Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review


Abstract
Simple Summary
Abstract
1. Introduction
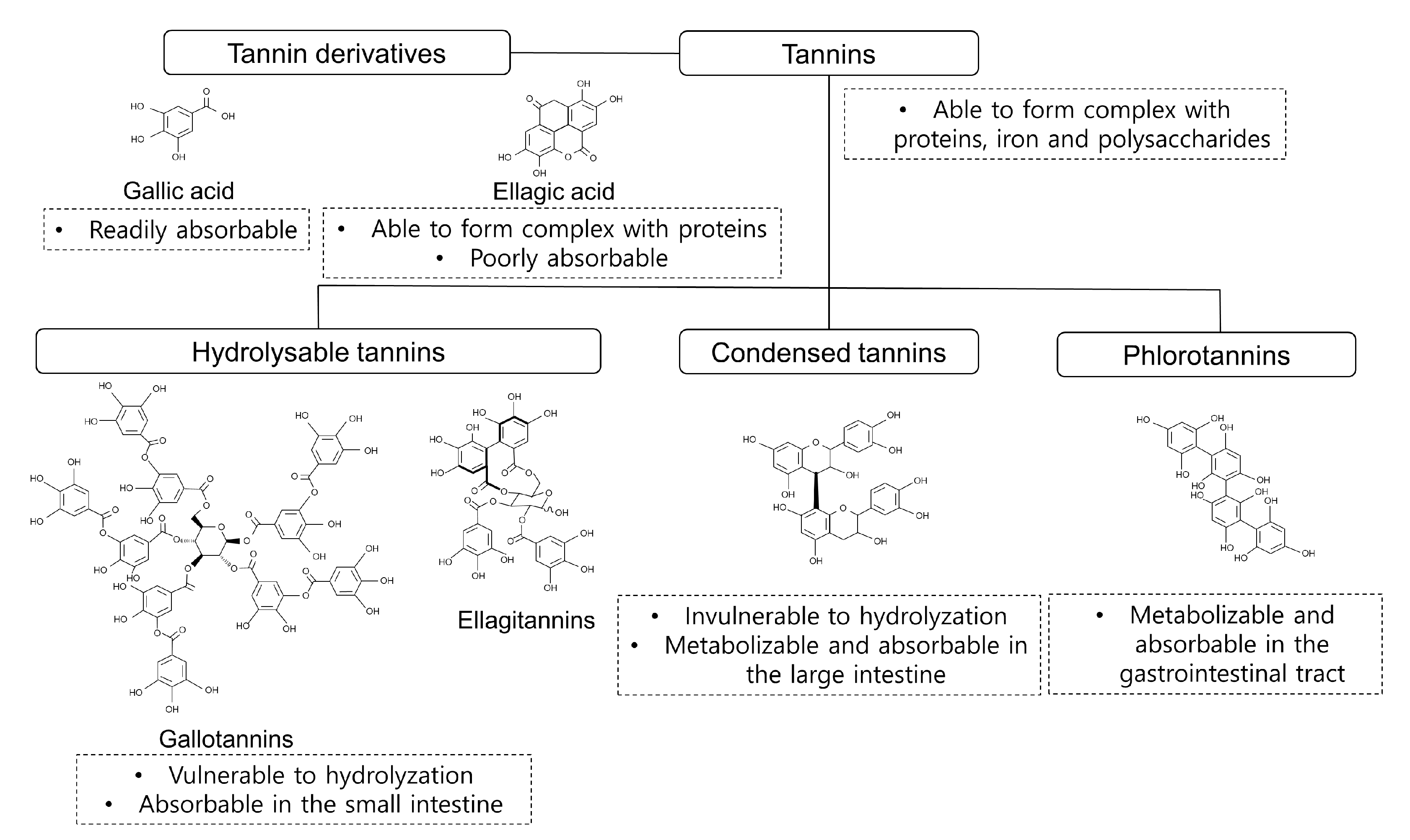
2. Classification and Bioavailability of Tannins
3. Traditional Viewpoints on Tannins as Antinutritional Factors in Poultry Production
4. Challenges in Poultry Production and Potential Solution by Using Tannins
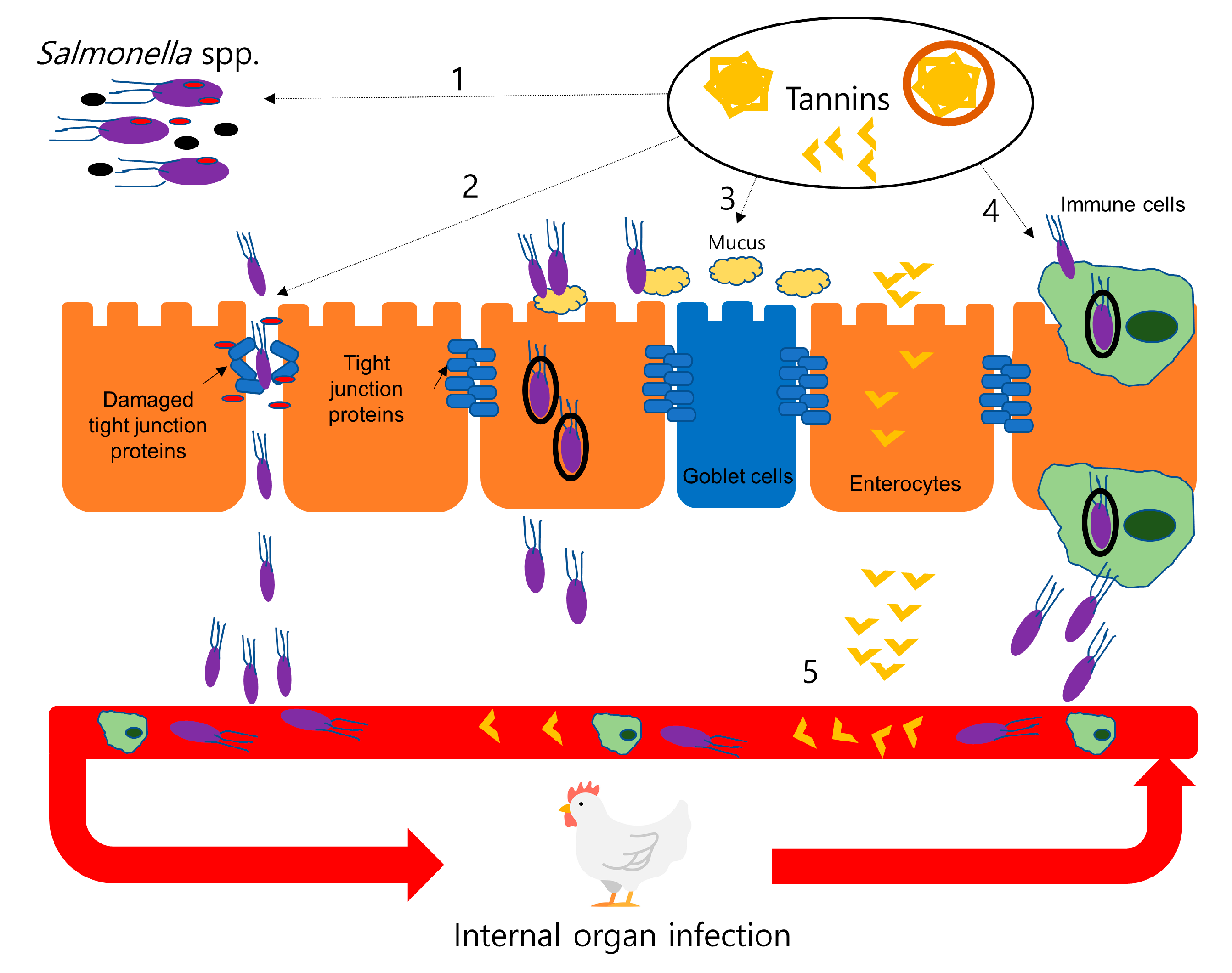
4.1. Effects of Tannins on Bacterial Infection (Salmonellosis)
4.2. Effects of Tannins on Coccidiosis
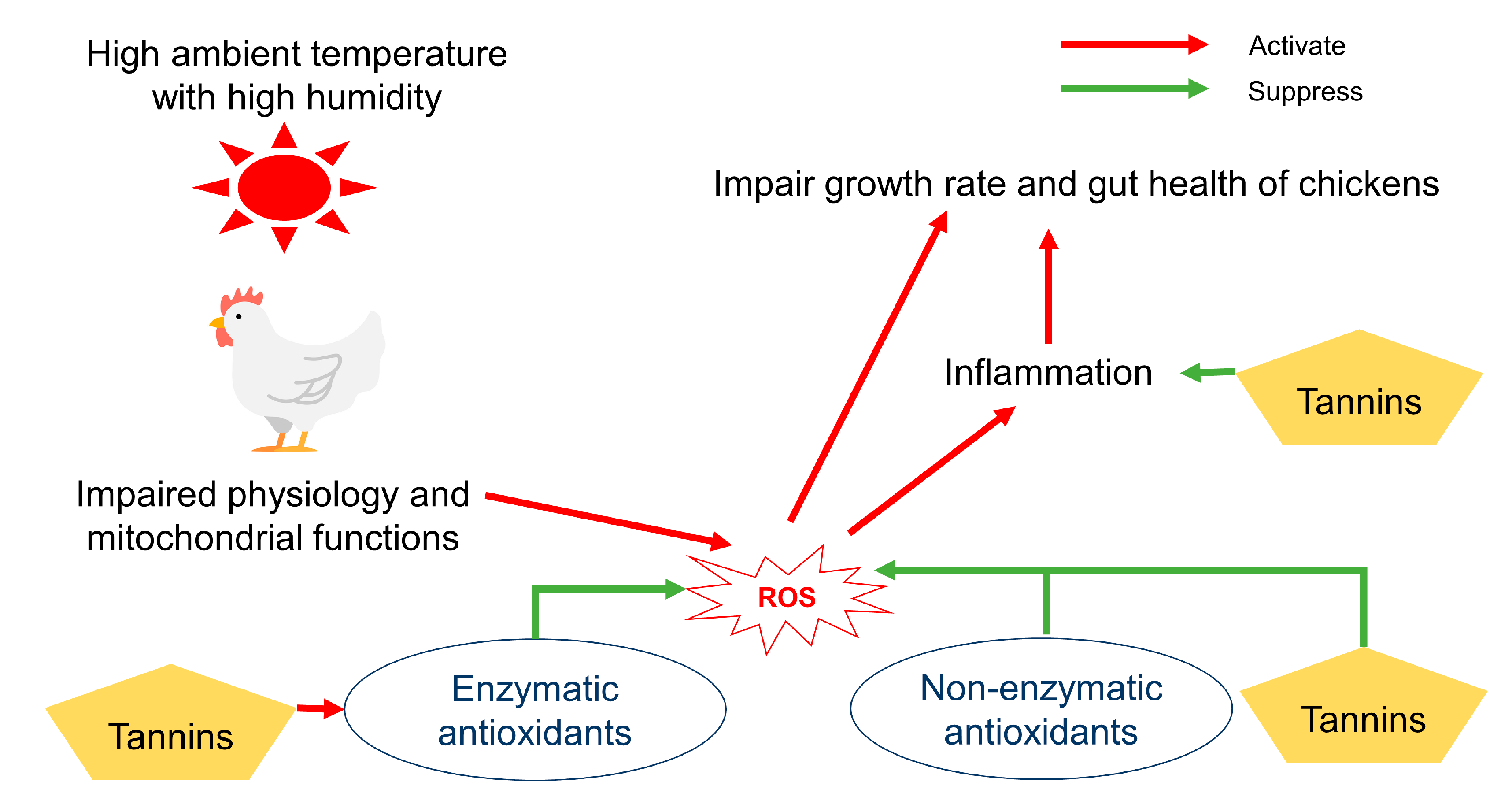
4.3. Effects of Tannins on Oxidative Stress, Including that Caused by Heat Stress
4.4. Effects of Tannins on Food Pad Dermatitis (FPD)
4.5. Effects of Tannins on Nitrogen Excretion and Emissions of Noxious and Greenhouse Gases
4.6. Effects of Tannins on Growth Performance, Immune System, Gut Microbiota, Gut Ecosystem and in Chickens Raised under General Conditions
5. Strategies to Maximize the Effects of Supplemental Tannins in the Chickens
5.1. Heat Process on Tannins
5.2. Co-Supplementation of Tannins with other Bioactive Compounds
5.3. Supplementation of Combined or Encapsulated Form of Tannins
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mottet, A.; Tempio, G. Global poultry production: Current state and future outlook and challenges. World. Poult. Sci. J. 2017, 73, 245–256. [Google Scholar] [CrossRef]
- Pawar, S.S.; Basavaraj, S.; Dhansing, L.V.; Kurade, N.P.; Sahebrao, K.A.; Vitthal, N.A.; Pandit, B.M.; Kumar, B.S. Assessing and Mitigating the Impact of Heat Stress in Poultry. Adv. Anim. Veter. Sci. 2016, 4, 332–341. [Google Scholar] [CrossRef]
- Nkukwana, T. Global poultry production: Current impact and future outlook on the South African poultry industry. South. Afr. J. Anim. Sci. 2019, 48, 869. [Google Scholar] [CrossRef]
- Akil, L.; Ahmad, H.A. Quantitative Risk Assessment Model of Human Salmonellosis Resulting from Consumption of Broiler Chicken. Diseases 2019, 7. [Google Scholar] [CrossRef]
- Hunter, J.M.; Anders, S.A.; Crowe, T.; Korver, D.R.; Bench, C.J. Practical assessment and management of foot pad dermatitis in commercial broiler chickens: A Field Study. J. Appl. Poult. Res. 2017, 26, 593–604. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Lahaye, L.; Gong, M.M.; Peng, J.; Gong, J.; Liu, S.; Gay, C.G.; Yang, C. Innovative drugs, chemicals, and enzymes within the animal production chain. Veter. Res. 2018, 49, 71. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Chen, C.; Indugu, N.; Werlang, G.O.; Singh, M.; Kim, W.K.; Thippareddi, H. Effect of antibiotic withdrawal in feed on chicken gut microbial dynamics, immunity, growth performance and prevalence of foodborne pathogens. PLoS ONE 2018, 13, e0192450. [Google Scholar] [CrossRef]
- Muhammad, J.; Khan, S.; Su, J.Q.; Hesham, A.E.-L.; Ditta, A.; Nawab, J.; Ali, A. Antibiotics in poultry manure and their associated health issues: A systematic review. J. Soils Sediments 2020, 20, 486–497. [Google Scholar] [CrossRef]
- Brown, K.; Uwiera, R.R.; Kalmokoff, M.L.; Brooks, S.P.; Inglis, G.D. Antimicrobial growth promoter use in livestock: A requirement to understand their modes of action to develop effective alternatives. Int. J. Antimicrob. Agents 2017, 49, 12–24. [Google Scholar] [CrossRef]
- Hu, Y.J.; Cowling, B.J. Reducing antibiotic use in livestock, China. Bull. World Health Organ. 2020, 98, 360. [Google Scholar] [CrossRef]
- Karavolias, J.; Salois, M.J.; Baker, K.T.; Watkins, K. Raised without antibiotics: impact on animal welfare and implications for food policy. Transl. Anim. Sci. 2018, 2, 337–348. [Google Scholar] [CrossRef]
- Singer, R.S.; Porter, L.J.; Thomson, D.U.; Gage, M.; Beaudoin, A.; Wishnie, J.K. Raising Animals Without Antibiotics: U.S. Producer and Veter.inarian Experiences and Opinions. Front. Veter. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Echeng, G.; Ehao, H.; Exie, S.; Ewang, X.; Edai, M.; Ehuang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Yang, C.; Chowdhury, M.A.K.; Hou, Y.; Gong, J. Phytogenic Compounds as Alternatives to In-Feed Antibiotics: Potentials and Challenges in Application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef]
- Redondo, L.M.; Chacana, P.A.; Dominguez, J.E.; Miyakawa, M.E.D.F. Perspectives in the use of tannins as alternative to antimicrobial growth promoter factors in poultry. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Trevino, J.; Ortiz, L.; Centeno, C. Effect of tannins from faba beans (Vicia faba) on the digestion of starch by growing chicks. Anim. Feed. Sci. Technol. 1992, 37, 345–349. [Google Scholar] [CrossRef]
- Garcia, R.; Mendes, A.; Sartori, J.; Paz, I.C.D.L.A.; Takahashi, S.; Pelícia, K.; Komiyama, C.; Quinteiro, R. Digestibility of feeds containing sorghum, with and without tannin, for broiler chickens submitted to three room temperatures. Brazil. J. Poultry Sci. 2004, 6, 55–60. [Google Scholar] [CrossRef]
- Brus, M.; Gradišnik, L.; Trapečar, M.; Škorjanc, D.; Frangež, R. Beneficial effects of water-soluble chestnut (Castanea sativa Mill.) tannin extract on chicken small intestinal epithelial cell culture. Poult. Sci. 2018, 97, 1271–1282. [Google Scholar] [CrossRef]
- Amirmohammadi, M.; Khajoenia, S.; Bahmani, M.; Rafieian-Kopaei, M.; Eftekhari, Z.; Qorbani, M. In vivo evaluation of antiparasitic effects of Artemisia abrotanum and Salvia officinalis extracts on Syphacia obvelata, Aspiculoris tetrapetra and Hymenolepis nana parasites. Asian Pac. J. Trop. Dis. 2014, 4, S250–S254. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Mannelli, F.; Team, H.; Tosi, G.; Secci, G.; Daghio, M.; Massi, P.; Fiorentini, L.; Galigani, I.; Lancini, S.; Rapaccini, S.; et al. Effect of Chestnut Tannins and Short Chain Fatty Acids as Anti-Microbials and as Feeding Supplements in Broilers Rearing and Meat Quality. Animals 2019, 9. [Google Scholar] [CrossRef]
