Heritability of Teat Condition in Italian Holstein Friesian and Its Relationship with Milk Production and Somatic Cell Score

 ,
,  ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dataset
2.2. Statistical Analysis
2.3. Genetic Parameters
2.4. Genetic Change in Teat Score Traits
3. Results
3.1. Data
3.2. Heritability and Environmental Effects Estimates
3.3. Genetic and Environmental Correlations
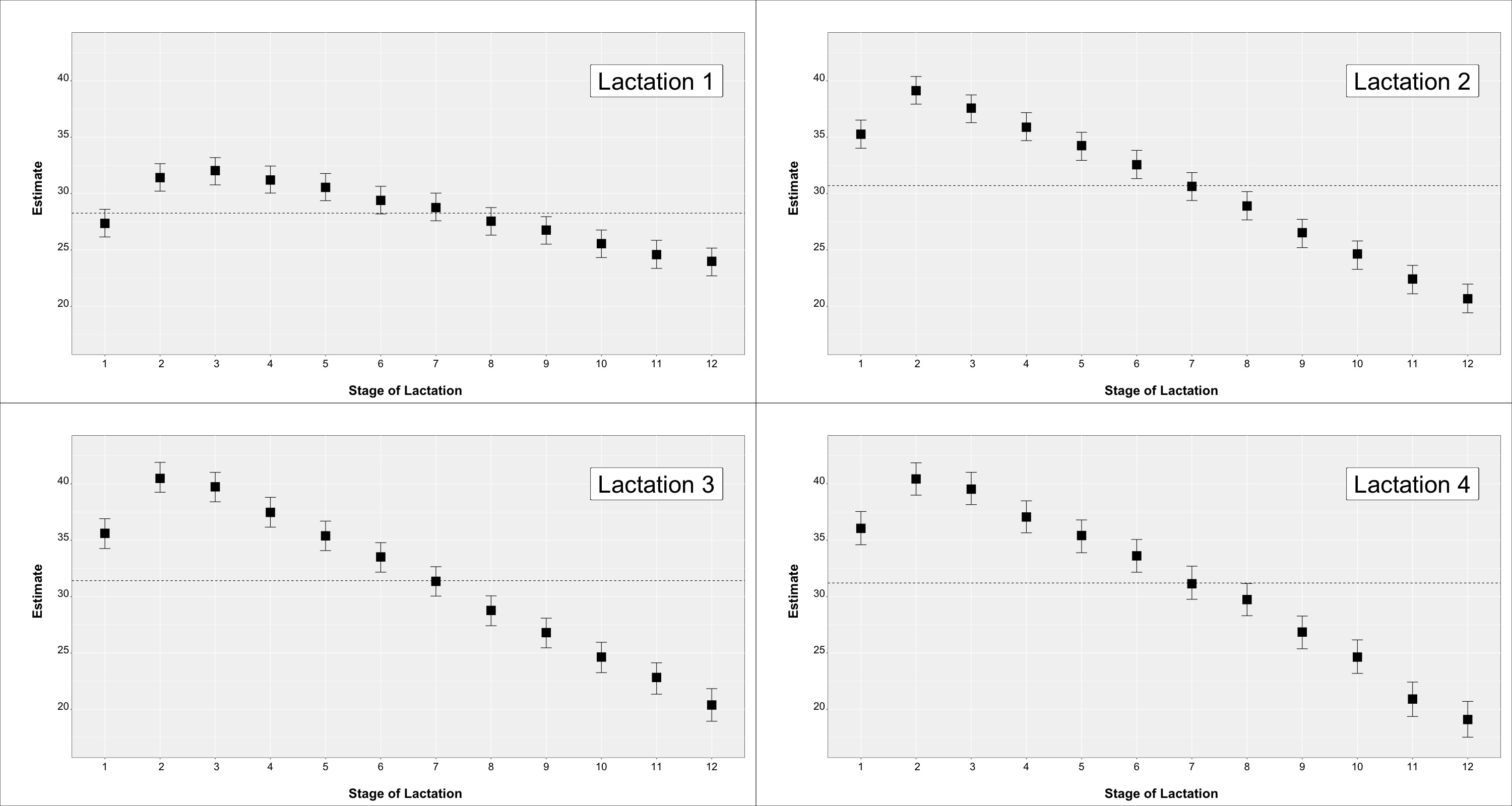
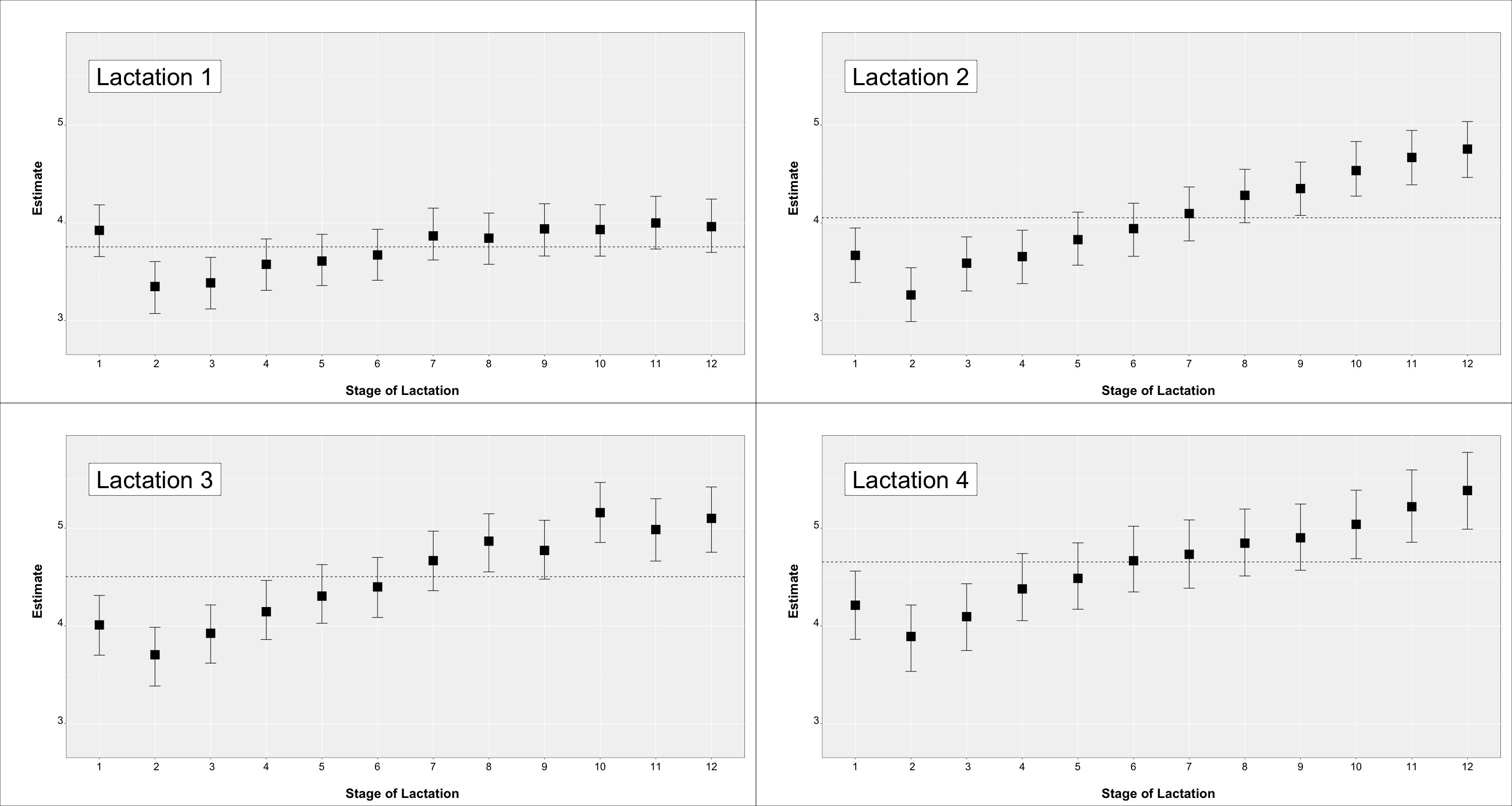
3.4. Influence of Fixed Effects on Teat Scores
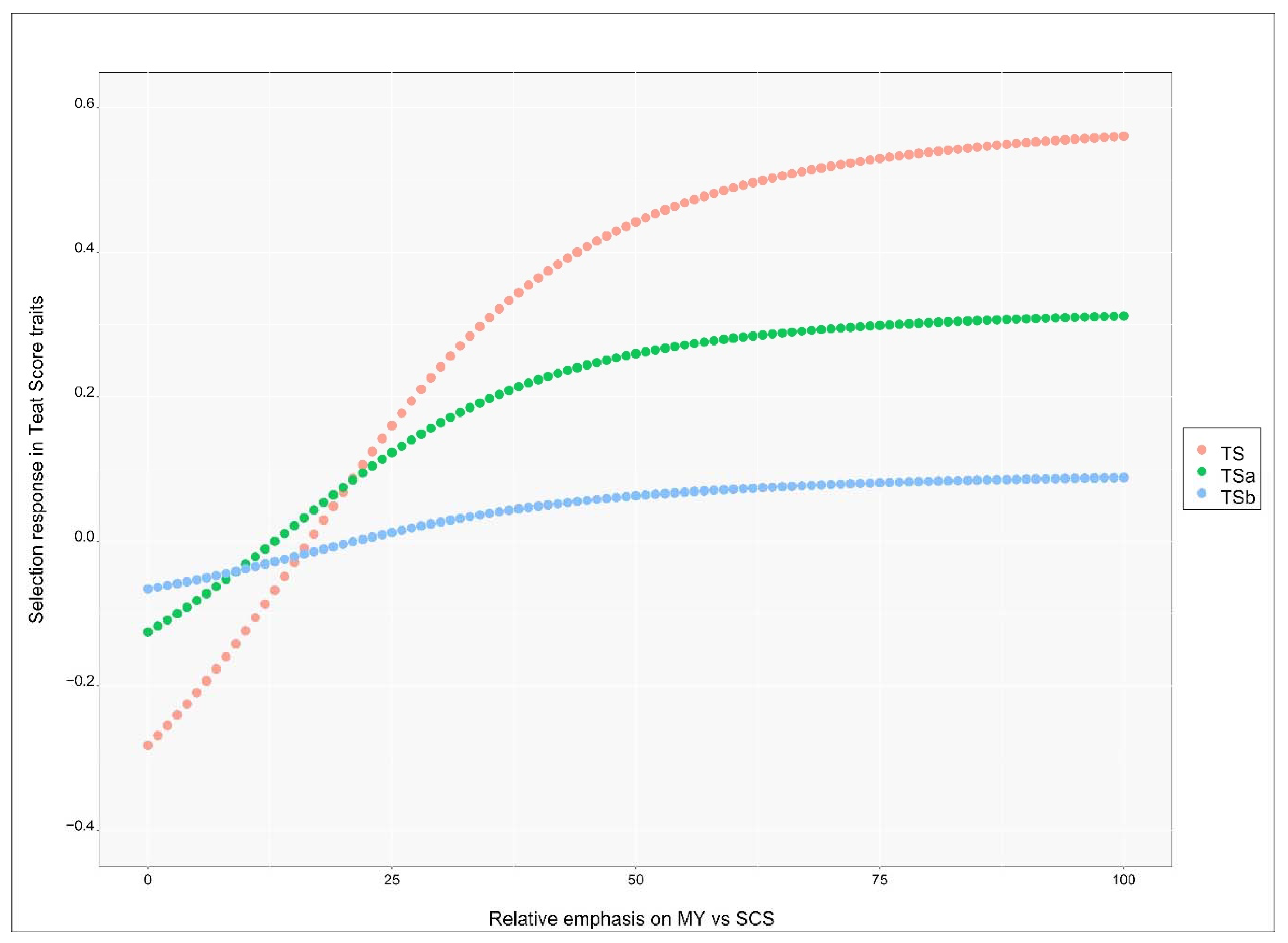
3.5. Genetic Response in Teat Score Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bobbo, T.; Penasa, M.; Finocchiaro, R.; Visentin, G.; Cassandro, M. Alternative somatic cell count traits exploitable in genetic selection for mastitis resistance in Italian Holsteins. J. Dairy Sci. 2018, 101, 10001–10010. [Google Scholar] [CrossRef]
- Santman-Berends, I.M.G.A.; Swinkels, J.M.; Lam, T.J.G.M.; Keurentjes, J.; van Schaik, G. Evaluation of udder health parameters and risk factors for clinical mastitis in Dutch dairy herds in the context of a restricted antimicrobial usage policy. J. Dairy Sci. 2016, 99, 2930–2939. [Google Scholar] [CrossRef]
- Martin, P.; Barkema, H.W.; Brito, L.F.; Narayana, S.G.; Miglior, F. Symposium review: Novel strategies to genetically improve mastitis resistance in dairy cattle. J. Dairy Sci. 2018, 101, 2724–2736. [Google Scholar] [CrossRef]
- Weigel, K.A.; Shook, G.E. Genetic Selection for Mastitis Resistance. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 457–472. [Google Scholar] [CrossRef]
- Zhao, X.; Ponchon, B.; Lanctôt, S.; Lacasse, P. Invited review: Accelerating mammary gland involution after drying-off in dairy cattle. J. Dairy Sci. 2019, 102, 6701–6717. [Google Scholar] [CrossRef]
- Dufour, S.; Fréchette, A.; Barkema, H.W.; Mussell, A.; Scholl, D.T. Invited review: Effect of udder health management practices on herd somatic cell count. J. Dairy Sci. 2011, 94, 563–579. [Google Scholar] [CrossRef]
