Improvement of Disease Resistance in Livestock: Application of Immunogenomics and CRISPR/Cas9 Technology

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
Simple Summary
Abstract
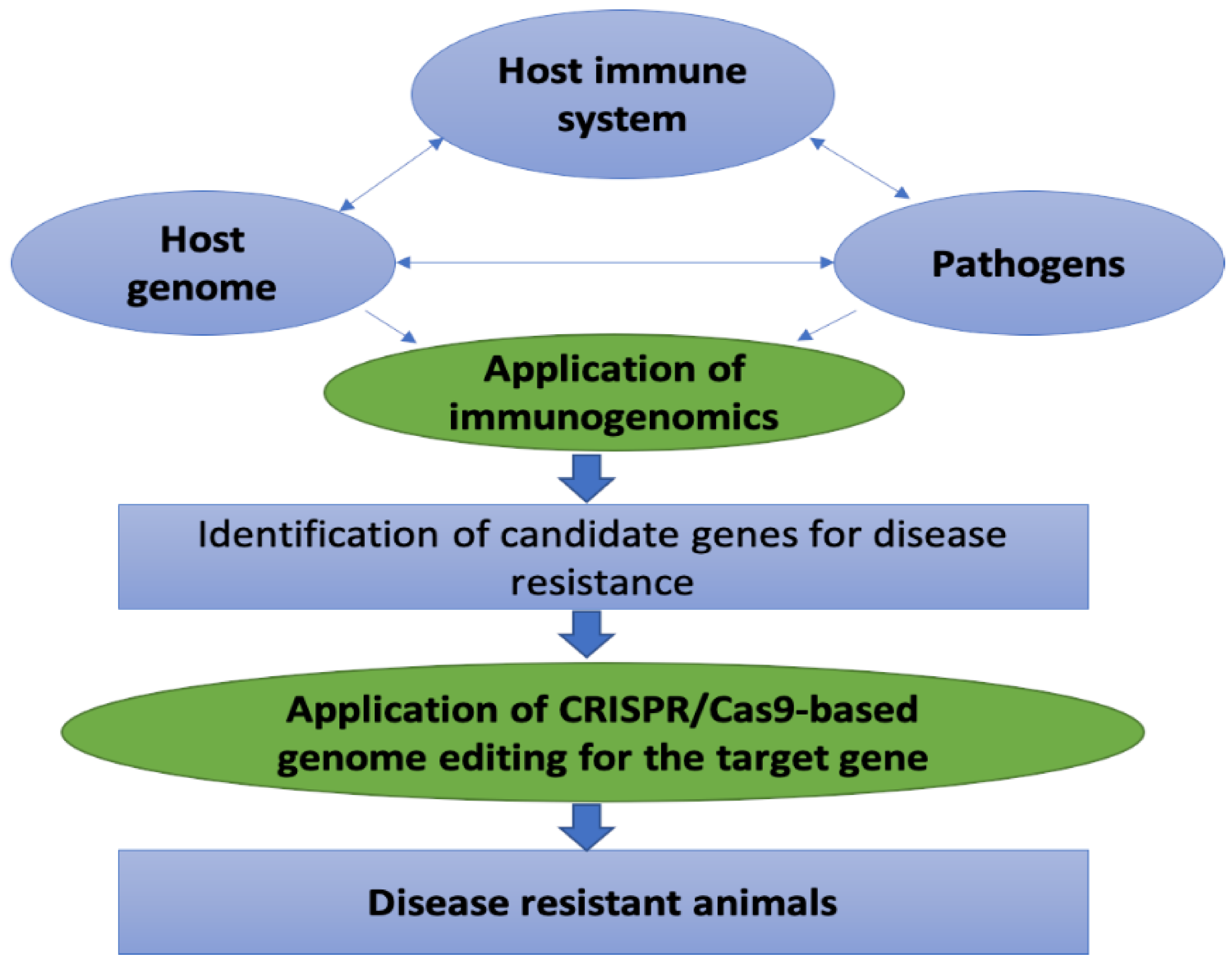
1. Introduction
2. Disease Resistance: The Phenotype
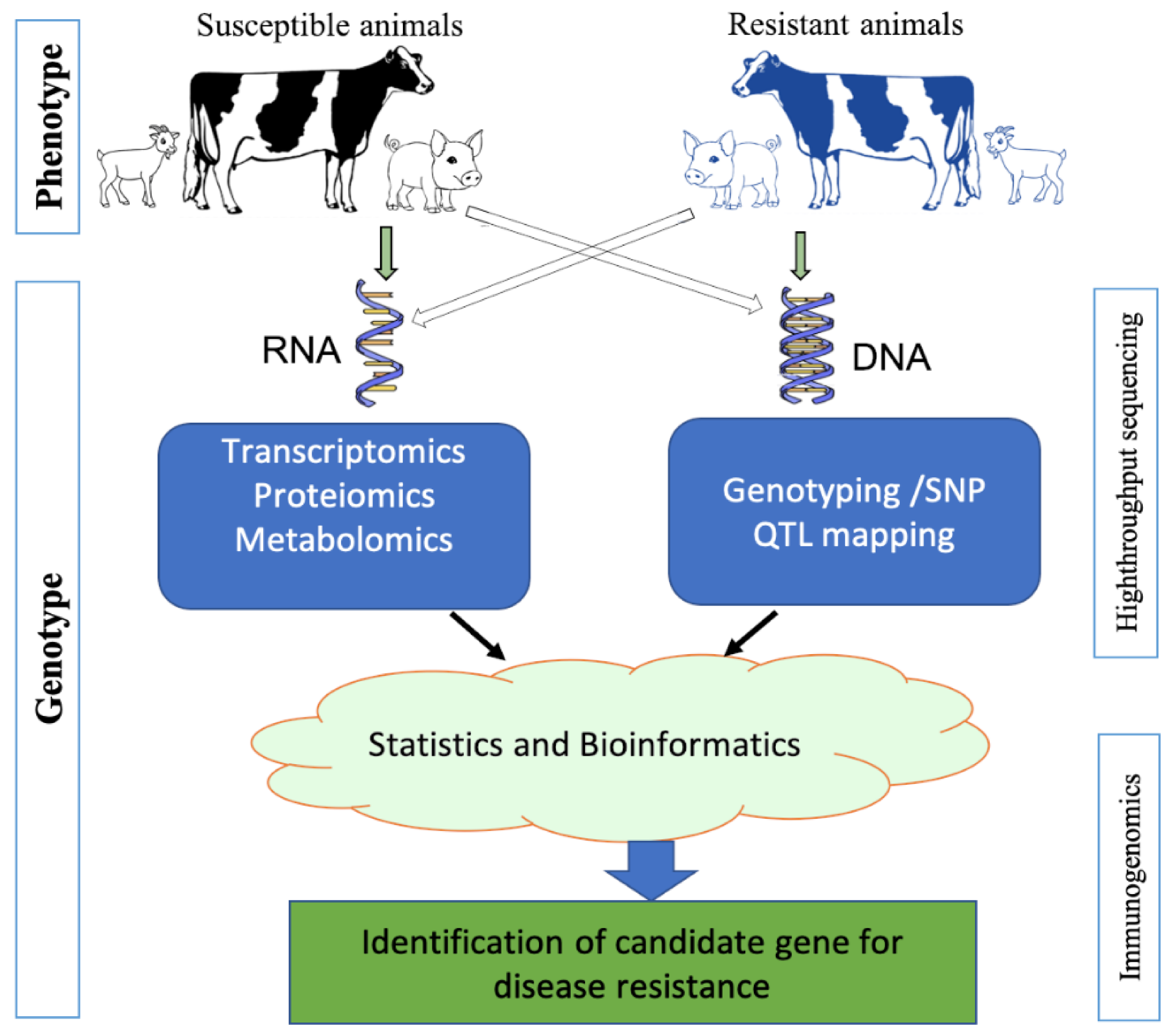
3. Advances of Immunogenomics
3.1. Sequencing Technology
3.2. Bioinformatics Tools
4. Applications of Immunogenomics in Livestock Disease Management
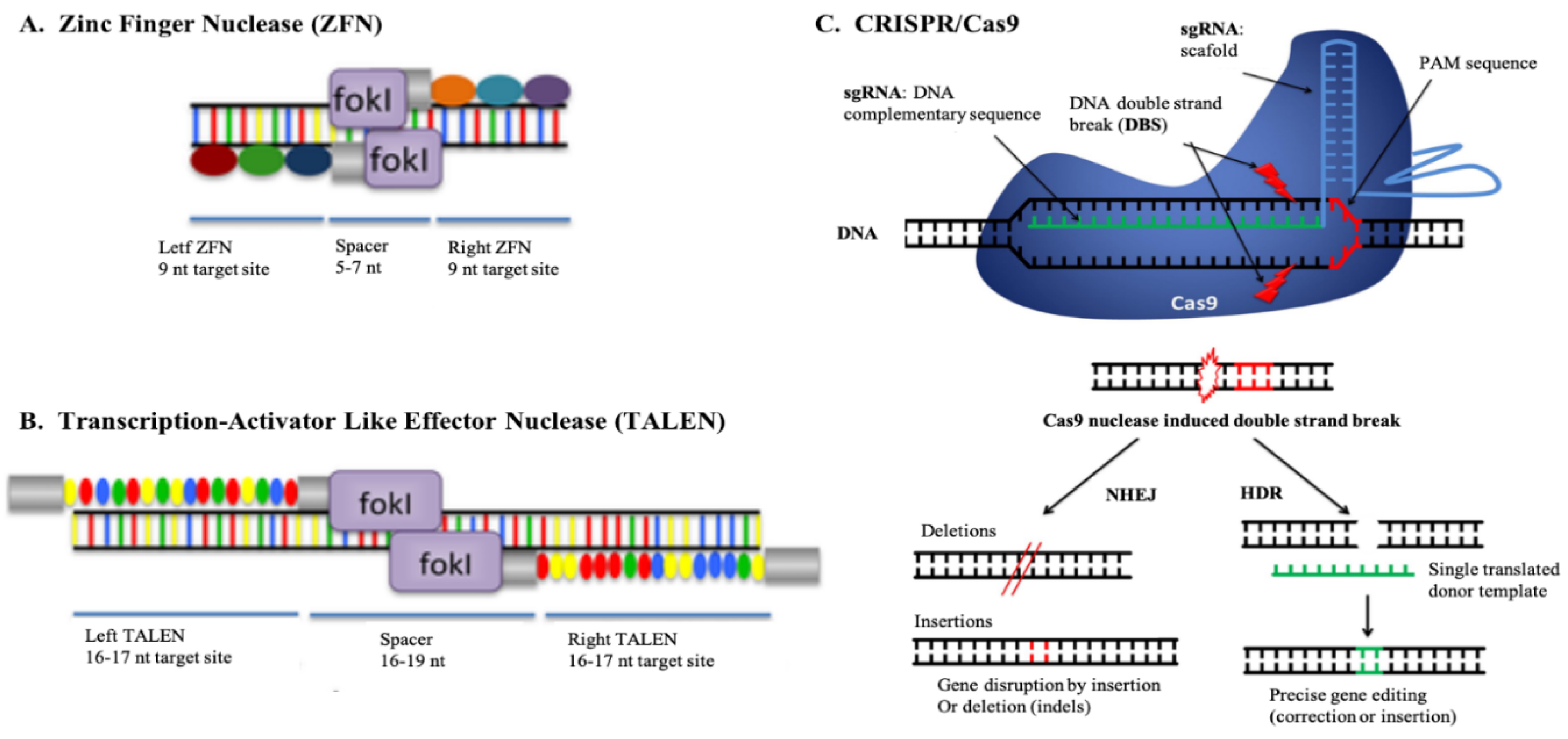
5. Advances of Genome Editing Technology
6. Applications of Genome Editing in Livestock Disease Management
6.1. Porcine Reproductive and Respiratory Syndrome (PRRS) in Pigs
6.2. African Swine Fever Resistance in Pig
6.3. Tuberculosis Resistance in Cattle
6.4. Enzootic Pneumonia Resistance in Cattle
7. Ethics, Regulations, and Social Acceptance of Genome-Edited Livestock Products
8. Potentials and Prospects of CRISPR/Cas9 Technology in Livestock Production
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Aso, H.; Rutten, V.P.M.G.; Takahashi, H.; van Eden, W.; Kitazawa, H. Immunobiotics for the Bovine Host: Their Interaction with Intestinal Epithelial Cells and Their Effect on Antiviral Immunity. Front. Immunol. 2018, 9, 326. [Google Scholar] [CrossRef]
- Davies, G.; Genini, S.; Bishop, S.C.; Giuffra, E. An assessment of opportunities to dissect host genetic variation in resistance to infectious diseases in livestock. Animal 2009, 3, 415–436. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.C.; Woolliams, J.A. Genomics and disease resistance studies in livestock. Livest. Sci. 2014, 166, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Loving, C.L.; Osorio, F.A.; Murtaugh, M.P.; Zuckermann, F.A. Innate and adaptive immunity against Porcine Reproductive and Respiratory Syndrome Virus. Vet. Immunol. Immunopathol. 2015, 167, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Flori, L.; Gao, Y.; Laloe, D.; Lemonnier, G.; Leplat, J.-J.; Teillaud, A.; Cossalter, A.-M.; Laffitte, J.; Pinton, P.; de Vaureix, C.; et al. Immunity traits in pigs: Substantial genetic variation and limited covariation. PLoS ONE 2011, 6, e22717. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, M.; Chen, H. Immunogenomics for identification of disease resistance genes in pigs: A review focusing on Gram-negative bacilli. J. Anim. Sci. Biotechnol. 2012, 3, 34. [Google Scholar] [CrossRef]
- Petersen, B. Basics of genome editing technology and its application in livestock species. Reprod. Domest. Anim. 2017, 52 (Suppl. 3), 4–13. [Google Scholar] [CrossRef]
- Pellagatti, A.; Dolatshad, H.; Valletta, S.; Boultwood, J. Application of CRISPR/Cas9 genome editing to the study and treatment of disease. Arch. Toxicol. 2015, 89, 1023–1034. [Google Scholar] [CrossRef]