- Haslam, E. Plant. Polyphenols: Vegetable Tannins Revisited. Australia (UK); Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef]
- Lamy, E.; Pinheiro, C.; Rodrigues, L.; Capela-Silva, F.; Lopes, O.; Tavares, S.; Gaspar, R. Determinants of Tannin-Rich Food and Beverage Consumption: Oral Perception vs. Psychosocial Aspects; Universidade De Évora: Évora, Portugal, 2016. [Google Scholar]
- Kamiloglu, S.; Capanoglu, E.; Bilen, F.D.; Gonzales, G.B.; Grootaert, C.; Van de Wiele, T.; Van Camp, J. Bioaccessibility of polyphenols from plant-processing byproducts of black carrot (Daucus carota L.). J. Agric. Food Chem. 2016, 64, 2450–2458. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Lee, R.; Heber, D. Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate (Punica granatum L.) juice. Clin. Chim. Acta 2004, 348, 63–68. [Google Scholar] [CrossRef]
- Martinez, K.B.; Mackert, J.D.; McIntosh, M.K. Polyphenols and intestinal health. In Nutrition and Functional Foods for Healthy Aging; Elsevier: Amsterdam, The Netherlands, 2017; pp. 191–210. [Google Scholar] [CrossRef]
- Selma, M.V.; Espin, J.C.; Tomas-Barberan, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Costain, D.J.; Guha, A.K.; Liwski, R.S.; Lee, T.D. Murine hypodense eosinophils induce tumour cell apoptosis by a granzyme B-dependent mechanism. Cancer Immunol. Immunother. 2001, 50, 293–299. [Google Scholar] [CrossRef]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53, S310–S329. [Google Scholar] [CrossRef]
- Kahle, K.; Huemmer, W.; Kempf, M.; Scheppach, W.; Erk, T.; Richling, E. Polyphenols are intensively metabolized in the human gastrointestinal tract after apple juice consumption. J. Agric. Food Chem. 2007, 55, 10605–10614. [Google Scholar] [CrossRef]
- Bawadi, H.; Bansode, R.; Trappey, A., II; Truax, R.; Losso, J. Inhibition of Caco-2 colon, MCF-7 and Hs578T breast, and DU 145 prostatic cancer cell proliferation by water-soluble black bean condensed tannins. Cancer Lett. 2005, 218, 153–162. [Google Scholar] [CrossRef]
- Ragan, M.; Glombitza, K. Handbook of Physiological Methods; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Kirke, D.; Smyth, T.; Rai, D.; Kenny, O.; Stengel, D. The chemical and antioxidant stability of isolated low molecular weight phlorotannins. Food Chem. 2017, 221, 1104–1112. [Google Scholar] [CrossRef]
- Corona, G.; Coman, M.M.; Guo, Y.; Hotchkiss, S.; Gill, C.; Yaqoob, P.; Spencer, J.P.; Rowland, I. Effect of simulated gastrointestinal digestion and fermentation on polyphenolic content and bioactivity of brown seaweed phlorotannin-rich extracts. Mol. Nutr. Food Res. 2017, 61, 1700223. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Barry, T.; Manley, T.; Duncan, S. The role of condensed tannins in the nutritional value of Lotus pedunculatus for sheep:* 4. Sites of carbohydrate and protein digestion as influenced by dietary reactive tannin concentration. Br. J. Nutr. 1986, 55, 123–137. [Google Scholar] [CrossRef]
- Berard, N.C.; Wang, Y.; Wittenberg, K.; Krause, D.; Coulman, B.; McAllister, T.; Ominski, K. Condensed tannin concentrations found in vegetative and mature forage legumes grown in western Canada. Can. J. Plant Sci. 2011, 91, 669–675. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P. Methane mitigation from ruminants using tannins and saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef]
- Medugu, C.; Saleh, B.; Igwebuike, J.; Ndirmbita, R. Strategies to improve the utilization of tannin-rich feed materials by poultry. Int. J. Poult. Sci. 2012, 11, 417. [Google Scholar] [CrossRef]
- Butler, L.G.; Riedl, D.J.; Lebryk, D.; Blytt, H. Interaction of proteins with sorghum tannin: Mechanism, specificity and significance. J. Am. Oil Chem. Soc. 1984, 61, 916–920. [Google Scholar] [CrossRef]
- Marzo, F.; Urdaneta, E.; Santidrian, S. Liver proteolytic activity in tannic acid-fed birds. Poult. Sci. 2002, 81, 92–94. [Google Scholar] [CrossRef]
- Lee, S.; Shinde, P.; Choi, J.; Kwon, I.; Lee, J.; Pak, S.; Cho, W.; Chae, B. Effects of tannic acid supplementation on growth performance, blood hematology, iron status and faecal microflora in weanling pigs. Livest. Sci. 2010, 131, 281–286. [Google Scholar] [CrossRef]
- Tonda, R.; Rubach, J.; Lumpkins, B.; Mathis, G.; Poss, M. Effects of tannic acid extract on performance and intestinal health of broiler chickens following coccidiosis vaccination and/or a mixed-species Eimeria challenge. Poult. Sci. 2018, 97, 3031–3042. [Google Scholar] [CrossRef]
- Cengiz, Ö.; Köksal, B.H.; Tatlı, O.; Sevim, Ö.; Ahsan, U.; Bilgili, S.F.; Önol, A.G. Effect of dietary tannic acid supplementation in corn-or barley-based diets on growth performance, intestinal viscosity, litter quality, and incidence and severity of footpad dermatitis in broiler chickens. Livest. Sci. 2017, 202, 52–57. [Google Scholar] [CrossRef]
- Starčević, K.; Krstulović, L.; Brozić, D.; Maurić, M.; Stojević, Z.; Mikulec, Ž.; Bajić, M.; Mašek, T. Production performance, meat composition and oxidative susceptibility in broiler chicken fed with different phenolic compounds. J. Sci. Food Agric. 2015, 95, 1172–1178. [Google Scholar] [CrossRef]
- Mašek, T.; Starčević, K.; Mikulec, Ž. The influence of the addition of thymol, tannic acid or gallic acid to broiler diet on growth performance, serum malondialdehyde value and cecal fermentation. Yeast 2014, 5, 5.0. [Google Scholar] [CrossRef]
- Kubena, L.; Byrd, J.; Young, C.; Corrier, D. Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks. Poult. Sci. 2001, 80, 1293–1298. [Google Scholar] [CrossRef]
