Temporary Relocation during Rest Periods: Relocation Stress and Other Factors Influence Hair Cortisol Concentrations in Horses

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Hair Sampling and Storage
2.2. Hair Steroid Extraction
2.3. Hair Cortisol Detection and Validation Tests
2.4. Study 1: Relocation Effect on HCCs
2.5. Study 2: Seasonal Effect on HCCs and Hair Growth
2.6. Statistical Analyses
3. Results
3.1. Biochemical Validation of the Enzyme Immunoassay
3.2. Study 1: Relocation Effect on HCCs
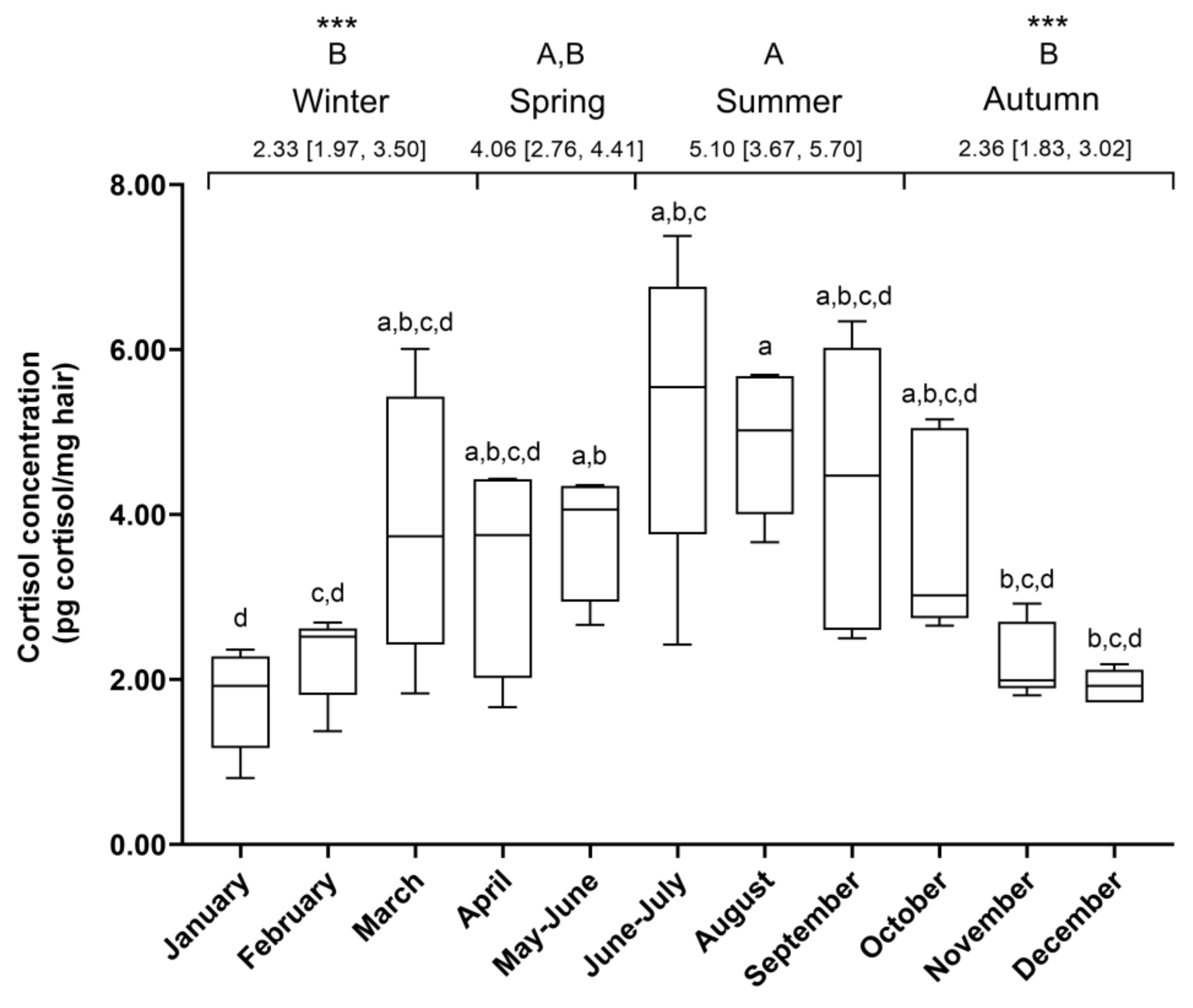
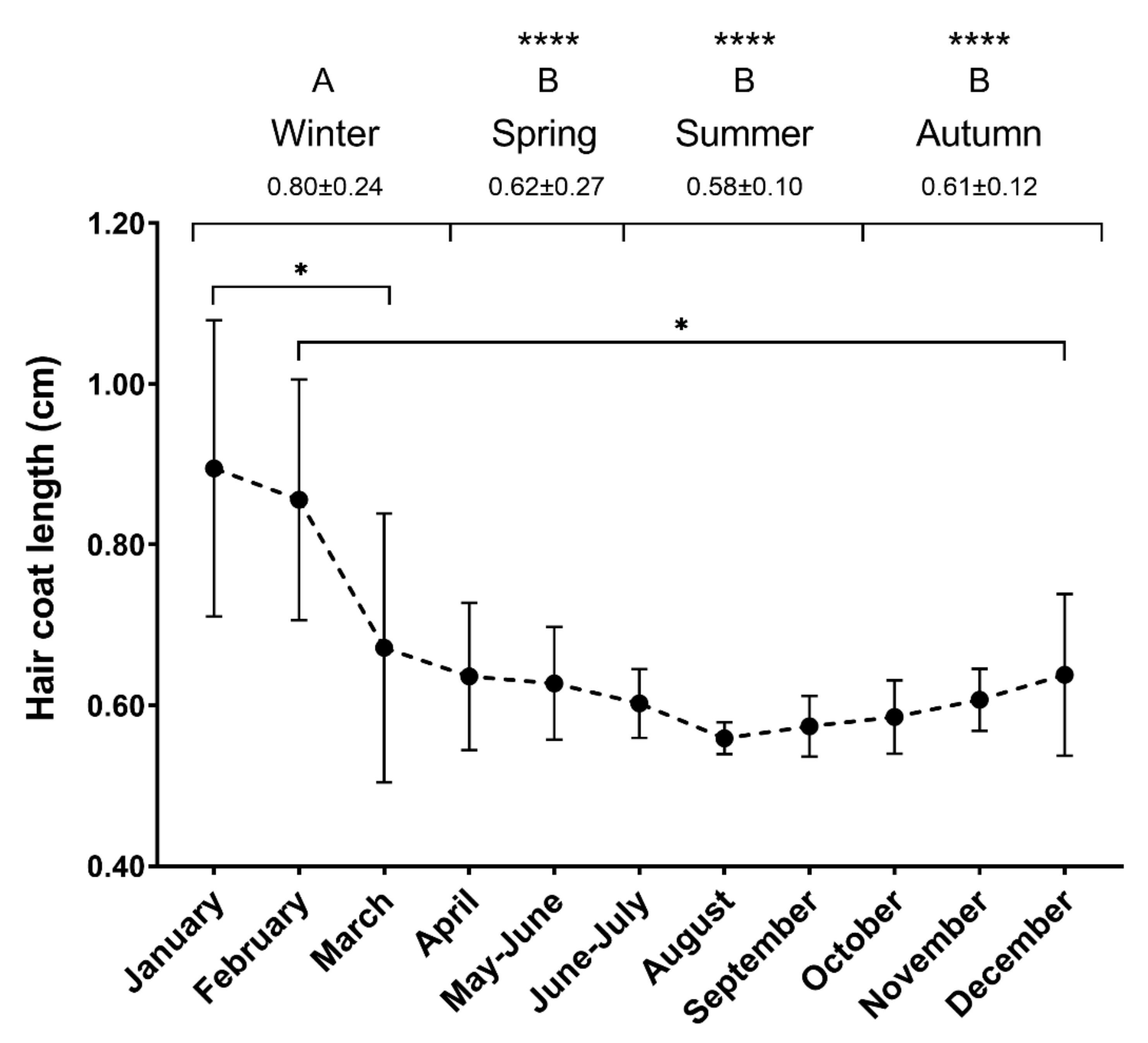
3.3. Study 2: Seasonal Effect on HCCs and Hair Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FEI Endurance Rules. Available online: https://inside.fei.org/node/3835/ (accessed on 3 February 2020).
