Assessing the Diversity and Population Substructure of Sarda Breed Bucks by Using Mtdna and Y-Chromosome Markers

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Sampling and DNA Purification
2.3. Y-Chromosome Genotyping
2.4. Mitochondrial DNA Analysis
2.5. Statistical and Computational Analysis
3. Results
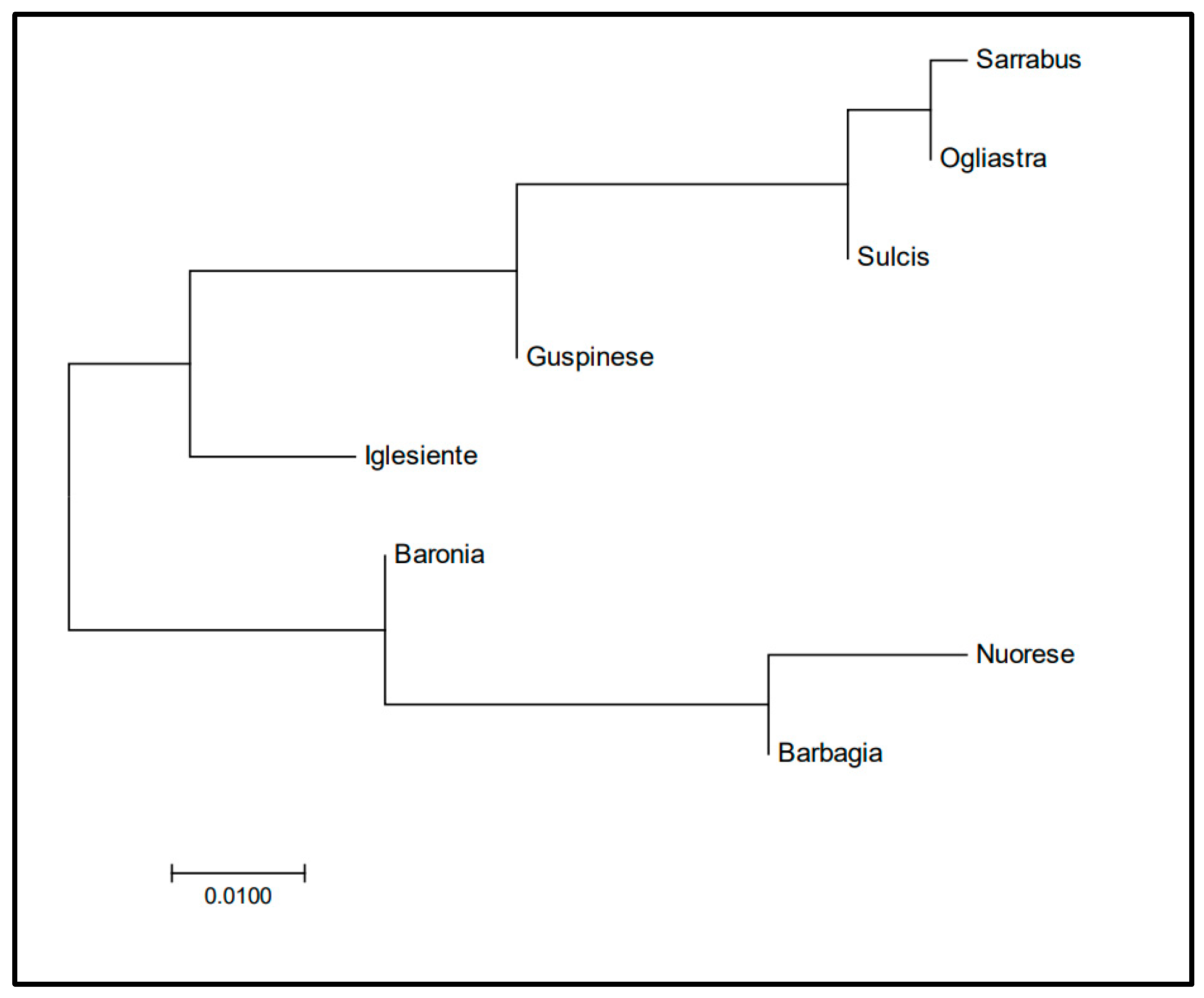
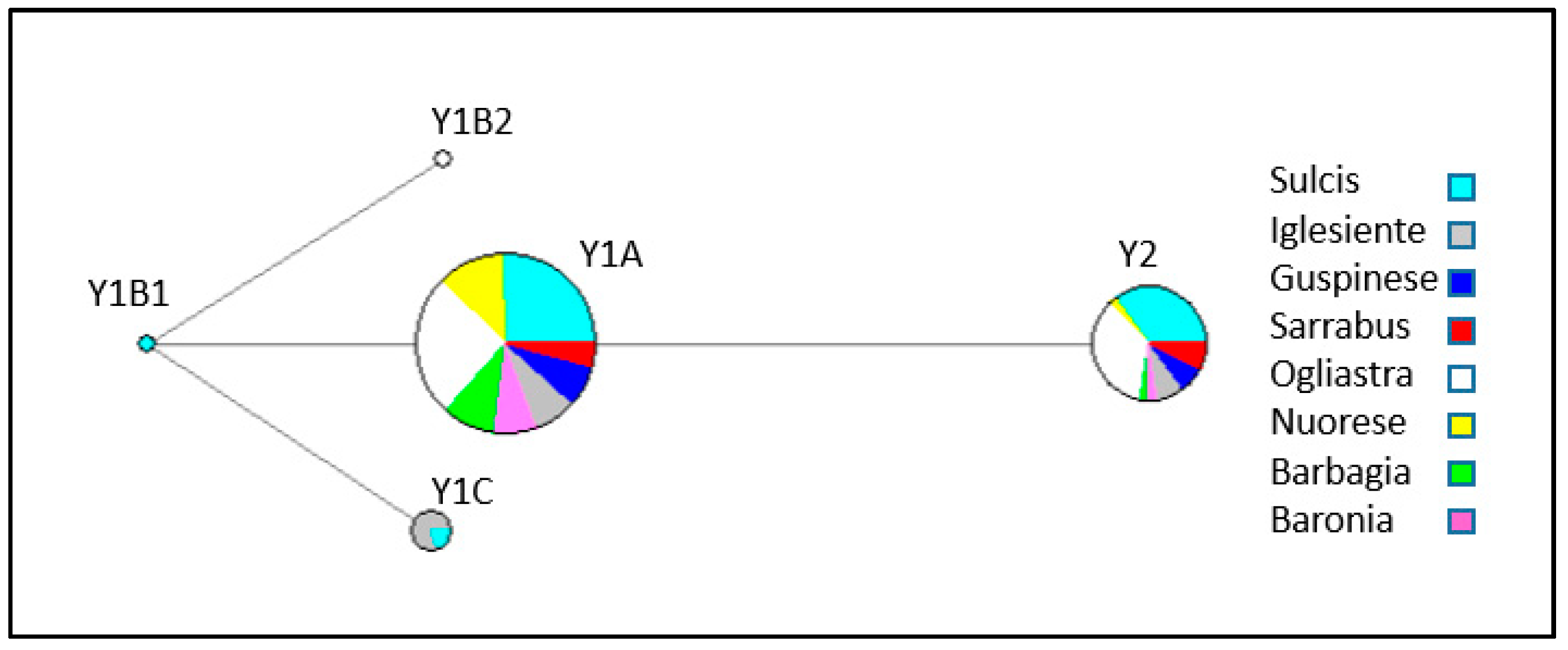
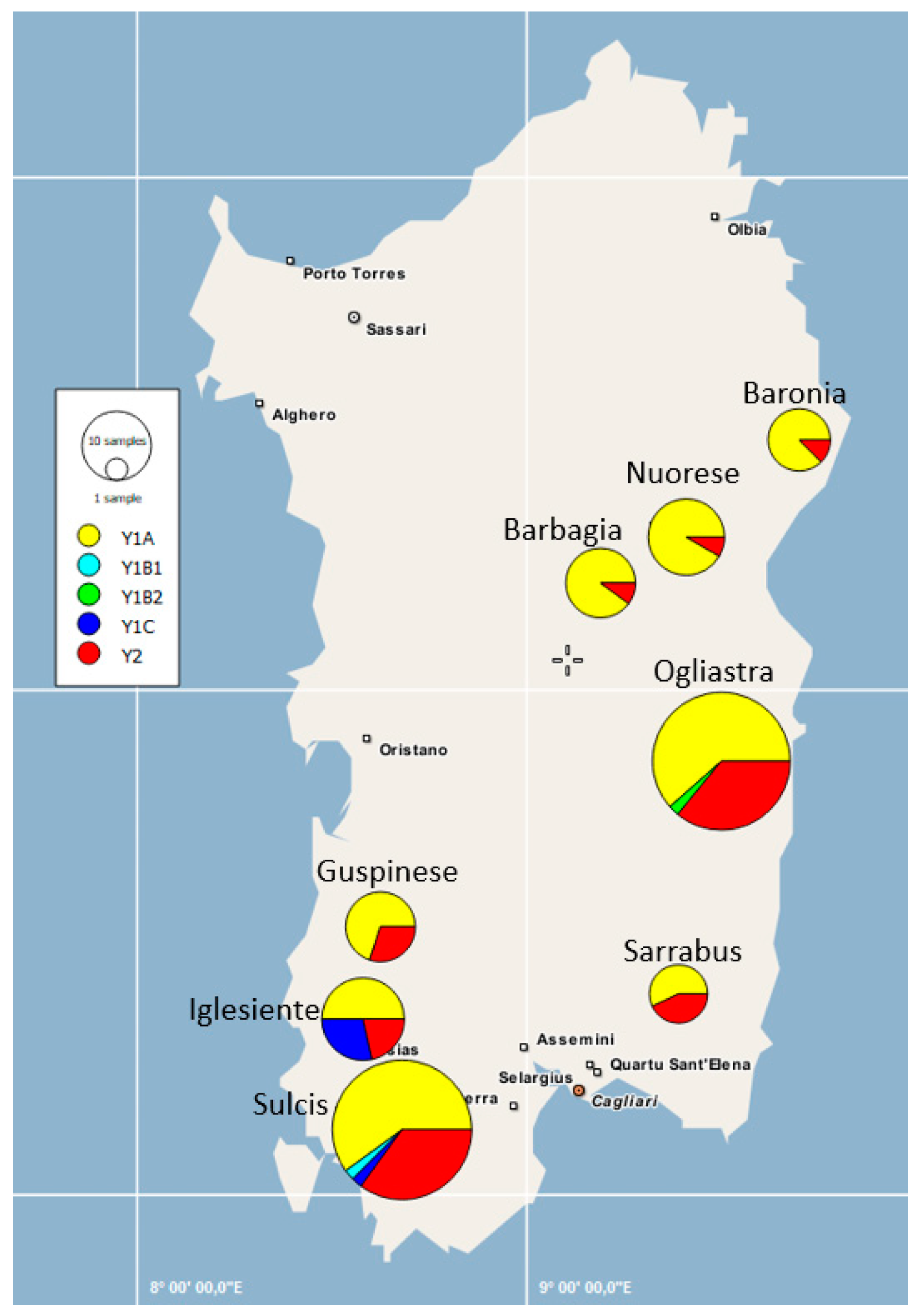
3.1. Y-Chromosome Markers
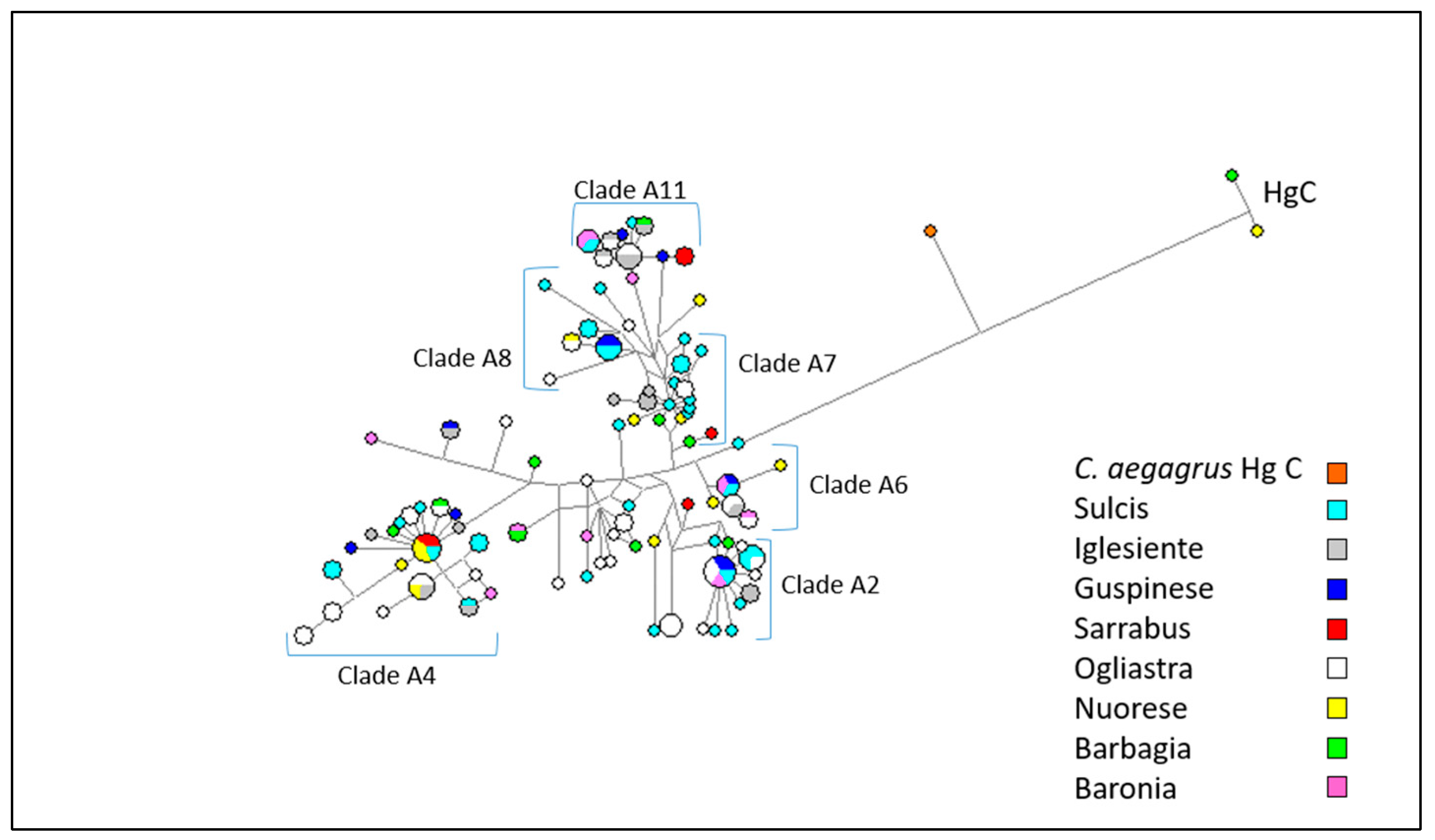
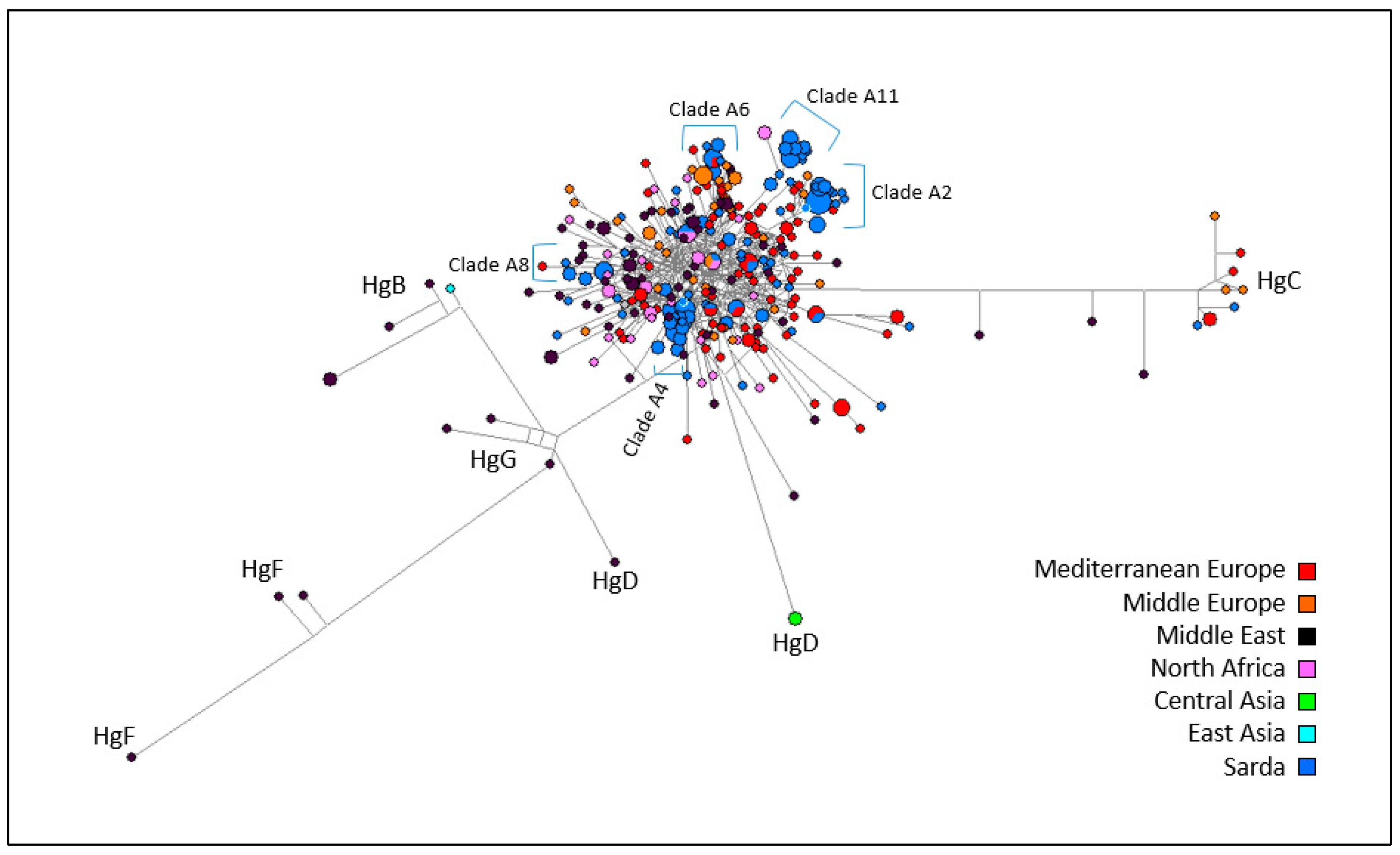
3.2. Mitochondrial DNA Analysis
4. Discussion
4.1. Patterns of Y-Chromosome and Mitochondrial Diversity in Sarda Bucks
4.2. Lack of Population Substructure in the Sarda Breed
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Commission on Genetic Resources for Food and Agricolture. Report of a Consultation on the Definition of Breed Categories. In Proceedings of the Item 5 of the Provisional Agenda, Seventh Session, Rome, Italy, 24–26 October 2012. [Google Scholar]
- Marshall, K. Optimizing the use of breed types in developing country livestock production systems: A neglected research area. J. Anim. Breed. Genet. 2014, 131, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Usai, M.G.; Casu, S.; Molle, G.; Decandia, M.; Ligios, S.; Carta, A. Using cluster analysis to characterize the goat farming system in Sardinia. Livest. Sci. 2006, 104, 63–76. [Google Scholar] [CrossRef]