- Ebrahim, R.; Liang, J.B.; Jahromi, M.F.; Shokryazdan, P.; Ebrahimi, M.; Li Chen, W.; Goh, Y.M. Effects of tannic acid on performance and fatty acid composition of breast muscle in broiler chickens under heat stress. Ital. J. Anim. Sci. 2015, 14, 3956. [Google Scholar] [CrossRef]
- Mor-Mur, M.; Yuste, J. Emerging bacterial pathogens in meat and poultry: An overview. Food Bioproc. Tech. 2010, 3, 24. [Google Scholar] [CrossRef]
- Adhikari, P.; Cosby, D.E.; Cox, N.A.; Franca, M.S.; Williams, S.M.; Gogal, R.M., Jr.; Ritz, C.W.; Kim, W.K. Effect of dietary fructooligosaccharide supplementation on internal organs Salmonella colonization, immune response, ileal morphology, and ileal immunohistochemistry in laying hens challenged with Salmonella enteritidis. Poult. Sci. 2018, 97, 2525–2533. [Google Scholar] [CrossRef]
- Thung, T.; Mahyudin, N.; Basri, D.; Radzi, C.W.M.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance of Salmonella Enteritidis and Salmonella Typhimurium in raw chicken meat at retail markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef]
- Wu-Yuan, C.; Chen, C.; Wu, R. Gallotannins inhibit growth, water-insoluble glucan synthesis, and aggregation of mutans streptococci. J. Dent. Res. 1988, 67, 51–55. [Google Scholar] [CrossRef]
- Delimont, N.M.; Rosenkranz, S.K.; Haub, M.D.; Lindshield, B.L. Salivary proline-rich protein may reduce tannin-iron chelation: A systematic narrative review. Nutr. Metab. 2017, 14, 47. [Google Scholar] [CrossRef]
- Karamać, M. Chelation of Cu (II), Zn (II), and Fe (II) by tannin constituents of selected edible nuts. Int. J. Mol. Sci. 2009, 10, 5485–5497. [Google Scholar] [CrossRef]
- Tan, Z. Roles of Iron Uptake in the Survival, Colonization and Virulence of Salmonella in Animal Guts. Master’s Thesis, University of Manitoba, Winnipeg, MB, Canada, 2019. [Google Scholar]
- Mahadwar, G.; Chauhan, K.; Bhagavathy, G.; Murphy, C.; Smith, A.; Bhagwat, A. Swarm motility of S almonella enterica serovar T yphimurium is inhibited by compounds from fruit peel extracts. Lett. Appl. Microbiol. 2015, 60, 334–340. [Google Scholar] [CrossRef]
- Sivasankar, C.; Jha, N.K.; Ghosh, R.; Shetty, P.H. Anti quorum sensing and anti virulence activity of tannic acid and it’s potential to breach resistance in Salmonella enterica Typhi/Paratyphi A clinical isolates. Microb. Pathog. 2020, 138, 103813. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, L.; Gao, J.; Liu, X.; Feng, Y.; Wu, Q.; Baloch, A.B.; Cui, L.; Xia, X. Tannin-rich fraction from pomegranate rind inhibits quorum sensing in Chromobacterium violaceum and biofilm formation in Escherichia coli. Foodborne Pathog. Dis. 2016, 13, 28–35. [Google Scholar] [CrossRef]
- Costabile, A.; Sanghi, S.; Martín-Pelaez, S.; Mueller-Harvey, I.; Gibson, G.R.; Rastall, R.A.; Klinder, A. Inhibition of Salmonella Typhimurium by tannins in vitro. J. Food. Agric. Environ. 2011, 9, 119–124. [Google Scholar]
- Reyes, A.W.B.; Hong, T.G.; Hop, H.T.; Arayan, L.T.; Huy, T.X.N.; Min, W.; Lee, H.J.; Lee, K.S.; Kim, S. The in vitro and in vivo protective effects of tannin derivatives against Salmonella enterica serovar Typhimurium infection. Microb. Pathog. 2017, 109, 86–93. [Google Scholar] [CrossRef]
- Daing, M.I.; Pathak, A.; Bhat, M.A.; Zargar, M.A. Antioxidant and antibacterial potential of condensed tannins containing tree leaves extract. Vet. Pract. 2017, 18, 118–121. [Google Scholar]
- Maisetta, G.; Batoni, G.; Caboni, P.; Esin, S.; Rinaldi, A.C.; Zucca, P. Tannin profile, antioxidant properties, and antimicrobial activity of extracts from two Mediterranean species of parasitic plant Cytinus. BMC Complement Altern. Med. 2019, 19, 82. [Google Scholar] [CrossRef]
- Elizondo, A.M.; Mercado, E.C.; Rabinovitz, B.C.; Fernandez-Miyakawa, M.E. Effect of tannins on the in vitro growth of Clostridium perfringens. Vet. Microbiol. 2010, 145, 308–314. [Google Scholar] [CrossRef]
- Liu, H.; Li, K.; Zhao, J.; Deng, W. Effects of chestnut tannins on intestinal morphology, barrier function, pro-inflammatory cytokine expression, microflora and antioxidant capacity in heat-stressed broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 717–726. [Google Scholar] [CrossRef]
- Kamali Sangani, A.; Masoudi, A.A.; Hosseini, S.A. The effects of herbal plants on Mucin 2 gene expression and performance in ascetic broilers. Iran. J. Vet. Med. 2014, 8, 47–52. [Google Scholar] [CrossRef]
- Moghaddam, H.S.; Moghaddam, H.N.; Kermanshahi, H.; Mosavi, A.H.; Raji, A. The effect of threonine on mucin2 gene expression, intestinal histology and performance of broiler chicken. Ital. J. Anim. Sci. 2011, 10, e14. [Google Scholar] [CrossRef]
- Urdaneta, V.; Casadesús, J. Interactions between bacteria and bile salts in the gastrointestinal and hepatobiliary tracts. Front. Med. 2017, 4, 163. [Google Scholar] [CrossRef]
- Li, X.; Bleumink-Pluym, N.M.; Luijkx, Y.M.; Wubbolts, R.W.; van Putten, J.P.; Strijbis, K. MUC1 is a receptor for the Salmonella SiiE adhesin that enables apical invasion into enterocytes. PLoS Pathog. 2019, 15, e1007566. [Google Scholar] [CrossRef]
- Van Parys, A.; Boyen, F.; Dewulf, J.; Haesebrouck, F.; Pasmans, F. The use of tannins to control Salmonella typhimurium infections in pigs. Zoonoses Public Health 2010, 57, 423–428. [Google Scholar] [CrossRef]
- Jamroz, D.; Wiliczkiewicz, A.; Skorupińska, J.; Orda, J.; Kuryszko, J.; Tschirch, H. Effect of sweet chestnut tannin (SCT) on the performance, microbial status of intestine and histological characteristics of intestine wall in chickens. Br. Poult. Sci. 2009, 50, 687–699. [Google Scholar] [CrossRef]