- Mukai, K.; Ohmura, H.; Hiraga, A.; Eto, D.; Takahashi, T.; Asai, Y.; Jones, J.H. Effect of detraining on cardiorespiratory variables in young Thoroughbred horses. Equine Vet. J. Suppl. 2006, 36, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Hiraga, A.; Takahashi, T.; Matsui, A.; Ohmura, H.; Aida, H.; Jones, J.H. Effects of maintaining different exercise intensities during detraining on aerobic capacity in Thoroughbreds. Am. J. Vet. Res. 2017, 78, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic–pituitary–adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Bohák, Z.; Szabó, F.; Beckers, J.F.; Melo de Sousa, N.; Kutasi, O.; Nagy, K.; Szenci, O. Monitoring the circadian rhythm of serum and salivary cortisol concentrations in the horse. Domest. Anim. Endocrinol. 2013, 45, 38–42. [Google Scholar] [CrossRef]
- Pawluski, J.; Jego, P.; Henry, S.; Bruchet, A.; Palme, R.; Coste, C.; Hausberger, M. Low plasma cortisol and fecal cortisol metabolite measures as indicators of compromised welfare in domestic horses (Equus caballus). PLoS ONE 2017, 12, e0182257. [Google Scholar] [CrossRef] [PubMed]
- Mercer-Bowyer, S.; Kersey, D.C.; Bertone, J.J. Use of fecal glucocorticoid and salivary cortisol concentrations as a measure of well-being of New York City carriage horses. J. Am. Vet. Med. Assoc. 2017, 250, 316–321. [Google Scholar] [CrossRef]
- Van Der Kolk, J.H.; Kalsbeek, H.C.; Wensing, T.; Breukink, H.J. Urinary concentration of corticoids in normal horses and horses with hyperadrenocorticism. Res. Vet. Sci. 1994, 56, 126–128. [Google Scholar] [CrossRef]
- Heimbürge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef]
- Ghassemi, J.; Kim, B.W.; Lee, B.H.; Sung, K. Coat and hair color: Hair cortisol and serotonin levels in lactating Holstein cows under heat stress conditions. Anim. Sci. J. 2017, 88, 190–194. [Google Scholar] [CrossRef]
- Meyer, J.S.; Novak, M.A. Minireview: Hair cortisol: A novel biomarker of hypothalamic-pituitary-adrenocortical activity. Endocrinology 2012, 153, 4120–4127. [Google Scholar] [CrossRef]
- Henderson, G.L. Mechanisms of drug incorporation into hair. Forensic Sci. Int. 1993, 63, 19–29. [Google Scholar] [CrossRef]
- Davenport, M.D.; Lutz, C.K.; Tiefenbacher, S.; Novak, M.A.; Meyer, J.S. A Rhesus Monkey Model of Self-Injury: Effects of Relocation Stress on Behavior and Neuroendocrine Function. Biol. Psychiatry 2008, 63, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Tallo-Parra, O.; Lopez-Bejar, M.; Carbajal, A.; Monclús, L.; Manteca, X.; Devant, M. Acute ACTH-induced elevations of circulating cortisol do not affect hair cortisol concentrations in calves. Gen. Comp. Endocrinol. 2017, 240, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Del Rosario, G.D.L.V.; Lemus-Ramirez, V.; Vázquez-Chagoyán, J.C.; Villa-Godoy, A.; Romano, M.C. Effects of adrenocorticotropic hormone challenge and age on hair cortisol concentrations in dairy cattle. Can. J. Vet. Res. 2011, 75, 216–221. [Google Scholar]