- Tait-Burkard, C.; Doeschl-Wilson, A.; McGrew, M.J.; Archibald, A.L.; Sang, H.M.; Houston, R.D.; Whitelaw, C.B.; Watson, M. Livestock 2.0—Genome editing for fitter, healthier, and more productive farmed animals. Genome Biol. 2018, 19, 204. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.C.; Stear, M.J. Modeling of host genetics and resistance to infectious diseases: Understanding and controlling nematode infections. Vet. Parasitol. 2003, 115, 147–166. [Google Scholar] [CrossRef]
- Bishop, S.C. A consideration of resistance and tolerance for ruminant nematode infections. Front. Genet. 2012, 3, 168. [Google Scholar] [CrossRef] [PubMed]
- Grenfell, B.T.; Dobson, A.P. Ecology of Infectious Diseases in Natural Populations; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Albers, G.A.; Gray, G.D.; Piper, L.R.; Barker, J.S.; Le Jambre, L.F.; Barger, I.A. The genetics of resistance and resilience to Haemonchus contortus infection in young merino sheep. Int. J. Parasitol. 1987, 17, 1355–1363. [Google Scholar] [CrossRef]
- Berghof, T.V.L.; Poppe, M.; Mulder, H.A. Opportunities to Improve Resilience in Animal Breeding Programs. Front. Genet. 2019, 9, 692. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Chakravarty, A.K. Genetics and Breeding for Disease Resistance in Livestock; Imprint Academic Press: Cambridge, MA, USA, 2019; p. 384. [Google Scholar] [CrossRef]
- Robbertse, L.; Richards, S.A.; Maritz-Olivier, C. Bovine Immune Factors Underlying Tick Resistance: Integration and Future Directions. Front. Cell. Infect. Microbiol. 2017, 7, 522. [Google Scholar] [CrossRef] [PubMed]
- Bronzo, V.; Lopreiato, V.; Riva, F.; Amadori, M.; Curone, G.; Addis, M.F.; Cremonesi, P.; Moroni, P.; Trevisi, E.; Castiglioni, B. The Role of Innate Immune Response and Microbiome in Resilience of Dairy Cattle to Disease: The Mastitis Model. Animals 2020, 10, 1397. [Google Scholar] [CrossRef] [PubMed]
- Pröll, M.J.; Neuhoff, C.; Schellander, K.; Uddin, M.J.; Cinar, M.U.; Sahadevan, S.; Qu, X.; Islam, M.A.; Müller, N.A.; Drosten, C.; et al. Transcriptome profile of lung dendritic cells after in vitro porcine reproductive and respiratory syndrome virus (PRRSV) infection. PLoS ONE 2017, 12, e0187735. [Google Scholar] [CrossRef]
- Islam, M.A.; Neuhoff, C.; Rony, S.A.; Grosse-Brinkhaus, C.; Uddin, M.J.; Hoelker, M.; Tesfaye, D.; Tholen, E.; Schellander, K.; Proll-Cornilissen, M.J. PBMCs transcriptome profiles identified breed-specific transcriptome signatures for PRRSV vaccination in German Landrace and Pietrain pigs. PLoS ONE 2019, 14, e0222513. [Google Scholar] [CrossRef]
- Chris, T. Transcriptomics today: Microarrays, RNA-seq, and more. Science 2015, 349, 544–546. [Google Scholar]