- Vacca, G.M.; Paschino, P.; Dettori, M.L.; Bergamaschi, M.; Cipolat-Gotet, C.; Bittante, G.; Pazzola, M. Environmental, morphological, and productive characterization of Sardinian goats and use of latent explanatory factors for population analysis. J. Anim. Sci. 2016, 94, 3947–3957. [Google Scholar] [CrossRef] [PubMed]
- Vacca, G.M.; Daga, C.; Pazzola, M.; Carcangiu, V.; Dettori, M.L.; Cozzi, M.C. D-loop sequence mitochondrial DNA variability of Sarda goat and other goat breeds and populations reared in the Mediterranean area. J. Anim. Breed. Genet. 2010, 127, 352–360. [Google Scholar] [CrossRef]
- Piras, D.; Doro, M.G.; Casu, G.; Melis, M.P.; Vaccargiu, S.; Piras, I.; Debora, P.; Stradoni, R.; Frongia, B.; Lai, G.; et al. Haplotype affinities resolve a major component of goat (Capra hircus) mtDNA D-loop diversity and reveal specific features of the Sardinian stock. PLoS ONE 2012, 7, e30785. [Google Scholar] [CrossRef]
- Pazzola, M.; Dettori, M.L.; Vacca, G.M. The Sarda goat, a resource for the extensive exploitation in the Mediterranean environment. In Sustainable Goat Production in Adverse Environments; Simoes, J., Gutierrez, C., Eds.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 181–190. [Google Scholar]
- Vacca, G.M.; Pazzola, M.; Pisano, C.; Carcangiu, V.; Diaz, M.L.; Nieddu, M.; Robledo, R.; Mezzanotte, R.; Dettori, M.L. Chromosomal localisation and genetic variation of the SLC11A1 gene in goats (Capra hircus). Vet. J. 2011, 190, 60–65. [Google Scholar] [CrossRef]
