.2. Liquid Chromatography Mass Spectrometry (LC–MS) Identification of Differential Milk Metabolites Among β-Casein Variants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rehan, F.; Ahemad, N.; Gupta, M. Casein nanomicelle as an emerging biomaterial—A comprehensive review. Colloids Surf. B 2019, 179, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Raynes, J.K.; Day, L.; Augustin, M.A.; Carver, J.A. Structural differences between bovine A(1) and A(2) beta-casein alter micelle self-assembly and influence molecular chaperone activity. J. Dairy Sci. 2015, 98, 2172–2182. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.M., Jr.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk-sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef]
- Cieslinska, A.; Fiedorowicz, E.; Zwierzchowski, G.; Kordulewska, N.; Jarmolowska, B.; Kostyra, E. Genetic polymorphism of beta-Casein gene in Polish Red Cattle-preliminary study of A1 and A2 frequency in genetic conservation herd. Animals (Basel) 2019, 9, 377. [Google Scholar] [CrossRef]
- Ng-Kwai-Hang, K.F.; Hayes, J.F.; Moxley, J.E.; Monardes, H.G. Association of genetic variants of casein and milk serum proteins with milk, fat, and protein production by dairy cattle. J. Dairy Sci. 1984, 67, 835–840. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Solah, V.A.; Busetti, F.; Smolenski, G.; Cooney, T. Application of ultra-high performance liquid chromatography coupled to high-resolution mass spectrometry (Orbitrap) for the determination of beta-casein phenotypes in cow milk. Food Chem. 2020, 307, 125532. [Google Scholar] [CrossRef]
- Fuerer, C.; Jenni, R.; Cardinaux, L.; Andetsion, F.; Wagniere, S.; Moulin, J.; Affolter, M. Protein fingerprinting and quantification of beta-casein variants by ultra-performance liquid chromatography-high-resolution mass spectrometry. J. Dairy Sci. 2020, 103, 1193–1207. [Google Scholar] [CrossRef]
- Gustavsson, F.; Buitenhuis, A.J.; Johansson, M.; Bertelsen, H.P.; Glantz, M.; Poulsen, N.A.; Lindmark Månsson, H.; Stålhammar, H.; Larsen, L.B.; Bendixen, C.; et al. Effects of breed and casein genetic variants on protein profile in milk from Swedish Red, Danish Holstein, and Danish Jersey cows. J. Dairy Sci. 2014, 97, 3866–3877. [Google Scholar] [CrossRef]
- Huang, W.; Penagaricano, F.; Ahmad, K.R.; Lucey, J.A.; Weigel, K.A.; Khatib, H. Association between milk protein gene variants and protein composition traits in dairy cattle. J. Dairy Sci. 2012, 95, 440–449. [Google Scholar] [CrossRef]
- Nguyen, H.T.H.; Schwendel, H.; Harland, D.; Day, L. Differences in the yoghurt gel microstructure and physicochemical properties of bovine milk containing A(1)A(1) and A(2)A(2) beta-casein phenotypes. Food Res. Int. 2018, 112, 217–224. [Google Scholar] [CrossRef]
- Boudonck, K.J.; Mitchell, M.W.; Wulff, J.; Ryals, J.A. Characterization of the biochemical variability of bovine milk using metabolomics. Metabolomics 2009, 5, 375–386. [Google Scholar] [CrossRef]
- Garwolinska, D.; Namiesnik, J.; Kot-Wasik, A.; Hewelt-Belka, W. Chemistry of human breast milk-A comprehensive review of the composition and role of milk metabolites in child development. J. Agric. Food Chem. 2018, 66, 11881–11896. [Google Scholar] [CrossRef] [PubMed]