- Spitzer, M.H.; Nolan, G.P. Mass Cytometry: Single Cells, Many Features. Cell 2016, 165, 780–791. [Google Scholar] [CrossRef]
- Nishimura, T.; Kaneko, S.; Kawana-Tachikawa, A.; Tajima, Y.; Goto, H.; Zhu, D.; Nakayama-Hosoya, K.; Iriguchi, S.; Uemura, Y.; Shimizu, T.; et al. Generation of rejuvenated antigen-specific T cells by reprogramming to pluripotency and redifferentiation. Cell Stem Cell 2013, 12, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. DNA sequencing technologies: 2006–2016. Nat. Protoc. 2017, 12, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Daugharthy, E.R.; Scheiman, J.; Kalhor, R.; Yang, J.L.; Ferrante, T.C.; Terry, R.; Jeanty, S.S.F.; Li, C.; Amamoto, R.; et al. Highly multiplexed subcellular RNA sequencing in situ. Science 2014, 343, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef] [PubMed]
- Coskun, A.F.; Eser, U.; Islam, S. Cellular identity at the single-cell level. Mol. BioSyst. 2016, 12, 2965–2979. [Google Scholar] [CrossRef]
- Huang, X.; Li, X.; Qin, P.; Zhu, Y.; Xu, S.; Chen, J. Technical Advances in Single-Cell RNA Sequencing and Applications in Normal and Malignant Hematopoiesis. Front. Oncol. 2018, 8, 582. [Google Scholar] [CrossRef]
- Garalde, D.R.; Snell, E.A.; Jachimowicz, D.; Sipos, B.; Lloyd, J.H.; Bruce, M.; Pantic, N.; Admassu, T.; James, P.; Warland, A.; et al. Highly parallel direct RNA sequencing on an array of nanopores. bioRXiv 2016. [Google Scholar] [CrossRef]
- Oikonomopoulos, S.; Bayega, A.; Fahiminiya, S.; Djambazian, H.; Berube, P.; Ragoussis, J. Methodologies for Transcript Profiling Using Long-Read Technologies. Front. Genet. 2020, 11, 606. [Google Scholar] [CrossRef]
- Goodwin, S.; Mcpherson, J.D.; Mccombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The reference human genome annotation for the ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef]
- Matlin, A.J.; Clark, F.; Smith, C.W. Understanding alternative splicing: Towards a cellular code. Nat. Rev. Mol. Cell Biol. 2005, 6, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, P.G.; Steijger, T.; Sipos, B.; Grant, G.R.; Kahles, A.; Ratsch, G.; Goldman, N.; Hubbard, T.J.; Harrow, J.; Guigó, R.; et al. Systematic evaluation of spliced alignment programs for RNA-seq data. Nat. Methods 2013, 10, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Steijger, T.; Abril, J.F.; Engstrom, P.G.; Kokocinski, F.; Hubbard, T.J.; Guigo, R.; Harrow, J.; Bertone, P.; The RGASP Consortium. Assessment of transcript reconstruction methods for RNA-seq. Nat. Methods 2013, 10, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Cartolano, M.; Huettel, B.; Hartwig, B.; Reinhardt, R.; Schneeberger, K. cDNA library enrichment of full-length transcripts for smrt long read sequencing. PLoS ONE 2016, 11, e0157779. [Google Scholar] [CrossRef]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 5, 115–121. [Google Scholar] [CrossRef]