- Adhikari, P.; Cosby, D.; Cox, N.; Kim, W. Colonization of mature laying hens with Salmonella Enteritidis by oral or intracloacal inoculation. J. Appl. Poult. Res. 2017, 26, 286–294. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Cheng, Y.; Li, Y.; Wen, C.; Zhou, Y. Dietary l-threonine supplementation attenuates lipopolysaccharide-induced inflammatory responses and intestinal barrier damage of broiler chickens at an early age. Br. J. Nutr. 2018, 119, 1254–1262. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Liu, S.; Hui, Q.; Zhang, H.; Lahaye, L.; Bodin, J.-C.; Gong, J.; Nyachoti, M.; Yang, C. Thymol improves barrier function and attenuates inflammatory responses in porcine intestinal epithelial cells during lipopolysaccharide (LPS)-induced inflammation. J. Agric. Food Chem. 2018, 67, 615–624. [Google Scholar] [CrossRef]
- van Ampting, M.T.; Schonewille, A.J.; Vink, C.; Brummer, R.J.M.; van der Meer, R.; Bovee-Oudenhoven, I.M. Damage to the intestinal epithelial barrier by antibiotic pretreatment of salmonella-infected rats is lessened by dietary calcium or tannic acid. J. Nutr. 2010, 140, 2167–2172. [Google Scholar] [CrossRef]
- Adhikari, P.; Lee, C.; Cosby, D.; Cox, N.; Kim, W. Effect of probiotics on fecal excretion, colonization in internal organs and immune gene expression in the ileum of laying hens challenged with SalmonellaEnteritidis. Poult. Sci. 2019, 98, 1235–1242. [Google Scholar] [CrossRef]
- Sieniawska, E.; Baj, T. Tannins. In Pharmacognosy; Elsevier: Amsterdam, The Netherlands, 2017; pp. 199–232. [Google Scholar]
- Konishi, Y.; Kobayashi, S.; Shimizu, M. Transepithelial transport of p-coumaric acid and gallic acid in Caco-2 cell monolayers. Biosci. Biotechnol. Biochem. 2003, 67, 2317–2324. [Google Scholar] [CrossRef]
- Grilli, G.; Borgonovo, F.; Tullo, E.; Fontana, I.; Guarino, M.; Ferrante, V. A pilot study to detect coccidiosis in poultry farms at early stage from air analysis. Biosyst. Eng. 2018, 173, 64–70. [Google Scholar] [CrossRef]
- Chapman, H. Milestones in avian coccidiosis research: A review. Poult. Sci. 2014, 93, 501–511. [Google Scholar] [CrossRef]
- Clark, E.L.; Macdonald, S.E.; Thenmozhi, V.; Kundu, K.; Garg, R.; Kumar, S.; Ayoade, S.; Fornace, K.M.; Jatau, I.D.; Moftah, A. Cryptic Eimeria genotypes are common across the southern but not northern hemisphere. Int. J. Parasitol. 2016, 46, 537–544. [Google Scholar] [CrossRef]
- Li, C.; Yan, X.; Lillehoj, H.S.; Oh, S.; Liu, L.; Sun, Z.; Gu, C.; Lee, Y.; Xianyu, Z.; Zhao, H. Eimeria maxima-induced transcriptional changes in the cecal mucosa of broiler chickens. Parasites Vectors 2019, 12, 285. [Google Scholar] [CrossRef]
- Yin, G.; Lin, Q.; Wei, W.; Qin, M.; Liu, X.; Suo, X.; Huang, Z. Protective immunity against Eimeria tenella infection in chickens induced by immunization with a recombinant C-terminal derivative of EtIMP1. Vet. Immunol. Immunopathol. 2014, 162, 117–121. [Google Scholar] [CrossRef]
- Teng, P.-Y.; Yadav, S.; de Souza Castro, F.L.; Tompkins, Y.H.; Fuller, A.L.; Kim, W.K. Graded Eimeria challenge linearly regulated growth performance, dynamic change of gastrointestinal permeability, apparent ileal digestibility, intestinal morphology, and tight junctions of broiler chickens. Poult. Sci. 2020, 99, 4203–4216. [Google Scholar] [CrossRef]
- Abbas, R.; Iqbal, Z.; Mansoor, M. Role of natural antioxidants for the control of coccidiosis in poultry. Pak. Vet. J. 2013, 33, 401. [Google Scholar]
- Prescott, J.F.; Smyth, J.A.; Shojadoost, B.; Vince, A. Experimental reproduction of necrotic enteritis in chickens: A review. Avian Dis. 2016, 45, 317–322. [Google Scholar] [CrossRef]
- Wu, S.-B.; Stanley, D.; Rodgers, N.; Swick, R.A.; Moore, R.J. Two necrotic enteritis predisposing factors, dietary fishmeal and Eimeria infection, induce large changes in the caecal microbiota of broiler chickens. Vet. Microbiol. 2014, 169, 188–197. [Google Scholar] [CrossRef]
- Peek, H.; Landman, W. Coccidiosis in poultry: Anticoccidial products, vaccines and other prevention strategies. Vet. Q. 2011, 31, 143–161. [Google Scholar] [CrossRef]
- Noack, S.; Chapman, H.D.; Selzer, P.M. Anticoccidial drugs of the livestock industry. Parasitol. Res. 2019, 118, 2009–2026. [Google Scholar] [CrossRef]
- Levine, R.; Horst, G.; Tonda, R.; Lumpkins, B.; Mathis, G. Evaluation of the effects of feeding dried algae containing beta-1, 3-glucan on broilers challenged with Eimeria. Poult. Sci. 2018, 97, 3494–3500. [Google Scholar] [CrossRef]
- Christaki, E.; Florou-Paneri, P.; Giannenas, I.; Papazahariadou, M.; Botsoglou, N.A.; Spais, A.B. Effect of a mixture of herbal extracts on broiler chickens infected with Eimeria tenella. Anim. Res. 2004, 53, 137–144. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Munawar, S.H.; Manzoor, Z.; Iqbal, Z.; Khan, M.N.; Saleemi, M.K.; Zia, M.A.; Yousaf, A. Anticoccidial effects of acetic acid on performance and pathogenic parameters in broiler chickens challenged with Eimeria tenella. Pesqui. Vet. Bras. 2011, 31, 99–103. [Google Scholar] [CrossRef]
- Yang, C.; Kennes, Y.M.; Lepp, D.; Yin, X.; Wang, Q.; Yu, H.; Yang, C.; Gong, J.; Diarra, M.S. Effects of encapsulated cinnamaldehyde and citral on the performance and cecal microbiota of broilers vaccinated or not vaccinated against coccidiosis. Poult. Sci. 2020, 99, 936–948. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Rothrock, M.J.; Vieira, B.S.; Mallo, J.J.; Puyalto, M.; Hofacre, C.; Applegate, T.J. Supplementation of protected sodium butyrate alone or in combination with essential oils modulated the cecal microbiota of broiler chickens challenged with coccidia and Clostridium perfringens. Front. Sustain. Food Syst. 2018, 2, 72. [Google Scholar] [CrossRef]