- Rollin, E.; Dhuyvetter, K.C.; Overton, M.W. The cost of clinical mastitis in the first 30 days of lactation: An economic modeling tool. Prev. Vet. Med. 2015, 122, 257–264. [Google Scholar] [CrossRef]
- Seykora, A.J.; McDaniel, B.T. Udder and teat morphology related to mastitis resistance: A review. J. Dairy Sci. 1985, 68, 2087–2093. [Google Scholar] [CrossRef]
- Neijenhuis, F.; Barkema, H.W.; Hogeveen, H.; Noordhuizen, J.P. Relationship between teat-end callosity and occurrence of clinical mastitis. J. Dairy Sci. 2001, 12, 2664–2672. [Google Scholar] [CrossRef]
- Mein, G.A. The role of the milking machine in mastitis control. Vet. Clin. N. Am. Food Anim. Pract. 2012, 28, 307–320. [Google Scholar] [CrossRef]
- Pantoja, J.C.F.; Correia, L.B.N.; Rossi, R.S.; Latosinski, G.S. Association between teat-end hyperkeratosis and mastitis in dairy cows: A systematic review. J. Dairy Sci. 2020, 103, 1843–1855. [Google Scholar] [CrossRef]
- Magro, G.; Biffani, S.; Minozzi, G.; Ehricht, R.; Monecke, S.; Luini, M.; Piccinini, R. Virulence Genes of S. aureus from Dairy Cow Mastitis and Contagiousness Risk. Toxins 2017, 9, 195. [Google Scholar] [CrossRef]
- Blowey, R.W.; Weaver, A.D. Chapter 11—Udder and teat disorders. In Color Atlas of Diseases and Disorders of Cattle, 3nd ed.; Blowey, R.W., Weaver, A.D., Eds.; Mosby Elsevier: New York, NY, USA, 2011; Volume 3, pp. 203–219. [Google Scholar]
- Odorčić, M.; Rasmussen, M.D.; Paulrud, C.O.; Bruckmaier, R.M. Review: Milking machine settings, teat condition and milking efficiency in dairy cows. Animal 2019, 13, 94–99. [Google Scholar] [CrossRef]
- Neijenhuis, F.; Barkema, H.W.; Hogeveen, H.; Noordhuizen, J.P. Classification and longitudinal examination of callused teat ends in dairy cows. J. Dairy Sci. 2000, 3, 2795–2804. [Google Scholar] [CrossRef]