- Krøigard, A.B.; Thomassen, M.; Lænkholm, A.-V.; Kruse TALarsen, M.J. Evaluation of Nine Somatic Variant Callers for Detection of Somatic Mutations in Exome and Targeted Deep Sequencing Data. PLoS ONE 2016, 11, e0151664. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Carissa, A.P.; James, M.R. Building a livestock genetic and genomic information knowledgebase through integrative developments of Animal QTLdb and CorrDB. Nucleic Acids Res. 2019, 47, D701–D710. [Google Scholar] [CrossRef]
- Greiff, V.; Bhat, P.; Cook, S.C.; Menzel, U.; Kang, W.; Reddy, S.T. A bioinformatic framework for immune repertoire diversity profiling enables detection of immunological status. Genome Med. 2015, 7, 49. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Breuer, K.; Amir, K.F.; Matthew, R.L.; Carol, C.; Anastasia, S.; Raymond, L.; Geoffrey, L.; Robert, E.W.H.; Fiona, S.L.B.; David, J.L. InnateDB: Systems biology of innate immunity and beyond—Recent updates and continuing curation. Nucleic Acids Res. 2013, 41, D1228–D1233. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef] [PubMed]
- Elsik, C.G.; Unni, D.R.; Diesh, C.M.; Tayal, A.; Emery, M.L.; Nguyen, H.N.; Hagen, D.E. Bovine Genome Database: New tools for gleaning function from the Bos taurus genome. Nucleic Acids Res. 2016, 44, D834–D839. [Google Scholar] [CrossRef] [PubMed]
- Mudunuri, U.; Che, A.; Yi, M.; Stephens, R.M. bioDBnet: The biological database network. Bioinformatics 2009, 25, 555–556. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Holt, R.A. Immunogenomics: A foundation for intelligent immune design. Genome Med. 2015, 7, 116. [Google Scholar] [CrossRef][Green Version]
- Tuggle, C.K.; Giuffra, E.; White, S.N.; Clarke, L.; Zhou, H.; Ross, P.J.; Acloque, H.; Reecy, J.M.; Archibald, A.; Bellone, R.R.; et al. GO-FAANG meeting: A Gathering on Functional Annotation of Animal Genomes. Anim. Genet. 2016, 47, 528–533. [Google Scholar] [CrossRef]
- Doeschl-Wilson, A.B.; Bishop, S.C.; Kyriazakis, I.; Villanueva, B. Novel methods for quantifying individual host response to infectious pathogens for genetic analyses. Front. Genet. 2012, 3, 266. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, B.; Mallard, B. Selection for high immune response: An alternative approach to animal health maintenance? Vet. Immunol. Immunopathol. 1999, 72, 231–235. [Google Scholar] [CrossRef]
- Rowland, R.R.; Lunney, J.; Dekkers, J. Control of porcine reproductive and respiratory syndrome (PRRS) through genetic improvements in disease resistance and tolerance. Front. Genet. 2012, 3, 260. [Google Scholar] [CrossRef]