- Caboni, P.; Murgia, A.; Porcu, A.; Manis, C.; Ibba, I.; Contu, M.; Scano, P. A metabolomics comparison between sheep’s and goat’s milk. Food Res. Int. 2019, 119, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zheng, N.; Zhao, X.; Zhang, Y.; Han, R.; Yang, J.; Zhao, S.; Li, S.; Guo, T.; Zang, C.; et al. Metabolomic biomarkers identify differences in milk produced by Holstein cows and other minor dairy animals. J. Proteomics 2016, 136, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Boiani, M.; Sundekilde, U.; Bateman, L.M.; McCarthy, D.G.; Maguire, A.R.; Gulati, A.; Guinee, T.P.; Fenelon, M.; Hennessy, D.; FitzGerald, R.J.; et al. Integration of high and low field 1H NMR to analyse the effects of bovine dietary regime on milk metabolomics and protein-bound moisture characterisation of the resulting mozzarella cheeses during ripening. Int. Dairy J. 2019, 91, 155–164. [Google Scholar] [CrossRef]
- Billa, P.A.; Faulconnier, Y.; Larsen, T.; Leroux, C.; Pires, J.A.A. Milk metabolites as noninvasive indicators of nutritional status of mid-lactation Holstein and Montbeliarde cows. J. Dairy Sci. 2020, 103, 3133–3146. [Google Scholar] [CrossRef]
- Tomassini, A.; Curone, G.; Solè, M.; Capuani, G.; Sciubba, F.; Conta, G.; Miccheli, A.; Vigo, D. NMR-based metabolomics to evaluate the milk composition from Friesian and autochthonous cows of Northern Italy at different lactation times. Nat. Prod. Res. 2019, 33, 1085–1091. [Google Scholar] [CrossRef]
- Wittenburg, D.; Melzer, N.; Willmitzer, L.; Lisec, J.; Kesting, U.; Reinsch, N.; Repsilber, D. Milk metabolites and their genetic variability. J. Dairy Sci. 2013, 96, 2557–2569. [Google Scholar] [CrossRef]
- Goodacre, R.; Vaidyanathan, S.; Dunn, W.B.; Harrigan, G.G.; Kell, D.B. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004, 22, 245–252. [Google Scholar] [CrossRef]
- Tenori, L.; Santucci, C.; Meoni, G.; Morrocchi, V.; Matteucci, G.; Luchinat, C. NMR metabolomic fingerprinting distinguishes milk from different farms. Food Res. Int. 2018, 113, 131–139. [Google Scholar] [CrossRef]
- Segers, K.; Declerck, S.; Mangelings, D.; Heyden, Y.V.; Eeckhaut, A.V. Analytical techniques for metabolomic studies: A review. Bioanalysis 2019, 11, 2297–2318. [Google Scholar] [CrossRef] [PubMed]
- Royo, L.J.; del Cerro, A.; Vicente, F.; Carballal, A.; de la Roza-Delgado, B. An accurate high-resolution melting method to genotype bovine β-casein. Eur. Food Res. Technol. 2014, 238, 295–298. [Google Scholar] [CrossRef]
- Zhao, X.W.; Qi, Y.X.; Huang, D.W.; Pan, X.C.; Cheng, G.L.; Zhao, H.L.; Yang, Y.X. Changes in serum metabolites in response to ingested colostrum and milk in neonatal calves, measured by nuclear magnetic resonance-based metabolomics analysis. J. Dairy Sci. 2018, 101, 7168–7181. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Dominguez, A.; Duran-Guerrero, E.; Fernandez-Recamales, A.; Lechuga-Sancho, A.M.; Sayago, A.; Schwarz, M.; Segundo, C.; Gonzalez-Dominguez, R. An overview on the importance of combining complementary analytical platforms in metabolomic research. Curr. Top. Med. Chem. 2017, 17, 3289–3295. [Google Scholar] [CrossRef]