- Terwissen, C.V.; Mastromonaco, G.F.; Murray, D.L. Influence of adrenocorticotrophin hormone challenge and external factors (age, sex, and body region) on hair cortisol concentration in Canada lynx (Lynx canadensis). Gen. Comp. Endocrinol. 2013, 194, 162–167. [Google Scholar] [CrossRef]
- Mastromonaco, G.F.; Gunn, K.; McCurdy-Adams, H.; Edwards, D.B.; Schulte-Hostedde, A.I. Validation and use of hair cortisol as a measure of chronic stress in eastern chipmunks (Tamias striatus). Conserv. Physiol. 2014, 2, 1–12. [Google Scholar] [CrossRef]
- Endo, N.; Yamane, H.; Rahayu, L.P.; Tanaka, T. Effect of repeated adrenocorticotropic hormone administration on reproductive function and hair cortisol concentration during the estrous cycle in goats. Gen. Comp. Endocrinol. 2018, 259, 207–212. [Google Scholar] [CrossRef]
- Ashley, N.T.; Barboza, P.S.; Macbeth, B.J.; Janz, D.M.; Cattet, M.R.L.; Booth, R.K.; Wasser, S.K. Glucocorticosteroid concentrations in feces and hair of captive caribou and reindeer following adrenocorticotropic hormone challenge. Gen. Comp. Endocrinol. 2011, 172, 382–391. [Google Scholar] [CrossRef]
- Schubach, K.M.; Cooke, R.F.; Brandao, A.P.; Lippolis, K.; Hinchliff, M.T.; Bohnert, D.W.; Cerri, R.L.A. Using hair cortisol concentrations to assess the adrenocortical stress response in beef cattle administered corticotrophin-release hormone. J. Anim. Sci. 2016, 94, 109. [Google Scholar] [CrossRef]
- Comin, A.; Veronesi, M.C.; Montillo, M.; Faustini, M.; Valentini, S.; Cairoli, F.; Prandi, A. Hair cortisol level as a retrospective marker of hypothalamic–pituitary–adrenal axis activity in horse foals. Vet. J. 2012, 194, 131–132. [Google Scholar] [CrossRef]
- Duran, M.C.; Janz, D.M.; Waldner, C.L.; Campbell, J.R.; Marques, F.J. Hair Cortisol Concentration as a Stress Biomarker in Horses: Associations with Body Location and Surgical Castration. J. Equine Vet. Sci. 2017, 55, 27–33. [Google Scholar] [CrossRef]
- Montillo, M.; Comin, A.; Corazzin, M.; Peric, T.; Faustini, M.; Veronesi, M.C.; Valentini, S.; Bustaffa, M.; Prandi, A. The Effect of temperature, rainfall, and light conditions on hair cortisol concentrations in newborn foals. J. Equine Vet. Sci. 2014, 34, 774–778. [Google Scholar] [CrossRef]
- MacDougall-Shackleton, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “Stress” Are Not Synonymous. Integr. Org. Biol. 2019, 1, 1–8. [Google Scholar] [CrossRef]
- Tallo-Parra, O.; Manteca, X.; Sabes-Alsina, M.; Carbajal, A.; Lopez-Bejar, M. Hair cortisol detection in dairy cattle by using EIA: Protocol validation and correlation with faecal cortisol metabolites. Animal 2015, 9, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.L.; Huntington, P.J. Body condition scoring and weight estimation of horses. Equine Vet. J. 1988, 20, 41–45. [Google Scholar] [CrossRef]
- Popescu, S.; Diugan, E.A.; Spinu, M. The interrelations of good welfare indicators assessed in working horses and their relationships with the type of work. Res. Vet. Sci. 2014, 96, 406–414. [Google Scholar] [CrossRef]