- Vacca, G.M.; Dettori, M.L.; Piras, G.; Manca, F.; Paschino, P.; Pazzola, M. Goat casein genotypes are associated with milk production traits in the Sarda breed. Anim. Genet. 2014, 45, 723–731. [Google Scholar] [CrossRef]
- Usai, M.; Casu, S.; Sechi, T.; Salaris, S.; Miari, S.; Sechi, S.; Carta, P.; Carta, A. Mapping genomic regions affecting milk traits in Sarda sheep by using the OvineSNP50 Beadchip and principal components to perform combined linkage and linkage disequilibrium analysis. GSE 2019, 51, 65. [Google Scholar] [CrossRef]
- Bruford, M.W.; Bradley, D.G.; Luikart, G. DNA markers reveal the complexity of livestock domestication. Nat. Rev. Genet. 2003, 4, 900–910. [Google Scholar] [CrossRef]
- Butler, J.M. Y-Chromosome DNA Testing. In Advanced Topics in Forensic DNA Typing: Methodology; Elsevier Inc.: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Pidancier, N.; Jordan, S.; Luikart, G.; Taberlet, P. Evolutionary history of the genus Capra (Mammalia, Artiodactyla): Discordance between mitochondrial DNA and Y-chromosome phylogenies. Mol. Phylogenet. Evol. 2006, 40, 739–749. [Google Scholar] [CrossRef]