- Batistel, F.; Arroyo, J.M.; Bellingeri, A.; Wang, L.; Saremi, B.; Parys, C.; Trevisi, E.; Cardoso, F.C.; Loor, J.J. Ethyl-cellulose rumen-protected methionine enhances performance during the periparturient period and early lactation in Holstein dairy cows. J. Dairy Sci. 2017, 100, 7455–7467. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, W.; Lin, X.Y.; Hu, Z.Y.; Yan, Z.G.; Wang, Y.; Shi, K.R.; Liu, G.M.; Wang, Z.H. Effects of rumen-protected methionine and other essential amino acid supplementation on milk and milk component yields in lactating Holstein cows. J. Dairy Sci. 2019, 102, 7936–7947. [Google Scholar] [CrossRef]
- Patton, R.A. Effect of rumen-protected methionine on feed intake, milk production, true milk protein concentration, and true milk protein yield, and the factors that influence these effects: A meta-analysis. J. Dairy Sci. 2010, 93, 2105–2118. [Google Scholar] [CrossRef]
- Ng-Kwai-Hang, K.F.; Monardes, H.G.; Hayes, J.F. Association between genetic polymorphism of milk proteins and production traits during three lactations. J. Dairy Sci. 1984, 73, 3414–3420. [Google Scholar] [CrossRef]
- Olenski, K.; Kamiński, S.; Szyda, J.; Cieslinska, A. Polymorphism of the beta-casein gene and its associations with breeding value for production traits of Holstein–Friesian bulls. Livest. Sci. 2010, 131, 137–140. [Google Scholar] [CrossRef]
- Wu, G.; Knabe, D.A. Free and protein-bound amino acids in sow’s colostrum and milk. J. Nutr. 1994, 124, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Bruckental, I.; Ascarelli, I.; Alumot, B.Y.E. Effect of duodenal proline infusion on milk production and composition in dairy cows. Anim. Sci. 2010, 53, 299–303. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino. Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Caldow, M.K.; Ham, D.J.; Trieu, J.; Chung, J.D.; Lynch, G.S.; Koopman, R. Glycine protects muscle cells from wasting in vitro via mTORC1 signaling. Front. Nutr. 2019, 6, 172. [Google Scholar] [CrossRef] [PubMed]
- Thelwall, P.E.; Simpson, N.E.; Rabbani, Z.N.; Clark, M.D.; Pourdeyhimi, R.; Macdonald, J.M.; Blackband, S.J.; Gamcsik, M.P. In vivo MR studies of glycine and glutathione metabolism in a rat mammary tumor. NMR Biomed. 2012, 25, 271–278. [Google Scholar] [CrossRef]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant properties dedicated to nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef]
- Wallace, T.C.; Blusztajn, J.K.; Caudill, M.A.; Klatt, K.C.; Natker, E.; Zeisel, S.H.; Zelman, K.M. Choline: The underconsumed and underappreciated essential nutrient. Nutr. Today 2018, 53, 240–253. [Google Scholar] [CrossRef]
- Wiedeman, A.M.; Whitfield, K.C.; March, K.M.; Chen, N.N.; Kroeun, H.; Sokhoing, L.; Sophonneary, P.; Dyer, R.A.; Xu, Z.; Kitts, D.D.; et al. Concentrations of water-soluble forms of choline in human milk from lactating women in Canada and Cambodia. Nutrients 2018, 10, 381. [Google Scholar] [CrossRef]
- Baldi, A.; Pinotti, L. Choline metabolism in high-producing dairy cows: Metabolic and nutritional basis. Can. J. Anim. Sci. 2006, 86, 207–212. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Zhong, Z.; Stubelius, A.; Sweeney, S.R.; Booshehri, L.M.; Antonucci, L.; Liu-Bryan, R.; Lodi, A.; Terkeltaub, R.; Lacal, J.C.; et al. Choline uptake and metabolism modulate macrophage IL-1beta and IL-18 production. Cell Metab. 2019, 29, 1350–1362. [Google Scholar] [CrossRef] [PubMed]