- Werhahn, H.; Hessel, E.F.; Van den Weghe, H.F.A. Competition Horses Housed in Single Stalls (II): Effects of Free Exercise on the Behavior in the Stable, the Behavior during Training, and the Degree of Stress. J. Equine Vet. Sci. 2012, 32, 22–31. [Google Scholar] [CrossRef]
- Ruet, A.; Lemarchand, J.; Parias, C.; Mach, N.; Moisan, M.P.; Foury, A.; Briant, C.; Lansade, L. Housing horses in individual boxes is a challenge with regard to welfare. Animals 2019, 9, 621. [Google Scholar] [CrossRef]
- Stull, C.L.; Rodiek, A.V. Responses of Blood Glucose, Insulin and Cortisol Concentrations to Common Equine Diets. J. Nutr. 1988, 118, 206–213. [Google Scholar] [CrossRef]
- Jacob, S.I.; Geor, R.J.; Weber, P.S.D.; Harris, P.A.; McCue, M.E. Effect of dietary carbohydrates and time of year on ACTH and cortisol concentrations in adult and aged horses. Domest. Anim. Endocrinol. 2018, 63, 15–22. [Google Scholar] [CrossRef]
- Comin, A.; Prandi, A.; Peric, T.; Corazzin, M.; Dovier, S.; Bovolenta, S. Hair cortisol levels in dairy cows from winter housing to summer highland grazing. Livest. Sci. 2011, 138, 69–73. [Google Scholar] [CrossRef]
- Peric, T.; Comin, A.; Corazzin, M.; Montillo, M.; Canavese, F.; Stebel, M.; Prandi, A. Relocation and Hair Cortisol Concentrations in New Zealand White Rabbits. J. Appl. Anim. Welf. Sci. 2017, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Toribio, R.E. Disorders of the Endocrine System. In Equine Internal Medicine; Reed, S.M., Bayly, W.M., Sellon, D.C., Eds.; Elsevier: St. Louis, MO, USA, 2018; pp. 1029–1138. [Google Scholar]
- Donaldson, M.T.; McDonnell, S.M.; Schanbacher, B.J.; Lamb, S.V.; McFarlane, D.; Beech, J. Variation in Plasma Adrenocorticotropic Hormone Concentration and Dexamethasone Suppression Test Results with Season, Age, and Sex in Healthy Ponies and Horses. J. Vet. Intern. Med. 2005, 19, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Cordero, M.; Brorsen, B.W.; McFarlane, D. Circadian and circannual rhythms of cortisol, ACTH, and α-melanocyte-stimulating hormone in healthy horses. Domest. Anim. Endocrinol. 2012, 43, 317–324. [Google Scholar] [CrossRef]
- Cymbaluk, N.F.; Christison, G.I. Effects of Diet and Climate on Growing Horses. J. Anim. Sci. 1989, 67, 48. [Google Scholar] [CrossRef]
- Stachurska, A.; Robovský, J.; Bocian, K.; Janczarek, I. Changes of coat cover in primitive horses living on a reserve. J. Anim. Sci. 2011, 93, 1411–1417. [Google Scholar] [CrossRef]
- Bocian, K.; Strzelec, K.; Janczarek, I.; Jabłecki, Z.; Kolstrung, R. Length of winter coat in horses depending on husbandry conditions. Anim. Sci. J. 2017, 88, 339–346. [Google Scholar] [CrossRef]
- Prinsloo, M.; Hynd, P.; Franklin, S.; Weaver, S.; van den Boom, R. Hair cortisol concentration is inversely related to the severity of equine squamous gastric disease. Vet. J. 2019, 249, 58–59. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic–pituitary–adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar]