- Chrystal, M.A.; Seykora, A.J.; Hansen, L.B. Heritabilities of teat end shape and teat diameter and their relationships with somatic cell score. J. Dairy Sci. 1999, 82, 2017–2022. [Google Scholar] [CrossRef]
- Chrystal, M.A.; Seykora, A.J.; Hansen, L.B.; Freeman, A.E.; Kelley, D.H.; Healey, M.H. Heritability of teat-end shape and the relationship of teat-end shape with somatic cell score for an experimental herd of cows. J. Dairy Sci. 2001, 84, 2549–2554. [Google Scholar] [CrossRef]
- Ali, A.K.A.; Shook, G.E. An optimum transformation for somatic cell concentration in milk. J. Dairy Sci. 1980, 63, 487–490. [Google Scholar] [CrossRef]
- Parker Gaddis, K.L.; VanRaden, P.M.; Cole, J.B.; Norman, H.D.; Nicolazzi, E.; Dürr, J.W. Symposium review: Development, implementation, and perspectives of health evaluations in the United States. J. Dairy Sci. 2020, 103, 5354–5365. [Google Scholar] [CrossRef]
- Gleeson, D.; Meaney, W.; O’Callaghan, E. Effect of teat hyperkeratosis on somatic cell counts of dairy cows. Int. J. Appl. Res. Vet. 2004, 2, 115–122. [Google Scholar]
- Bhutto, A.L.; Murray, R.D.; Woldehiwet, Z. Udder shape and teat-end lesions as potential risk factors for high somatic cell counts and intra-mammary infections in dairy cows. Vet. J. 2010, 183, 63–67. [Google Scholar] [CrossRef]
- Haghkhah, M.; Ahmadi, M.R.; Gheisari, H.R.; Kadivar, A. Preliminary bacterial study on subclinical mastitis and teat condition in dairy herds around Shiraz. Turk. J. Vet. Anim. Sci. 2011, 35, 387–394. [Google Scholar] [CrossRef]
- de Pinho Manzi, M.; Nobrega, D.B.; Faccioli, P.Y.; Troncarelli, M.Z.; Menozzi, B.D. Langoni. Relationship between teat-end condition, udder cleanliness and bovine subclinical mastitis. Res. Vet. Sci. 2012, 93, 430–434. [Google Scholar] [CrossRef]
- Mitev, J.E.; Gergovska, I.; Miteva, T.M. Effect of teat end hyperkeratosis on milk somatic cell counts in Bulgarian black-andwhite dairy cattle. Bulg. J. Agric. Sci. 2012, 18, 451–454. [Google Scholar]