- Loizzi, R.F. Cyclic AMP changes in guinea pig mammary gland and milk. Am. J. Physiol. 1983, 245, E549–E554. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, G.; Bittante, G.; Polan, C.E. Comparison of fibrous materials as modifiers of in situ ruminal degradation of corn gluten meal. J. Dairy Sci. 1993, 76, 1106–1113. [Google Scholar] [CrossRef]
- Hernández, A.G.; Sánchez-Medina, F. The determination of acid-soluble nucleotides in milk by improved enzymic methods: A comparison with the ion-exchange column chromatography procedure. J. Sci. Food Agric. 1981, 32, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Tiemeyer, W.; Stohrer, M.; Giesecke, D. Metabolites of nucleic acids in bovine milk. J. Dairy Sci. 1984, 67, 723–728. [Google Scholar] [CrossRef]
- Schlimme, E.; Martin, D.; Meisel, H. Nucleosides and nucleotides: Natural bioactive substances in milk and colostrum. Br. J. Nutr. 2000, 84, S59–S68. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Metabolites | Chemical Shift a | Correlation Coefficients | log2(Fold -Change) | p-Value |
|---|---|---|---|---|
| heterozygote A1/A2 comparative with variant A1/A1 | ||||
| Valine | 1.04 (d), 1.00 (d) | 0.841 | 0.45 | 9.82 × 10−5 |
| 3-hydroxybutytrate | 1.20 (d) | 0.836 | 0.57 | 6.73 × 10−5 |
| Lactate | 1.33 (d),4.12 (q) | 0.826 | 0.41 | 8.44 × 10−5 |
| Alanine | 1.48 (d) | 0.785 | 0.49 | 3.08 × 10−4 |
| Lysine | 1.74 (m),1.90 (m) | 0.773 | 0.43 | 4.53 × 10−4 |
| Acetate | 1.92 (s) | 0.831 | 0.48 | 9.84 × 10−5 |
| Glutamate | 2.07 (m),2.35 (m) | 0.739 | 0.43 | 1.05 × 10−3 |
| Phenylalanine | 7.24 (m),7.30 (m),7.36 (m) | 0.88 | 0.68 | 1.69 × 10−6 |
| Histidine | 7.08 (s),7.85 (s) | 0.891 | 0.57 | 5.07 × 10−4 |
| Unknown metabolite | 6.20 (s) | 0.710 | 0.60 | 0.0019 |
| variant A2/A2 comparative with A1/A1 | ||||
| Valine | 1.04 (d), 1.00 (d) | 0.791 | 0.41 | 3.59 × 10−4 |
| 3-hydroxybutytrate | 1.20 (d) | 0.792 | 0.46 | 6.70 × 10−4 |
| Lactate | 1.33 (d),4.12 (q) | 0.844 | 0.39 | 6.45 × 10−5 |
| Alanine | 1.48 (d) | 0.751 | 0.42 | 1.09 × 10−3 |
| Lysine | 1.74 (m),1.90 (m) | 0.717 | 0.37 | 1.76 × 10−3 |
| Acetate | 1.92 (s) | 0.764 | 0.40 | 6.70 × 10−4 |
| Glutamate | 2.07 (m),2.35 (m) | 0.733 | 0.41 | 1.42 × 10−3 |
| Phenylalanine | 7.24 (m),7.30 (m),7.36 (m) | 0.693 | 0.57 | 1.27 × 10−3 |
| Histidine | 7.08 (s),7.85 (s) | 0.734 | 0.41 | 1.94 × 10−3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.; Liu, H.; Yang, Y.; Bu, D.; Zang, C.; Yang, K.; Yu, X.; Wang, J. Changes in Metabolites from Bovine Milk with β-Casein Variants Revealed by Metabolomics. Animals 2020, 10, 954. https://doi.org/10.3390/ani10060954
Lv Z, Liu H, Yang Y, Bu D, Zang C, Yang K, Yu X, Wang J. Changes in Metabolites from Bovine Milk with β-Casein Variants Revealed by Metabolomics. Animals. 2020; 10(6):954. https://doi.org/10.3390/ani10060954
Chicago/Turabian StyleLv, Zhongwang, Hui Liu, Yongxin Yang, Dengpan Bu, Changjiang Zang, Kailun Yang, Xiong Yu, and Jiaqi Wang. 2020. "Changes in Metabolites from Bovine Milk with β-Casein Variants Revealed by Metabolomics" Animals 10, no. 6: 954. https://doi.org/10.3390/ani10060954
APA StyleLv, Z., Liu, H., Yang, Y., Bu, D., Zang, C., Yang, K., Yu, X., & Wang, J. (2020). Changes in Metabolites from Bovine Milk with β-Casein Variants Revealed by Metabolomics. Animals, 10(6), 954. https://doi.org/10.3390/ani10060954