- Tallo-Parra, O.; Carbajal, A.; Monclús, L.; Manteca, X.; Lopez-Bejar, M. Hair cortisol and progesterone detection in dairy cattle: Interrelation with physiological status and milk production. Domest. Anim. Endocrinol. 2018, 64, 1–8. [Google Scholar] [CrossRef]
- Burnett, T.A.; Madureira, A.M.L.; Silper, B.F.; Tahmasbi, A.; Nadalin, A.; Veira, D.M.; Cerri, R.L.A. Relationship of concentrations of cortisol in hair with health, biomarkers in blood, and reproductive status in dairy cows. J. Dairy Sci. 2015, 98, 4414–4426. [Google Scholar] [CrossRef] [PubMed]
- Macbeth, B.J.; Cattet, M.R.L.; Stenhouse, G.B.; Gibeau, M.L.; Janz, D.M. Hair cortisol concentration as a noninvasive measure of long-term stress in free-ranging grizzly bears (Ursus arctos): Considerations with implications for other wildlife. Can. J. Zool. 2010, 88, 935–949. [Google Scholar] [CrossRef]
- Bryan, H.M.; Adams, A.G.; Invik, R.M.; Wynne-Edwards, K.E.; Smits, J.E. Hair as a Meaningful Measure of Baseline Cortisol Levels over Time in Dogs. J. Am. Assoc. Lab. Anim. Sci. 2013, 52, 189–196. [Google Scholar] [PubMed]
- Aurich, J.; Wulf, M.; Ille, N.; Erber, R.; von Lewinski, M.; Palme, R.; Aurich, C. Effects of season, age, sex, and housing on salivary cortisol concentrations in horses. Domest. Anim. Endocrinol. 2015, 52, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Hart, K.A.; Wochele, D.M.; Norton, N.A.; Mcfarlane, D.; Wooldridge, A.A.; Frank, N. Effect of Age, Season, Body Condition, and Endocrine Status on Serum Free Cortisol Fraction and Insulin Concentration in Horses. J. Vet. Intern. Med. 2016, 30, 653–663. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardela, J.; Carbajal, A.; Tallo-Parra, O.; Olvera-Maneu, S.; Álvarez-Rodríguez, M.; Jose-Cunilleras, E.; López-Béjar, M. Temporary Relocation during Rest Periods: Relocation Stress and Other Factors Influence Hair Cortisol Concentrations in Horses. Animals 2020, 10, 642. https://doi.org/10.3390/ani10040642
Gardela J, Carbajal A, Tallo-Parra O, Olvera-Maneu S, Álvarez-Rodríguez M, Jose-Cunilleras E, López-Béjar M. Temporary Relocation during Rest Periods: Relocation Stress and Other Factors Influence Hair Cortisol Concentrations in Horses. Animals. 2020; 10(4):642. https://doi.org/10.3390/ani10040642
Chicago/Turabian StyleGardela, Jaume, Annaïs Carbajal, Oriol Tallo-Parra, Sergi Olvera-Maneu, Manuel Álvarez-Rodríguez, Eduard Jose-Cunilleras, and Manel López-Béjar. 2020. "Temporary Relocation during Rest Periods: Relocation Stress and Other Factors Influence Hair Cortisol Concentrations in Horses" Animals 10, no. 4: 642. https://doi.org/10.3390/ani10040642
APA StyleGardela, J., Carbajal, A., Tallo-Parra, O., Olvera-Maneu, S., Álvarez-Rodríguez, M., Jose-Cunilleras, E., & López-Béjar, M. (2020). Temporary Relocation during Rest Periods: Relocation Stress and Other Factors Influence Hair Cortisol Concentrations in Horses. Animals, 10(4), 642. https://doi.org/10.3390/ani10040642