- Cinar Kul, B.; Bilgen, N.; Lenstra, J.A.; Korkmaz Agaoglu, O.; Akyuz, B.; Ertugrul, O.K. Y-chromosomal variation of local goat breeds of Turkey close to the domestication centre. J. Anim. Breed. Genet. 2015, 132, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Waki, A.; Sasazaki, S.; Kobayashi, E.; Mannen, H. Paternal phylogeography and genetic diversity of East Asian goats. Anim. Genet. 2015, 46, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Vidal, O.; Drögemüller, C.; Obexer-Ruff, G.; Reber, I.; Jordana, J.; Martínez, A.; Bâlteanu, V.A.; Delgado, J.V.; Eghbalsaied, S.; Landi, V.; et al. Differential distribution of Y-chromosome haplotypes in Swiss and Southern European goat breed. Sci. Rep. 2017, 7, 16161. [Google Scholar] [CrossRef] [PubMed]
- Luikart, G.; Gielly, L.; Excoffier, L.; Vigne, J.D.; Bouvet, J.; Taberlet, P. Multiple maternal origins and weak phylogeographic structure in domestic goats. Proc. Natl. Acad. Sci. USA 2001, 98, 5927–5932. [Google Scholar] [CrossRef] [PubMed]
- Naderi, S.; Rezaei, H.R.; Taberlet, P.; Zundel, S.; Rafat, S.A.; Naghash, H.R.; El-Barody, M.A.A.; Ertugrul, O.; Pompanon, F.; Econogene Consortium. Large scale mitochondrial DNA analysis of the domestic goat reveals six haplogroups with high diversity. PLoS ONE 2007, 2, e1012. [Google Scholar] [CrossRef] [PubMed]
- Colli, L.; Lancioni, H.; Cardinali, I.; Olivieri, A.; Capodiferro, M.R.; Pellecchia, M.; Rzepus, M.; Zamani, W.; Naderi, S.; Gandini, F.; et al. Whole mitochondrial genomes unveil the impact of domestication on goat matrilineal variability. BMC Genom. 2015, 16, 1115. [Google Scholar] [CrossRef] [PubMed]