- Clapperton, M.; Diack, A.B.; Matika, O.; Glass, E.J.; Gladney, C.D.; Mellencamp, M.A.; Hoste, A.; Bishop, S.C. Traits associated with innate and adaptive immunity in pigs: Heritability and associations with performance under different health status conditions. Genet. Sel. Evol. 2009, 41, 54. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, C.-A. Vaccine: General Aspects of Vaccinations; Elsevier Health Sciences Saunders: Amsterdam, The Netherlands, 2012; pp. 17–36. [Google Scholar]
- Glass, E.J. The molecular pathways underlying host resistance and tolerance to pathogens. Front. Genet. 2012, 3, 263. [Google Scholar] [CrossRef]
- Edfors-Lilja, I.; Wattrang, E.; Magnusson, U.; Fossum, C. Genetic variation in parameters reflecting immune competence of swine. Vet. Immunol. Immunopathol. 1994, 40, 1–16. [Google Scholar] [CrossRef]
- Schurch, N.J.; Schofield, P.; Gierliński, M.; Cole, C.; Sherstnev, A.; Singh, V.; Wrobel, N.; Gharbi, K.; Simpson, G.G.; Owen-Hughes, T.; et al. How many biological replicates are needed in an RNA-seq experiment and which differential expression tool should you use? RNA 2016, 22, 839–851, [published correction appears in RNA. 2016, 22(10):1641]. [Google Scholar] [CrossRef]
- Liu, X.S.; Mardis, E.R. Applications of Immunogenomics to Cancer. Cell 2017, 168, 600–612. [Google Scholar] [CrossRef]
- Sweeney, T.; Hanrahan, J.P.; Ryan, M.T.; Good, B. Immunogenomics of gastrointestinal nematode infection in ruminants—Breeding for resistance to produce food sustainably and safely. Parasite Immunol. 2016, 38, 569–586. [Google Scholar] [CrossRef]
- Sodeland, M.; Kent, M.P.; Olsen, H.G.; Opsal, M.A.; Svendsen, M.; Sehested, E.; Hayes, B.J.; Lien, S. Quantitative trait loci for clinical mastitis on chromosomes 2, 6, 14 and 20 in Norwegian Red cattle. Anim. Genet. 2011, 42, 457–465. [Google Scholar] [CrossRef]
- Meredith, B.K.; Berry, D.P.; Kearney, F.; Finlay, E.K.; Fahey, A.G.; Bradley, D.G.; Lynn, D.J. A genome-wide association study for somatic cell score using the Illumina high-density bovine beadchip identifies several novel QTL potentially related to mastitis susceptibility. Front. Genet. 2013, 4, 229. [Google Scholar] [CrossRef] [PubMed]
- Wijga, S.; Bastiaansen, J.W.; Wall, E.; Strandberg, E.; de Haas, Y.; Giblin, L.; Bovenhuis, H. Genomic associations with somatic cell score in first-lactation Holstein cows. J. Dairy Sci. 2012, 95, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska-Sabat, A.M.; Gunther, J.; Seyfert, H.M.; Olsaker, I. Combining quantitative trait loci and heterogeneous microarray data analyses reveals putative candidate pathways affecting mastitis in cattle. Anim. Genet. 