- Sandrucci, A.; Bava, L.; Zucali, M.; Tamburini, A. Management factors and cow traits influencing milk somatic cell counts and teat hyperkeratosis during different seasons. Rev. Bras. Zootec. 2014, 43, 505–511. [Google Scholar] [CrossRef]
- Asadpour, R.; Bagherniaee, H.; Houshmandzad, M.; Fatehi, H.; Rafat, A.; Nofouzi, K.; Maftouni, K. Relationship between teat end hyperkeratosis with intra mammary infection and somatic cell counts in lactating dairy cattle. Rev. Med. Vet. 2015, 166, 266–270. [Google Scholar]
- Guarin, J.F.; Paixao, M.G.; Ruegg, P.L. Association of anatomical characteristics of teats with quarter-level somatic cell count. J. Dairy Sci. 2017, 100, 643–652. [Google Scholar] [CrossRef]
- Cremonesi, P.; Pozzi, F.; Raschetti, M.; Bignoli, G.; Capra, E.; Graber, H.U.; Vezzoli, F.; Piccinini, R.; Bertasi, B.; Biffani, S.; et al. Genomic characteristics of Staphylococcus aureus strains associated with high within-herd prevalence of intramammary infections in dairy cows. J Dairy Sci. 2015, 10, 6828–6838. [Google Scholar] [CrossRef]
- Schreiner, D.A.; Ruegg, P.L. Relationship between udder and leg hygiene scores and subclinical mastitis. J. Dairy Sci. 2003, 86, 3460–3465. [Google Scholar] [CrossRef]
- Tsuruta, S.; Misztal, I. THRGIBBS1F90 for estimation of variance components with threshold linear models. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Minas Gerais, Brazil, 13–18 August 2006; pp. 27–31. [Google Scholar]
- Hadfield, J.D. MCMC Methods for Multi-Response Generalized Linear Mixed Models: The MCMCglmm R Package. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef]
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. CODA: Convergence Diagnosis and Output Analysis for MCMC. R News 2006, 6, 7–11. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Cameron, N.D. Selection Indices and Prediction of Genetic Merit in Animal Breeding; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Schneeberger, M.; Barwick, S.A.; Crow, G.H.; Hammond, K. Economic indices using breeding values predicted by BLUP. J. Anim. Breed. Genet. 1992, 109, 180–187. [Google Scholar] [CrossRef]