- Amills, M.; Capote, J.; Tosser-Klopp, G. Goat domestication and breeding: A jigsaw of historical, biological and molecular data with missing piece. Anim. Genet. 2017, 48, 631–644. [Google Scholar] [CrossRef]
- Sultana, S.; Mannen, H.; Tsuji, S. Mitochondrial DNA diversity of Pakistani goats. Anim. Genet. 2003, 34, 417–421. [Google Scholar] [CrossRef]
- Sardina, M.T.; Ballester, M.; Marmi, J.; Finocchiaro, R.; van Kaam, J.B.C.H.M.; Portolano, B.; Folch, J.M. Phylogenetic analysis of Sicilian goats reveals a new mtDNA lineage. Anim. Genet. 2006, 37, 376–378. [Google Scholar] [CrossRef]
- Naderi, S.; Rezaei, H.R.; Pompanon, F.; Blum, M.G.B.; Negrini, R.; Naghash, H.R.; Balkız, O.; Mashkour, M.; Gaggiotti, O.E.; Ajmone-Marsan, P.; et al. The goat domestication process inferred from large-scale mitochondrial DNA analysis of wild and domestic individuals. Proc. Natl. Acad. Sci. USA 2008, 105, 17659–17664. [Google Scholar] [CrossRef]
- Sechi, T.; Miari, S.; Piras, D.; Crasta, L.; Mulas, G.; Carta, A. Genetic variation of goat Y chromosome in the Sardinian population. Ital. J. Anim. Sci. 2009, 8, 159–161. [Google Scholar] [CrossRef]
- Doro, M.G.; Piras, D.; Leoni, G.G.; Casu, G.; Vaccargiu, S.; Parracciani, D.; Naitana, S.; Pirastu, M.; Novelletto, A. Phylogeny and Patterns of Diversity of Goat mtDNA Haplogroup A Revealed by Resequencing Complete Mitogenomes. PLoS ONE 2014, 9, e95969. [Google Scholar] [CrossRef] [PubMed]