2012, 43, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Takagi, M.; Fukuyama, K.; Komatsu, R.; Albarracin, L.; Nochi, T.; Suda, Y.; Ikeda-Ohtsubo, W.; Rutten, V.; Eden, V.W.; et al. Transcriptome analysis of inflammatory responses of bovine mammary epithelial cells: Exploring immunomodulatory target genes for bovine mastitis. Pathogens 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Brinster, R.L.; Hammer, R.E.; Trumbauer, M.E.; Rosenfeld, M.G.; Birnberg, N.C.; Evans, R.M. Dramatic growth of mice that develop from eggs microinjected with metallothionein-growth hormone fusion genes. Nature 1982, 300, 611–615. [Google Scholar] [CrossRef]
- Ivics, Z.; Hackett, P.B.; Plasterk, R.H.; Izsvák, Z. Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 1997, 91, 501–510. [Google Scholar] [CrossRef]
- Chevalier, B.S.; Stoddard, B.L. Homing endonucleases: Structural and functional insight into the catalysts of intron/intein mobility. Nucleic Acids Res. 2001, 29, 3757–3774. [Google Scholar] [CrossRef]
- Moore, F.E.; Reyon, D.; Sander, J.D.; Martinez, S.A.; Blackburn, J.S.; Khayter, C.; Ramirez, C.L.; Joung, J.K.; Langenau, D.M. Improved Somatic Mutagenesis in Zebrafish Using Transcription Activator-Like Effector Nucleases (TALENs). PLoS ONE 2012, 7, e37877. [Google Scholar] [CrossRef]
- Belfort, M.; Bonocora, R.P. Homing endonucleases: From genetic anomalies to programmable genomic clippers. Methods Mol. Biol. 2014, 1123, 1–26. [Google Scholar] [CrossRef]
- Joung, J.K.; Sander, J.D. TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Christian, M.; Cermak, T.; Doyle, E.L.; Schmidt, C.; Zhang, F.; Hummel, A.; Bogdanove, A.J.; Voytas, D.F. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 2010, 186, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Shao, Y.; Guan, Y.; Wang, L.; Qiu, Z.; Liu, M.; Chen, Y.; Wu, L.; Li, Y.; Ma, X.; Liu, M.; et al. CRISPR/Cas-mediated genome editing in the rat via direct injection of one-cell embryos. Nat. Protoc. 2014, 9, 2493–2512. [Google Scholar] [CrossRef]
- Harwood, W.; Proudfoot, C.; Burkard, C. Genome editing for disease resistance in livestock. Emerg. Top. Life Sci. 2017, 1, 209–219. [Google Scholar] [CrossRef]
- Lee, H.; Yoon, D.E.; Kim, K. Genome editing methods in animal models. Anim. Cells Syst. 2020, 24, 8–16. [Google Scholar] [CrossRef]
- Zhao, J.; Lai, L.; Ji, W.; Zhou, Q. Genome editing in large animals: Current status and future prospects. Nat. Sci. Rev. 2019, 6, 402–420. [Google Scholar] [CrossRef]
- Shrock, E.; Güell, M. CRISPR in Animals and Animal Models; Academic Press: Cambridge, MA, USA, 2017; pp. 95–114. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious Diseases of Humans: Dynamics and Control; Oxford University Press: New York, NY, USA, 1991; Available online: https://global.oup.com/academic/product/infectious-diseases-of-humans-9780198540403?cc=gb&lang=en& (accessed on 26 July 2020).