- Pretto, D.; López-Villalobos, N.; Penasa, M.; Cassandro, M. Genetic response for milk production traits, somatic cell score, acidity and coagulation properties in Italian Holstein–Friesian population under current and alternative selection indices and breeding objectives. Livestock Sci. 2012, 150, 59–66. [Google Scholar] [CrossRef]
- Lojda, L.; Stavikova, M.; Matouskova, O. The shape of the teat and teat-end and the location of the teat canal orifice in relation to subclinical mastitis in cattle. Acta Vet. Brno 1976, 45, 181–185. [Google Scholar]
- Seykora, A.J.; McDaniel, B.T. Heritabilities of Teat Traits and their Relationships with Milk Yield, Somatic Cell Count, and Percent Two-Minute Milk. J. Dairy Sci. 1985, 68, 2670–2683. [Google Scholar] [CrossRef]
- Biffani, S.; Tiezzi, F.; Fresi, P.; Stella, A. Giulietta Minozzi, Genetic parameters of weeping teats in Italian Saanen and Alpine dairy goats and their relationship with milk production and somatic cell score. J. Dairy Sci. 2020, 103, 9167–9176. [Google Scholar] [CrossRef]
- Miglior, F.; Fleming, A.; Malchiodi, F.; Brito, L.F.; Martin, P.; Baes, C.F. A 100-Year Review: Identification and genetic selection of economically important traits in dairy cattle. J. Dairy Sci. 2017, 100, 10251–10271. [Google Scholar] [CrossRef]
- Rupp, R.; Boichard, D. Genetic parameters for clinical mastitis, somatic cell score, production, udder type traits, and milking ease in first lactation Holsteins. J. Dairy Sci. 1999, 82, 2198–2204. [Google Scholar] [CrossRef]
- Tribout, T.; Croiseau, P.; Lefebvre, R.; Barbat, A.; Boussaha, M.; Fritz, S.; Boichard, D.; Hoze, C.; Sanchez, M.P. Confirmed effects of candidate variants for milk production, udder health, and udder morphology in dairy cattle. Genet. Sel. Evol. 2020, 52, 55–67. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Number of Records | Relative Frequency | ||