- Carcangiu, V.; Vacca, G.M.; Parmeggiani, A.; Mura, M.C.; Bini, P.P. Blood melatonin levels relating to the reproductive activity of sarda does. Small Rum. Res. 2005, 59, 7–13. [Google Scholar] [CrossRef]
- Pazzola, M.; Dettori, M.L.; Carcangiu, V.; Luridiana, S.; Mura, M.C.; Vacca, G.M. Relationship between milk urea, blood plasma urea and body condition score in primiparous browsing goats with different milk yield level. Arch. Tierz. 2011, 54, 546–556. [Google Scholar] [CrossRef][Green Version]
- Lenstra, J.A. Evolutionary and demographic history of sheep and goats suggested by nuclear mtDNA and Y-chromosome markers. In Proceedings of the International Workshop on the Role of Biotechnology for the Characterization of Crop, Forestry, Animal and Fishery Genetic Resources, Turin, Piemonte, Italy, 5–7 March 2005; pp. 97–100. [Google Scholar]
- Pereira, F.; Carneiro, J.; Soares, P.; Maciel, S.; Nejmeddine, F.; Lenstra, J.A.; Gusmão, L.; Amorim, A. A multiplex primer extension assay for the rapid identification of paternal lineages in domestic goat (Capra hircus): Laying the foundations for a detailed caprine Y chromosome phylogeny. Mol. Phylogenet. Evol. 2008, 49, 663–668. [Google Scholar] [CrossRef]
- Servei Veterinari de Genetica Molecular, Universitat Autònoma de Barcelona. Available online: http://sct.uab.cat/svgm/en (accessed on 9 October 2020).