- He, B.; Bortoluzzi, C.; King, W.; Graugnard, D.; Dawson, K.; Applegate, T. Zinc source influences the gene expression of zinc transporters in jejunum and cecal tonsils during broiler challenge with Eimeria maximaand Clostridium perfringens. Poult. Sci. 2019, 98, 1146–1152. [Google Scholar] [CrossRef]
- Teng, P.-Y.; Fuller, A.L.; Kim, W.K. Evaluation of nitro compounds as feed additives in diets of Eimeria-challenged broilers in vitro and in vivo. Poult. Sci. 2020, 99, 1320–1325. [Google Scholar] [CrossRef]
- Min, B.; Hart, S. Tannins for suppression of internal parasites. J. Anim. Sci. 2003, 81, E102–E109. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Chung, K.-T.; Lu, Z.; Chou, M. Mechanism of inhibition of tannic acid and related compounds on the growth of intestinal bacteria. Food Chem. Toxicol. 1998, 36, 1053–1060. [Google Scholar] [CrossRef]
- Parisi, F.; Mancini, S.; Mazzei, M.; Forzan, M.; Turchi, B.; Perrucci, S.; Poli, A.; Paci, G. Effect of dietary supplementation of a mix of chestnut and quebracho tannins on intestinal morphology, bacterial load, eimeria spp oocyst excretion and immune response after vaccination in rabbits. Am. J. Anim.Vet. Sci. 2018. [Google Scholar] [CrossRef]
- Kaleem, Q.M.; Akhtar, M.; Awais, M.M.; Saleem, M.; Zafar, M.; Iqbal, Z.; Muhammad, F.; Anwar, M.I. Studies on Emblica officinalis derived tannins for their immunostimulatory and protective activities against coccidiosis in industrial broiler chickens. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef]
- Wang, M.; Suo, X.; Gu, J.; Zhang, W.; Fang, Q.; Wang, X. Influence of grape seed proanthocyanidin extract in broiler chickens: Effect on chicken coccidiosis and antioxidant status. Poult. Sci. 2008, 87, 2273–2280. [Google Scholar] [CrossRef]
- Mansoori, B.; Modirsanei, M. Effects of dietary tannic acid and vaccination on the course of coccidiosis in experimentally challenged broiler chicken. Vet. Parasitol. 2012, 187, 119–122. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; El-Shafey, A.S.; Rehab, Y.A.; Kim, W.K. Enhancing tolerance of broiler chickens to heat stress by supplementation with vitamin E, vitamin C and/or probiotics. Ann. Anim. Sci. 2017, 17, 1155–1169. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.; Sakai, M.; Sá, L.; Ferreira, A.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef]
- Laudicina, D.C.; Marnett, L.J. Enhancement of hydroperoxide-dependent lipid peroxidation in rat liver microsomes by ascorbic acid. Arch. Biochem. Biophys. 1990, 278, 73–80. [Google Scholar] [CrossRef]
- Choi, J.; Li, W.; Schindell, B.; Ni, L.; Liu, S.; Zhao, X.; Gong, J.; Nyachoti, M.; Yang, C. Molecular cloning, tissue distribution and the expression of cystine/glutamate exchanger (xCT, SLC7A11) in different tissues during development in broiler chickens. Anim. Nutr. 2020, 6, 107–114. [Google Scholar] [CrossRef]
- Lin, H.; Decuypere, E.; Buyse, J. Acute heat stress induces oxidative stress in broiler chickens. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 144, 11–17. [Google Scholar] [CrossRef]
- Lee, S.; Kim, D.; Keum, M.; Han, E.; An, B.; Chang, H.; Choi, Y.; Moon, B.; Lee, K. Effects of fumonisin B1 and mycotoxin binders on growth performance, tibia characteristics, gut physiology, and stress indicators in broiler chickens raised in different stocking densities. Poult. Sci. 2018, 97, 845–854. [Google Scholar] [CrossRef]
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246. [Google Scholar] [CrossRef]
- Nawab, A.; Ibtisham, F.; Li, G.; Kieser, B.; Wu, J.; Liu, W.; Zhao, Y.; Nawab, Y.; Li, K.; Xiao, M. Heat stress in poultry production: Mitigation strategies to overcome the future challenges facing the global poultry industry. J. Therm. Biol. 2018, 78, 131–139. [Google Scholar] [CrossRef]
- Yang, R.; Hui, Q.; Jiang, Q.; Liu, S.; Zhang, H.; Wu, J.; Lin, F.; Yang, C. Effect of Manitoba-grown red-osier dogwood extracts on recovering Caco-2 cells from H2O2-induced oxidative damage. Antioxidants 2019, 8, 250. [Google Scholar] [CrossRef]
- Jiang, Q.; Zhang, H.; Yang, R.; Hui, Q.; Chen, Y.; Mats, L.; Tsao, R.; Yang, C. Red-Osier Dogwood Extracts Prevent Inflammatory Responses in Caco-2 Cells and a Caco-2 BBe1/EA. hy926 Cell Co-Culture Model. Antioxidants 2019, 8, 428. [Google Scholar] [CrossRef]
- Terruzzi, I.; Montesano, A.; Senesi, P.; Villa, I.; Ferraretto, A.; Bottani, M.; Vacante, F.; Spinello, A.; Bolamperti, S.; Luzi, L. L-Carnitine reduces oxidative stress and promotes cells differentiation and bone matrix proteins expression in human osteoblast-like cells. Biomed Res. Int. 2019, 2019. [Google Scholar] [CrossRef]
- Sahin, K.; Sahin, N.; Kucuk, O. Effects of chromium, and ascorbic acid supplementation on growth, carcass traits, serum metabolites, and antioxidant status of broiler chickens reared at a high ambient temperature (32 C). Nutr. Res. 2003, 23, 225–238. [Google Scholar] [CrossRef]
- Yun, S.-H.; Moon, Y.-S.; SoHn, S.-H.; Jang, I.-S. Effects of cyclic heat stress or vitamin C supplementation during cyclic heat stress on HSP70, inflammatory cytokines, and the antioxidant defense system in Sprague Dawley rats. Exp. Anim. 2012, 61, 543–553. [Google Scholar] [CrossRef]
- Kumbhar, S.; Khan, A.Z.; Parveen, F.; Nizamani, Z.A.; Siyal, F.A.; Abd El-Hack, M.E.; Gan, F.; Liu, Y.; Hamid, M.; Nido, S.A. Impacts of selenium and vitamin E supplementation on mRNA of heat shock proteins, selenoproteins and antioxidants in broilers exposed to high temperature. Amb Express 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.; Warner, R.D.; Dunshea, F.R. Growth performance and characterization of meat quality of broiler chickens supplemented with betaine and antioxidants under cyclic heat stress. Antioxidants 2019, 8, 336. [Google Scholar] [CrossRef]