|---|---|---|---|---|
| TS 1 | TSa 1 | TSb 1 | ||
| 1 | 7078 | 65.7 | 65.7 | 90.6 |
| 2 | 2687 | 24.9 | 34.3 | |
| 3 | 838 | 7.78 | 9.4 | |
| 4 | 173 | 1.61 | ||
| Parameter | Trait | ||||
|---|---|---|---|---|---|
| TS | TSa 1 | TSb 1 | MY 1 | SCS 1 | |
| Sire genetic variance | 0.438 | 0.128 | 0.036 | 0.958 | 0.148 |
| (0.233; 0.702) | (0.072; 0.188) | (0.010; 0.066) | (0.450; 1.608) | (0.084; 0.214) | |
| Cow permanent environmental variance | 3.998 | 1.449 | 0.430 | 15.12 | 1.488 |
| (3.465; 4.532) | (1.324; 1.577) | (0.371; 0.488) | (14.05; 16.15) | (1.398; 1.591) | |
| Herd-test day variance | 1.648 | 0.506 | 0.160 | 15.04 | 0.624 |
| (0.902; 2.548) | (0.288; 0.768) | (0.078; 0.249) | (7.85; 23.06) | (0.292; 0.925) | |
| Residual variance | 1.281 | 1.073 | 1.085 | 28.01 | 1.788 |
| (1.217; 1.343) | (1.051; 1.095) | (1.060; 1.105) | (27.57; 28.51) | (1.758; 1.817) | |
| Heritability (h2) | 0.238 | 0.162 | 0.084 | 0.065 | 0.146 |
| (0.116; 0.364) | (0.090; 0.235) | (0.025; 0.155) | (0.030; 0.109) | (0.080; 0.210) | |
| Intra-Herd heritability (h2-IH) | 0.289 | 0.248 | 0.250 | 0.126 | 0.234 |
| (0.153; 0.45) | (0.061; 0.434) | (0.135; 0.363) | (0.025; 0.217) | (0.114; 0.343) | |
| Cow repeatability | 0.544 | 0.460 | 0.251 | 0.257 | 0.368 |
| (0.469; 0.603) | (0.417; 0.503) | (0.222; 0.282) | (0.220; 0.290) | (0.333; 0.401) | |
| Herd-test-day repeatability | 0.221 | 0.159 | 0.093 | 0.251 | 0.153 |
| (0.141; 0.313) | (0.096; 0.224) | (0.051; 0.141) | (0.164; 0.353) | (0.093; 0.224) | |
| Item | MY 1 | SCS 1 | ||||
|---|---|---|---|---|---|---|
| TS 1 | TSa 1 | TSb 1 | TS 1 | TSa 1 | TSb 1 | |
| Genetic correlation | 0.86 | 0.89 | 0.54 | 0.44 | 0.36 | 0.38 |
| (0.71; 0.98) | (0.61; 0.99) | (−0.11; 0.96) | (0.10; 0.80) | (−0.02; 0.70) | (−0.09; 0.90) | |
| Cow permanent environmental correlation | 0.01 | 0.02 | −0.003 | 0.14 | 0.12 | 0.15 |
| (−0.04; 0.06) | (−0.03; 0.07) | (−0.08; 0.06) | (0.09; 0.19) | (0.07; 0.18) | (0.08; 0.22) | |