- Martinez, A.; Ferrando, A.; Manunza, A.; Gomez, M.; Landi, V.; Jordana, J.; Capote, J.; Badaoui, B.; Vidal, O.; Delgado, J.V.; et al. Inferring the demographic history of a highly endangered goat breed through the analysis of nuclear and mitochondrial genetic signatures. Small Rum. Res. 2012, 104, 78–84. [Google Scholar] [CrossRef]
- Amills, M.; Capote, J.; Tomàs, A.; Kelly, L.; Obexer-Ruff, G.; Angiolillo, A.; Sanchez, A. Strong phylogeographic relationships among three goat breeds from the Canary Islands. J. Dairy Res. 2004, 71, 257–262. [Google Scholar] [CrossRef]
- Ewing, L.; Green, P. Base-calling of automated sequencer traces using phred. II. Error probabilities. Genome Res. 1998, 8, 186–194. [Google Scholar] [CrossRef]
- Ewing, L.; Hillier, L.; Wendl, M.C.; Green, P. Base-calling of automated sequencer traces using phred I. Accuracy assessment. Genome Res. 1998, 8, 175–185. [Google Scholar] [CrossRef]
- Nickerson, D.A.; Tobe, V.O.; Taylor, S.L. PolyPhred: Automating the detection and genotyping of single nucleotide substitutions using fluorescence-based resequencing. Nucleic Acids Res. 1997, 25, 2745–2751. [Google Scholar] [CrossRef]
- Bhangale, T.R.; Stephens, M.; Nickerson, D.A. Automating resequencing based detection of insertion-deletion polymorphisms. Nat. Genet. 2006, 38, 1457–1462. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Abajian, C.; Green, P. Consed: A graphical tool for sequence finishing. Genome Res. 1998, 8, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Desmarais, C.; Green, P. Automated finishing with autofinish. Genome Res. 2001, 11, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D. Viewing and editing assembled sequences using Consed. In Current Protocols in Bioinformatics; Baxevanis, D., Davison, D.B., Eds.; Wiley: New York, NY, USA, 2004; pp. 1121–1124. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DNAspv5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Hudson, R.R.; Slatkin, M.; Maddison, W.P. Estimation of levels of gene flow from DNA sequence data. Genetics 1992, 132, 583–589. [Google Scholar]
- Network Software v.10. Available online: www.fluxus-engineering.com (accessed on 3 October 2020).
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Hassanin, A.; Bonillo, C.; Nguyen, B.X.; Cruaud, C. Comparisons between mitochondrial genomes of domestic goat (Capra hircus) reveal the presence of numts and multiple sequencing errors. Mitochondrial DNA 2010, 21, 68–76. [Google Scholar] [CrossRef]
- Paschino, P.; Stocco, G.; Dettori, M.L.; Pazzola, M.; Marongiu, M.L.; Pilo, C.E.; Cipolat-Gotet, C.; Vacca, G.M. Characterization of milk composition, coagulation properties and cheese-making ability of goats reared in extensive farms. J. Dairy Sci. 2020, 103, 5830–5843. [Google Scholar] [CrossRef]
- Mattone, A.; Lilliu, G. La Costante Resistenziale Sarda; Ilisso: Nuoro, Italy, 2002. [Google Scholar]
- Brown, P.J. Cultural adaptations to endemic malaria in Sardinia. Med. Anthropol. 1981, 5, 313–339. [Google Scholar] [CrossRef]
- Colli, L.; Milanesi, M.; Talenti, A.; Bertolini, F.; Chen, M.; Crisà, A.; Daly, K.G.; Del Corvo, M.; Guldbrandtsen, B.; Lenstra, J.A.; et al. Genome-wide SNP profiling of worldwide goat populations reveals strong partitioning of diversity and highlights post-domestication migration routes. Genet. Sel. Evol. 2018, 50, 58. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, T.F.; Amills, M.; Bertolini, F.; Rothschild, M.; Marras, G.; Boink, G.; Jordana, J.; Capote, J.; Carolan, S.; Hallsson, J.H.; et al. Patterns of homozygosity in insular and continental goat breeds. Genet. Sel. Evol. 2018, 50, 56. [Google Scholar] [CrossRef] [PubMed]