- Shi, J.; Yu, J.; Pohorly, J.E.; Kakuda, Y. Polyphenolics in grape seeds—biochemistry and functionality. J. Med. Food. 2003, 6, 291–299. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Ali, S.; Sahin, N.; Hayirli, A. Epigallocatechin-3-gallate prevents lipid peroxidation and enhances antioxidant defense system via modulating hepatic nuclear transcription factors in heat-stressed quails. Poult. Sci. 2010, 89, 2251–2258. [Google Scholar] [CrossRef]
- Ramnath, V.; Rekha, P. Brahma Rasayana enhances in vivo antioxidant status in cold-stressed chickens (Gallus gallus domesticus). Indian J. Pharmacol. 2009, 41, 115. [Google Scholar] [CrossRef]
- Hosseini-Vashan, S.J.; Safdari-Rostamabad, M.; Piray, A.H.; Sarir, H. The growth performance, plasma biochemistry indices, immune system, antioxidant status, and intestinal morphology of heat-stressed broiler chickens fed grape (Vitis vinifera) pomace. Anim. Feed Sci. Technol. 2020, 259, 114343. [Google Scholar] [CrossRef]
- Mujahid, A.; Yoshiki, Y.; Akiba, Y.; Toyomizu, M. Superoxide radical production in chicken skeletal muscle induced by acute heat stress. Poult. Sci. 2005, 84, 307–314. [Google Scholar] [CrossRef]
- Nakagawa, T.; Yokozawa, T. Direct scavenging of nitric oxide and superoxide by green tea. Food Chem. Toxicol. 2002, 40, 1745–1750. [Google Scholar] [CrossRef]
- Shepherd, E.; Fairchild, B. Footpad dermatitis in poultry. Poult. Sci. 2010, 89, 2043–2051. [Google Scholar] [CrossRef]
- Abd El-Wahab, A.; Radko, D.; Kamphues, J. High dietary levels of biotin and zinc to improve health of foot pads in broilers exposed experimentally to litter with critical moisture content. Poult. Sci. 2013, 92, 1774–1782. [Google Scholar] [CrossRef]
- Thøfner, I.C.N.; Poulsen, L.L.; Bisgaard, M.; Christensen, H.; Olsen, R.H.; Christensen, J.P. Correlation between footpad lesions and systemic bacterial infections in broiler breeders. Vet. Res. 2019, 50, 38. [Google Scholar] [CrossRef] [PubMed]
- Heidemann Olsen, R.; Christensen, H.; Kabell, S.; Bisgaard, M. Characterization of prevalent bacterial pathogens associated with pododermatitis in table egg layers. Avian Dis. 2018, 47, 281–285. [Google Scholar] [CrossRef] [PubMed]
- El-Wahab, A.A.; Visscher, C.; Beineke, A.; Beyerbach, M.; Kamphues, J. Experimental studies on the effects of different litter moisture contents and exposure time to wet litter on development and severity of foot pad dermatitis in young fattening turkeys. Arch. Geflugelkd. 2012, 76, 55–62. [Google Scholar]
- Mayne, R.; Else, R.; Hocking, P. High litter moisture alone is sufficient to cause footpad dermatitis in growing turkeys. Br. Poult. Sci. 2007, 48, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Cengız, Ö.; Hess, J.; Bilgili, S. Feed enzyme supplementation does not ameliorate foot pad dermatitis in broiler chickens fed on a corn-soyabean diet. Br. Poult. Sci. 2012, 53, 401–407. [Google Scholar] [CrossRef]
- Rezar, V.; Salobir, J. Effects of tannin-rich sweet chestnut (Castanea sativa mill.) wood extract supplementation on nutrient utilisation and excreta dry matter content in broiler chickens. Eur. Poult. Sci. 2014, 78, 1–10. [Google Scholar] [CrossRef]
- Salmond, G.G. Supplementation of Chelated Zn Cu and Mn in Broiler Diets to Improve Performance Skin Integrity and the Incidence and Severity of Footpad Dermatitis. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2015. [Google Scholar]
- Mayne, R.; Powell, F.; Else, R.; Kaiser, P.; Hocking, P. Foot pad dermatitis in growing turkeys is associated with cytokine and cellular changes indicative of an inflammatory immune response. Avian Dis. 2007, 36, 453–459. [Google Scholar] [CrossRef][Green Version]
- Bostami, A.; Ahmed, S.; Islam, M.; Mun, H.; Ko, S.; Kim, S.; Yang, C. Growth performance, fecal noxious gas emission and economic efficacy in broilers fed fermented pomegranate byproducts as residue of fruit industry. Int. J. Adv. Res 2015, 3, 102–114. [Google Scholar]
- Wang, S.-Y.; Huang, D.-J. Assessment of greenhouse gas emissions from poultry enteric fermentation. Asian-Australas J. Anim. Sci. 2005, 18, 873–878. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, M.; Meng, Q. Effects of atmospheric hydrogen sulfide concentration on growth and meat quality in broiler chickens. Poult. Sci. 2011, 90, 2409–2414. [Google Scholar] [CrossRef]
- McCrory, D.; Hobbs, P. Additives to reduce ammonia and odor emissions from livestock wastes: A review. J. Environ. Qual. 2001, 30, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Aneja, V.P.; Schlesinger, W.H.; Erisman, J.W.; Behera, S.N.; Sharma, M.; Battye, W. Reactive nitrogen emissions from crop and livestock farming in India. Atmos. Environ. 2012, 47, 92–103. [Google Scholar] [CrossRef]
- Xin, H.; Gates, R.S.; Green, A.R.; Mitloehner, F.M.; Moore Jr, P.A.; Wathes, C.M. Environmental impacts and sustainability of egg production systems. Poult. Sci. 2011, 90, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Malomo, G.A.; Bolu, S.A.; Madugu, A.S.; Usman, Z.S. Nitrogen Emissions and Mitigation Strategies in Chicken Production. Anim. Husband. Nutrit 2018, 43, 43–62. [Google Scholar]
- Aboagye, I.A.; Oba, M.; Koenig, K.M.; Zhao, G.Y.; Beauchemin, K.A. Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage. J. Anim. Sci. 2019, 97, 2230–2244. [Google Scholar] [CrossRef]
- Ahmed, S.T.; Yang, C.-J. Effects of dietary Punica granatum L. by-products on performance, immunity, intestinal and fecal microbiology, and odorous gas emissions from excreta in broilers. J. Poult. Sci. 2017. [Google Scholar] [CrossRef]
- Ramah, A.; Yasuda, M.; Ohashi, Y.; Urakawa, M.; Kida, T.; Yanagita, T.; Uemura, R.; Bakry, H.H.; Abdelaleem, N.M.; El-Shewy, E.A. Different doses of tannin reflect a double-edged impact on broiler chicken immunity. Vet. Immunol. Immunopathol. 2020, 220, 109991. [Google Scholar] [CrossRef]