- Wall, R.J.; Powell, A.M.; Paape, M.J.; Kerr, D.E.; Bannerman, D.D.; Pursel, V.G.; Wells, K.D.; Talbot, N.; Hawk, H.W. Genetically enhanced cows resist intramammary Staphylococcus aureus infection. Nat. Biotechnol. 2005, 23, 445–451. [Google Scholar] [CrossRef]
- Yu, S.; Luo, J.; Song, Z.; Ding, F.; Dai, Y.; Li, N. Highly efficient modification of beta-lactoglobulin (BLG) gene via zinc-finger nucleases in cattle. Cell Res. 2011, 21, 1638–1640. [Google Scholar] [CrossRef]
- Carlson, D.F.; Tan, W.; Lillico, S.G.; Stverakova, D.; Proudfoot, C.; Christian, M.; Fahrenkrug, S.C. Efficient TALEN-mediated gene knockout in livestock. Proc. Natl. Acad. Sci. USA 2012, 109, 17382–17387. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.F.; Lancto, C.A.; Zang, B.; Kim, E.S.; Walton, M.; Oldeschulte, D.; Fahrenkrug, S.C. Production of hornless dairy cattle from genome-edited cell lines. Nat. Biotechnol. 2016, 34, 479–481. [Google Scholar] [CrossRef] [PubMed]
- Maga, E.A.; Cullor, J.S.; Smith, W.; Anderson, G.B.; Murray, J.D. Human lysozyme expressed in the mammary gland of transgenic dairy goats can inhibit the growth of bacteria that cause mastitis and the cold-spoilage of milk. Foodborne Pathog. Dis. 2006, 3, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wu, H.; Wang, Y.; Liu, X.; Chen, L.; Li, Q.; Cui, C.; Liu, X.; Zhang, J.; Zhang, Y. Single Cas9 nickase induced generation of NRAMP1 knockin cattle with reduced off-target effects. Genome Biol. 2017, 18, 13. [Google Scholar] [CrossRef] [PubMed]
- Tuggle, C.K.; Waters, W.R. Tuberculosis-resistant transgenic cattle. Proc. Natl. Acad. Sci. USA 2015, 112, 3854–3855. [Google Scholar] [CrossRef]
- Lu, T.; Song, Z.; Li, Q.; Li, Z.; Wang, M.; Liu, L.; Tian, K.; Li, N. Overexpression of histone deacetylase 6 enhances resistance to porcine reproductive and respiratory syndrome virus in pigs. PLoS ONE 2017, 12, e0169317. [Google Scholar] [CrossRef]
- Islam, M.A.; Große-Brinkhaus, C.; Pröll, M.J.; Uddin, M.J.; Rony, S.A.; Tesfaye, D.; Tholen, E.; Hölker, M.; Schellander, K.; Neuhoff, C. Deciphering transcriptome profiles of peripheral blood mononuclear cells in response to PRRSV vaccination in pigs. BMC Genom. 2016, 17, 641. [Google Scholar] [CrossRef]
- Islam, M.A.; Große-Brinkhaus, C.; Pröll, M.J.; Uddin, M.J.; Rony, S.A.; Tesfaye, D.; Tholen, E.; Hoelker, M.; Schellander, K.; Neuhoff, C. PBMC transcriptome profiles identifies potential candidate genes and functional networks controlling the innate and the adaptive immune response to PRRSV vaccine in Pietrain pig. PLoS ONE 2017, 12, e0171828. [Google Scholar] [CrossRef]
- Van Gorp, H.; Van Breedam, W.; Delputte, P.L.; Nauwynck, H.J. The porcine reproductive and respiratory syndrome virus requires trafficking through CD163-positive early endosomes, but not late endosomes, for productive infection. Arch. Virol. 2009, 154, 1939–1943. [Google Scholar] [CrossRef]
- Duan, X.; Nauwynck, H.J.; Favoreel, H.W.; Pensaert, M.B. Identification of a putative receptor for porcine reproductive and respiratory syndrome virus on porcine alveolar macrophages. J. Virol. 1998, 72, 4520–4523. [Google Scholar] [CrossRef]
- Whitworth, K.M.; Rowland, R.R.; Ewen, C.L.; Trible, B.R.; Kerrigan, M.A.; Cino-Ozuna, A.G.; Samuel, M.S.; Lightner, J.E.; McLaren, D.G.; Mileham, A.J.; et al. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus. Nat. Biotechnol. 2016, 34, 20–22. [Google Scholar] [CrossRef]
- Burkard, C.; Lillico, S.G.; Reid, E.; Jackson, B.; Mileham, A.J.; Ait-Ali, T.; Whitelaw, C.B.; Archibald, A.L. Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function. PLoS Pathog. 2017, 13, e1006206. [Google Scholar] [CrossRef] [PubMed]
- Palgrave, C.J.; Gilmour, L.; Lowden, C.S.; Lillico, S.G.; Mellencamp, M.A.; Whitelaw, C.B.A. Species-specific variation in RELA underlies differences in NF-κB activity: A potential role in african swine fever pathogenesis. J. Virol. 2011, 85, 6008–6014. [Google Scholar] [CrossRef] [PubMed]
- Lillico, S.G.; Proudfoot, C.; King, T.J.; Tan, W.; Zhang, L.; Mardjuki, R.; Paschon, D.E.; Rebar, E.J.; Urnov, F.D.; Mileham, A.J.; et al. Mammalian interspecies substitution of immune modulatory alleles by genome editing. Sci. Rep. 2016, 6, 21645. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.L.; Müller, B.; van Helden, P.D. Mycobacterium bovis at the animal–human interface: A problem, or not? Vet. Microbiol. 2010, 140, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, Y.; Zhang, Y.; Yang, M.; Lv, J.; Liu, J.; Zhang, Y. TALE nickase-mediated SP110 knockin endows cattle with increased resistance to tuberculosis. Proc. Natl. Acad. Sci. USA 2015, 112, E1530–E1539. [Google Scholar] [CrossRef] [PubMed]