- Abulafia, D. The Mediterranean in History; Getty Publications: Los Angeles, CA, USA, 2011. [Google Scholar]
- Porter, V.; Alderson, L.; Hall, S.J.G.; Sponenberg, D.P. (Eds.) Mason’s World Encyclopedia of Livestock Breeds and Breeding; CABI: Wallingford, UK, 2016; pp. 343–420. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haplotype | SRY 2971 | SRY 3098 | SRY 1876 | AMELY 42 | ZFY 527 | ZFY 46 | DDX3Y 56 | Haplotype Frequency% |
|---|---|---|---|---|---|---|---|---|
| Y1A | T | G | A | C | A | C | G | 66.44 |
| Y2 | T | A | A | C | G | C | C | 28.57 |
| Y1C | A | G | C | C | A | C | G | 3.57 |
| Y1B1 | A | G | A | C | A | C | G | 0.71 |
| Y1B2 | A | G | A | C | A | T | G | 0.71 |
| Region | Number of Haplotypes | Haplotype Diversity | Number of Animals |
|---|---|---|---|
| Nuorese | 2 | 0.166 | 12 |
| Barbagia | 2 | 0.200 | 10 |
| Baronia | 2 | 0.250 | 8 |
| Ogliastra | 3 | 0.504 | 39 |
| Sarrabus | 2 | 0.571 | 7 |
| Guspinese | 2 | 0.466 | 10 |
| Iglesiente | 3 | 0.670 | 14 |
| Sulcis | 4 | 0.529 | 40 |
| Overall population | 5 | 0.479 | 140 |
| Heading | Sulcis | Guspinese | Sarrabus | Ogliastra | Nuorese | Baronia | Barbagia |
|---|---|---|---|---|---|---|---|
| Guspinese | 0.00 | ||||||
| Sarrabus | 0.00 | 0.00 | |||||
| Ogliastra | 0.00 | 0.00 | 0.00 | ||||
| Nuorese | 0.14 | 0.05 | 0.16 | 0.16 | |||
| Baronia | 0.07 | 0.00 | 0.08 | 0.08 | 0.00 | ||
| Barbagia | 0.12 | 0.02 | 0.13 | 0.13 | 0.00 | 0.00 | |
| Iglesiente | 0.05 | 0.03 | 0.05 | 0.06 | 0.09 | 0.05 | 0.07 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Among populations | 7 | 6.828 | 0.01755 Va | 2.47% |
| Within populations | 132 | 91.350 | 0.69205 Vb | 97.53% |
| Total | 139 | 98.179 | 0.70960 | |
| Fixation Index FST: 0.02473 | p-value = 0.18 |
| Region | Nucleotide Diversity, Π | Number of Haplotypes | Haplotype Diversity, Hd | Sample Size |
|---|---|---|---|---|
| Nuorese | 0.017 | 11 | 0.984 | 12 |
| Barbagia | 0.020 | 10 | 1.000 | 10 |
| Baronia | 0.015 | 9 | 0.977 | 10 |
| Ogliastra | 0.013 | 30 | 0.984 | 42 |
| Sarrabus | 0.012 | 4 | 0.866 | 6 |
| Guspinese | 0.013 | 8 | 0.955 | 10 |
| Iglesiente | 0.013 | 13 | 0.980 | 15 |
| Sulcis | 0.012 | 32 | 0.989 | 39 |
| Overall population | 0.013 | 94 | 0.992 | 144 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Among populations | 7 | 40.850 | −0.02873 Va | −0.46% |
| Within populations | 136 | 858.553 | 6.31289 Vb | 100.46% |
| Total | 143 | 899.403 | 6.28416 | |
| Fixation Index FST: −0.00457 | p-value = 0.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dettori, M.L.; Petretto, E.; Pazzola, M.; Vidal, O.; Amills, M.; Vacca, G.M. Assessing the Diversity and Population Substructure of Sarda Breed Bucks by Using Mtdna and Y-Chromosome Markers. Animals 2020, 10, 2194. https://doi.org/10.3390/ani10122194
Dettori ML, Petretto E, Pazzola M, Vidal O, Amills M, Vacca GM. Assessing the Diversity and Population Substructure of Sarda Breed Bucks by Using Mtdna and Y-Chromosome Markers. Animals. 2020; 10(12):2194. https://doi.org/10.3390/ani10122194
Chicago/Turabian StyleDettori, Maria Luisa, Elena Petretto, Michele Pazzola, Oriol Vidal, Marcel Amills, and Giuseppe Massimo Vacca. 2020. "Assessing the Diversity and Population Substructure of Sarda Breed Bucks by Using Mtdna and Y-Chromosome Markers" Animals 10, no. 12: 2194. https://doi.org/10.3390/ani10122194
APA StyleDettori, M. L., Petretto, E., Pazzola, M., Vidal, O., Amills, M., & Vacca, G. M. (2020). Assessing the Diversity and Population Substructure of Sarda Breed Bucks by Using Mtdna and Y-Chromosome Markers. Animals, 10(12), 2194. https://doi.org/10.3390/ani10122194