- Karaffová, V.; Bobíková, K.; Levkut, M.; Revajová, V.; Ševčíková, Z.; Levkut, M. The influence of Farmatan® and Flimabend® on the mucosal immunity of broiler chicken. Poult. Sci. 2019, 98, 1161–1166. [Google Scholar] [CrossRef]
- Erlejman, A.G.; Jaggers, G.; Fraga, C.G.; Oteiza, P.I. TNFα-induced NF-κB activation and cell oxidant production are modulated by hexameric procyanidins in Caco-2 cells. Arch. Biochem. Biophys. 2008, 476, 186–195. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Redondo, E.A.; Pin Viso, N.D.; Redondo, L.M.; Farber, M.D.; Fernandez Miyakawa, M.E. Tannins and bacitracin differentially modulate gut microbiota of broiler chickens. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Koo, B.; Nyachoti, C.M. Effects of thermally oxidized canola oil and tannic acid supplementation on nutrient digestibility and microbial metabolites in finishing pigs. J. Anim. Sci. 2019, 97, 2468–2478. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Ordaz, R.; Wall-Medrano, A.; Goñi, M.G.; Ramos-Clamont-Montfort, G.; Ayala-Zavala, J.F.; González-Aguilar, G. Effect of phenolic compounds on the growth of selected probiotic and pathogenic bacteria. Lett. Appl. Microbiol. 2018, 66, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Viveros, A.; Chamorro, S.; Pizarro, M.; Arija, I.; Centeno, C.; Brenes, A. Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 2011, 90, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef]
- Bilić-Šobot, D.; Kubale, V.; Škrlep, M.; Čandek-Potokar, M.; Prevolnik Povše, M.; Fazarinc, G.; Škorjanc, D. Effect of hydrolysable tannins on intestinal morphology, proliferation and apoptosis in entire male pigs. Arch. Anim. Nutr. 2016, 70, 378–388. [Google Scholar] [CrossRef]
- Kim, T.; Silva, J.; Kim, M.; Jung, Y. Enhanced antioxidant capacity and antimicrobial activity of tannic acid by thermal processing. Food Chem. 2010, 118, 740–746. [Google Scholar] [CrossRef]
- Adhikari, P.A.; Kim, W.K. Overview of prebiotics and probiotics: Focus on performance, gut health and immunity–a review. Ann. Anim. Sci. 2017, 17, 949–966. [Google Scholar] [CrossRef]
- Huyghebaert, G.; Ducatelle, R.; Van Immerseel, F. An update on alternatives to antimicrobial growth promoters for broilers. Vet. J. 2011, 187, 182–188. [Google Scholar] [CrossRef]
- Khalil, R.K. Influence of gallic acid and catechin polyphenols on probiotic properties of Streptococcus thermophilus CHCC 3534 strain. World J. Microbiol. Biotechnol. 2010, 26, 2069–2079. [Google Scholar] [CrossRef]
- Jarquin, R.; Nava, G.; Wolfenden, A.; Donoghue, A.; Hanning, I.; Higgins, S.; Hargis, B. The evaluation of organic acids and probiotic cultures to reduce Salmonella enteriditis horizontal transmission and crop infection in broiler chickens. Int. J. Poult. Sci 2007, 6, 182–186. [Google Scholar] [CrossRef][Green Version]
- Al-Harbi, R.; Shaaban, M.; Al-Wegaisi, R.; Moharram, F.; El-Rahman, O.A.; El-Messery, S. Antimicrobial Activity and Molecular Docking of Tannins from Pimenta dioica. Lett. Drug Des. Discov. 2018, 15, 508–515. [Google Scholar] [CrossRef]
- Khan, S.H.; Iqbal, J. Recent advances in the role of organic acids in poultry nutrition. J. Appl. Anim. Res. 2016, 44, 359–369. [Google Scholar] [CrossRef]
- De Lange, C.; Pluske, J.; Gong, J.; Nyachoti, C. Strategic use of feed ingredients and feed additives to stimulate gut health and development in young pigs. Livest. Sci. 2010, 134, 124–134. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Yu, B.; Chen, D.; Mao, X.; Zheng, P.; Luo, J.; He, J. Dietary chlorogenic acid improves growth performance of weaned pigs through maintaining antioxidant capacity and intestinal digestion and absorption function. J. Anim. Sci. 2018, 96, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Wang, L.; Liu, S.; Lu, P.; Zhao, X.; Liu, H.; Lahaye, L.; Santin, E.; Liu, S.; Nyachoti, M. Effects of a microencapsulated formula of organic acids and essential oils on nutrient absorption, immunity, gut barrier function, and abundance of enterotoxigenic Escherichia coli F4 in weaned piglets challenged with E. coli F4. J. Anim. Sci. 2020, 98, skaa259. [Google Scholar] [CrossRef] [PubMed]
- Gracia, L.G.; de Castro, M.L. Development and validation of a flow-injection method for the determination of albumin tannate, the active component of a pharmaceutical preparation. J. Pharm. Biomed. Anal. 1997, 15, 447–452. [Google Scholar] [CrossRef]
- Choi, J.; Wang, L.; Ammeter, E.; Lahaye, L.; Liu, S.; Nyachoti, M.; Yang, C. Evaluation of lipid matrix microencapsulation for intestinal delivery of thymol in weaned pigs. Transl. Anim. Sci. 2020, 4, 411–422. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Kim, S.; Guo, T.; Wang, Q.; Gong, J.; Lahaye, L.; Bodin, J.-C.; Nyachoti, M.; Liu, S.; Yang, C. Development of novel microparticles for effective delivery of thymol and lauric acid to pig intestinal tract. J. Agric. Food Chem. 2018, 66, 9608–9615. [Google Scholar] [CrossRef]
- Liu, H.; Gong, J.; Chabot, D.; Miller, S.S.; Cui, S.W.; Ma, J.; Zhong, F.; Wang, Q. Protection of heat-sensitive probiotic bacteria during spray-drying by sodium caseinate stabilized fat particles. Food Hydrocoll. 2015, 51, 459–467. [Google Scholar] [CrossRef]
- Choi, J. Lipid Matrix Microencapsulation for Effective Delivery of Essential Oils and Organic Acids to Improve gut Health in Weaned Piglets. Master’s Thesis, University of Manitoba, Winnieg, MB, Canada, 2019. [Google Scholar]