| Herd-test day Correlation | 0.30 | 0.30 | 0.46 | −0.25 | −0.25 | −0.47 |
| (−0.06; 0.64) | (−0.03; 0.62) | (0.13; 0.72) | (−0.57; 0.10) | (−0.58; 0.08) | (−0.76; −0.17) | |
| Trait | Hygiene Score 2 | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| TSa 1 | 0.326 | 0.364 | 0.382 | 0.317 |
| (0.285; 0.375) | (0.314; 0.423) | (0.309; 0.451) | (0.248; 0.397) | |
| TSb 1 | 0.093 | 0.094 | 0.094 | 0.095 |
| (0.092; 0.094) | (0.094; 0.094) | (0.094; 0.096) | (0.094; 0.097) | |
| Trait | Udder Quarters | |||
|---|---|---|---|---|
| Front Left | Front Right | Rear Left | Rear Right | |
| TSa 1 | 0.393 | 0.388 | 0.302 | 0.312 |
| (0.327; 0.462) | (0.323; 0.455) | (0.265; 0.34) | (0.271; 0.356) | |
| TSb 1 | 0.095 | 0.095 | 0.094 | 0.094 |
| (0.093; 0.097) | (0.093; 0.097) | (0.092; 0.096) | (0.092; 0.096) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiezzi, F.; Maisano, A.M.; Chessa, S.; Luini, M.; Biffani, S. Heritability of Teat Condition in Italian Holstein Friesian and Its Relationship with Milk Production and Somatic Cell Score. Animals 2020, 10, 2271. https://doi.org/10.3390/ani10122271
Tiezzi F, Maisano AM, Chessa S, Luini M, Biffani S. Heritability of Teat Condition in Italian Holstein Friesian and Its Relationship with Milk Production and Somatic Cell Score. Animals. 2020; 10(12):2271. https://doi.org/10.3390/ani10122271
Chicago/Turabian StyleTiezzi, Francesco, Antonio Marco Maisano, Stefania Chessa, Mario Luini, and Stefano Biffani. 2020. "Heritability of Teat Condition in Italian Holstein Friesian and Its Relationship with Milk Production and Somatic Cell Score" Animals 10, no. 12: 2271. https://doi.org/10.3390/ani10122271
APA StyleTiezzi, F., Maisano, A. M., Chessa, S., Luini, M., & Biffani, S. (2020). Heritability of Teat Condition in Italian Holstein Friesian and Its Relationship with Milk Production and Somatic Cell Score. Animals, 10(12), 2271. https://doi.org/10.3390/ani10122271