- Shanthalingam, S.; Tibary, A.; Beever, J.E.; Kasinathan, P.; Brown, W.C.; Srikumaran, S. Precise gene editing paves the way for derivation of Mannheimia haemolytica leukotoxin-resistant cattle. Proc. Natl. Acad. Sci. USA 2016, 113, 13186–13190. [Google Scholar] [CrossRef]
- Ishii, T. Genome-edited livestock: Ethics and social acceptance. Anim. Front. 2017, 7. [Google Scholar] [CrossRef]
- Bruce, A. Genome edited animals: Learning from GO crops? Transgenic Res. 2017, 26, 385–398. [Google Scholar] [CrossRef]
- Araki, M.; Ishii, T. Towards social acceptance of plant breeding by genome editing. Trends Plant Sci. 2015, 20, 145–149. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Bioinformatics Tools/Databases | Potential Implications | References |
|---|---|---|
| ‘Bowtie’, ‘msa’ | Sequence-read alignment, | https://cran.r-project.org/ |
| R/Bioconductor, limma, DESeq2 | Differential gene expression analyses | https://cran.r-project.org/ |
| GSEA-Gene Set Enrichment Analysis | Gene set enrichment analysis | [41] https://www.gsea-msigdb.org/gsea/index.jsp |
| DAVID | Gene ontology and pathway analysis | [42] https://david.ncifcrf.gov/ |
| KEEG-Kyoto Encyclopedia of Genes and Genomes | Gene ontology and pathway analysis | https://www.genome.jp/kegg/ |
| InnateDB | Database for gene ontology, pathway analysis and prediction interactome | [43] https://www.innatedb.com/ |
| REACTOME | Database for gene ontology and pathway analysis | [44] https://reactome.org/ |
| QTLdb | Database of quantitative trait loci of animals | [39] https://www.animalgenome.org/cgi-bin/QTLdb/index |
| BovineMine | Annotation and functions of gene | [45] http://128.206.116.13:8080/bovinemine/begin.do |
| bioDBnet-Biological database network | Interconnected access to many types of biological databases, conversion of gene or protein identifies | [46] https://biodbnet-abcc.ncifcrf.gov/ |
| STRING | Functional protein association network analysis and visualization | [47] https://string-db.org/ |
| NetworkAnalyst | Co-regulatory gene or protein network analysis and visualization | [48] https://www.networkanalyst.ca/ |
| WGCNA | Weighted gene co-expression network analysis | [49] https://cran.r-project.org/ |
| Cytoscape | Creation and visualization gene network | [50] https://cytoscape.org/ |
| Species | Disease | Targets of Genome Modification | Reference |
|---|---|---|---|
| Goat | Mastitis | Lysozyme (human) | [88] |
| Cattle | Mastitis | Lysostaphin (Staphylococcus simulans) | [84] |
| Enzootic pneumonia | Cluster of differentiation 18 (CD18) | [89] | |
| Tuberculosis | The natural resistance to infection with intracellular pathogens 1 (NRAMP1) gene | [90] | |
| Pigs | African swine fever | RELA | [86] |
| PRRS | Histone deacetylase HDAC6 | [91] | |
| Cluster of differentiation 163 (CD163) | [92,93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.; Rony, S.A.; Rahman, M.B.; Cinar, M.U.; Villena, J.; Uddin, M.J.; Kitazawa, H. Improvement of Disease Resistance in Livestock: Application of Immunogenomics and CRISPR/Cas9 Technology. Animals 2020, 10, 2236. https://doi.org/10.3390/ani10122236
Islam MA, Rony SA, Rahman MB, Cinar MU, Villena J, Uddin MJ, Kitazawa H. Improvement of Disease Resistance in Livestock: Application of Immunogenomics and CRISPR/Cas9 Technology. Animals. 2020; 10(12):2236. https://doi.org/10.3390/ani10122236
Chicago/Turabian StyleIslam, Md. Aminul, Sharmin Aqter Rony, Mohammad Bozlur Rahman, Mehmet Ulas Cinar, Julio Villena, Muhammad Jasim Uddin, and Haruki Kitazawa. 2020. "Improvement of Disease Resistance in Livestock: Application of Immunogenomics and CRISPR/Cas9 Technology" Animals 10, no. 12: 2236. https://doi.org/10.3390/ani10122236
APA StyleIslam, M. A., Rony, S. A., Rahman, M. B., Cinar, M. U., Villena, J., Uddin, M. J., & Kitazawa, H. (2020). Improvement of Disease Resistance in Livestock: Application of Immunogenomics and CRISPR/Cas9 Technology. Animals, 10(12), 2236. https://doi.org/10.3390/ani10122236