- Tang, Z.; Huang, X.; Baxi, S.; Chambers, J.R.; Sabour, P.M.; Wang, Q. Whey protein improves survival and release characteristics of bacteriophage Felix O1 encapsulated in alginate microspheres. Food Res. Int. 2013, 52, 460–466. [Google Scholar] [CrossRef]
- Jang, I.; Kwon, C.H.; Ha, D.M.; Jung, D.Y.; Kang, S.Y.; Park, M.J.; Han, J.H.; Park, B.-C.; Lee, C.Y. Effects of a lipid-encapsulated zinc oxide supplement on growth performance and intestinal morphology and digestive enzyme activities in weanling pigs. J. Anim. Sci. Technol. 2014, 56, 29. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.J.; Zhu, J.; Wang, B.; Cheng, C.; Du, Y.J.; Wang, M.Q. In vitro stability evaluation of coated lipase. Asian-Australas J. Anim. Sci. 2017, 30, 192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gong, J.; Yu, H.; Guo, Q.; Defelice, C.; Hernandez, M.; Yin, Y.; Wang, Q. Alginate-whey protein dry powder optimized for target delivery of essential oils to the intestine of chickens. Poult. Sci. 2014, 93, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, I.; Ktari, N.; Slima, S.B.; Bouchaala, K.; Salah, R.B. Effects of supplementation with L. plantarum TN8 encapsulated in alginate-chitosan in broiler chickens. Int. J. Biol. Macromol. 2016, 89, 677–681. [Google Scholar] [CrossRef]
- Fernández, H.; Catanese, F.; Puthod, G.; Distel, R.A.; Villalba, J.J. Depression of rumen ammonia and blood urea by quebracho tannin-containing supplements fed after high-nitrogen diets with no evidence of self-regulation of tannin intake by sheep. Small Rumin. Res. 2012, 105, 126–134. [Google Scholar] [CrossRef]
- Adejoro, F.A.; Hassen, A.; Akanmu, A.M. Effect of lipid-encapsulated acacia tannin extract on feed intake, nutrient digestibility and methane emission in sheep. Animals 2019, 9, 863. [Google Scholar] [CrossRef]
- Wang, M.; Huang, H.; Hu, Y.; Huang, J.; Yang, H.; Wang, L.; Chen, S.; Chen, C.; He, S. Effects of dietary microencapsulated tannic acid supplementation on the growth performance, intestinal morphology, and intestinal microbiota in weaning piglets. J. Anim. Sci. 2020, 98, skaa112. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dosages of Tannic Acid | Outcomes | References |
|---|---|---|
| 0.5 g/kg | Improved growth performance and immune system in broiler chickens challenged with Eimeria spp. | [47] |
| 2 g/kg | Improved foot pad dermatitis of the chickens without affecting growth performance | [48] |
| 5 g/kg | Improved growth performance and decreased lipid oxidation. | [49] |
| 5 g/kg | Increased growth performance, modulated cecal microbial metabolites and decreased cecal pH. | [50] |
| 7.5 and 15 g/kg | Decreased growth performance and did not inhibit the growth of Salmonella typhimurium in broiler chickens challenged with S. typhimurium. | [51] |
| 10 g/kg | Attenuated fatty acid profile of breast but decreased growth performance in the heat stressed chickens. | [52] |
| 25 g/kg | Decreased growth performance and showed liver toxicity by inducing liver proteolytic activity | [45] |
| Tannins Sources | Strains | Results and Conclusions | References |
|---|---|---|---|
| Ellagitannins from Chestnut wood Gallotannins from Tara and Sumach (Gall nuts) Condensed tannins from Quebaracho and Calliandra calothyrsus Flavanol gallates from Tea and Acacia nilotica | Salmonella Typhimurium | All of the tannins inhibited the growth of S. Typhimurium. | [63] |
| Tannic acid Gallic acid | S. Typhimurium | Tannic acid and gallic acid had bactericidal effects and gallic acid had higher bactericidal effects than tannic acid | [64] |
| Condensed tannins extracted from tree leaves viz. babool (Acacia nilotica), jamun (Eugenia jambolana), peepal (Ficus religiosa), subabul (Leucaenia leucocephala) and guajava (Psidium guajava) | Escherichia coli Staphylococcus aureus S. enteritis Enterococcus faecalis | All of the selected five condensed tannins inhibited the growth of the four pathogenic bacteria | [65] |
| Tannin extracts from Cytinus hypocistis and C. ruber | S. aureus S. epidermidis E. faecium Pseudomonas aeruginosa Klebsiella pneumoniae | Tannin extracts from C. hypocistis and C. ruber showed antibacterial and antibiofilm activities against gram positive and negative human pathogens. | [66] |
| Chestnut tannins (80% hydrolysable tannins) Quebracho tannins (75% condensed tannins) | Clostridium perfringens | Both hydrolysable and condensed showed antimicrobial effects against C. perfringens and neutralized its cytotoxicity. | [67] |
| Tannins | Other Bioactive Compounds | Outcomes | References |
|---|---|---|---|
| 100 mg/kg tannic acid extract | Probiotics (1 × 104 spores/kg Bacillus coagulans) | Improved feed conversion ratio of coccidiosis vaccinated broilers. | [47] |
| 240 mg/kg tannic acid | Organic acids (420 mg/kg lactic, 480 mg/kg butyric acid and 480 mg/kg acetic acid) | Decreased S. enteritis horizontal transmission in broiler chickens | [161] |
| Chestnut tannins | Saturated short medium chain fatty acids (C4:0 to C12:0) | Showed strong antimicrobial effects in in-vitro conditions and did not affect growth performance and meat quality of in-vivo chicken models. | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Kim, W.K. Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review. Animals 2020, 10, 2389. https://doi.org/10.3390/ani10122389
Choi J, Kim WK. Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review. Animals. 2020; 10(12):2389. https://doi.org/10.3390/ani10122389
Chicago/Turabian StyleChoi, Janghan, and Woo Kyun Kim. 2020. "Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review" Animals 10, no. 12: 2389. https://doi.org/10.3390/ani10122389
APA StyleChoi, J., & Kim, W. K. (2020). Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review. Animals, 10(12), 2389. https://doi.org/10.3390/ani10122389