Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias latipes)

 ,
,  ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Broodstock
2.2. Experimental Diet
2.3. Evaluation of the Reproductive Effect
2.3.1. Sperm
2.3.2. COX Activity
2.3.3. Immunohistochemistry Detection of COX2
2.3.4. Eggs
2.4. Larvae
2.5. Statistical Analysis
3. Results
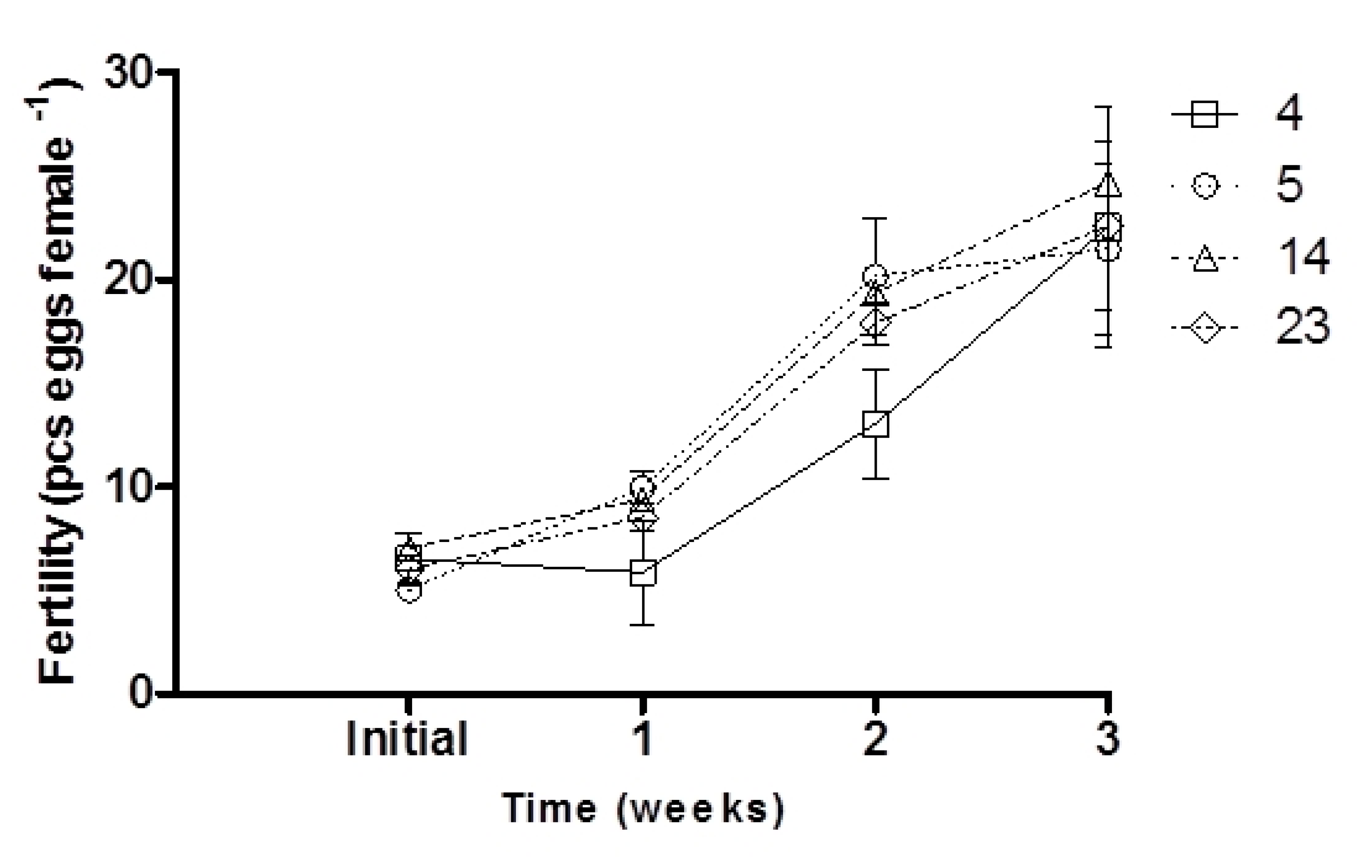
3.1. Effect of the ARA Diets on Broodstock and Gametes
3.2. Effect of the ARA Diets on Embryos
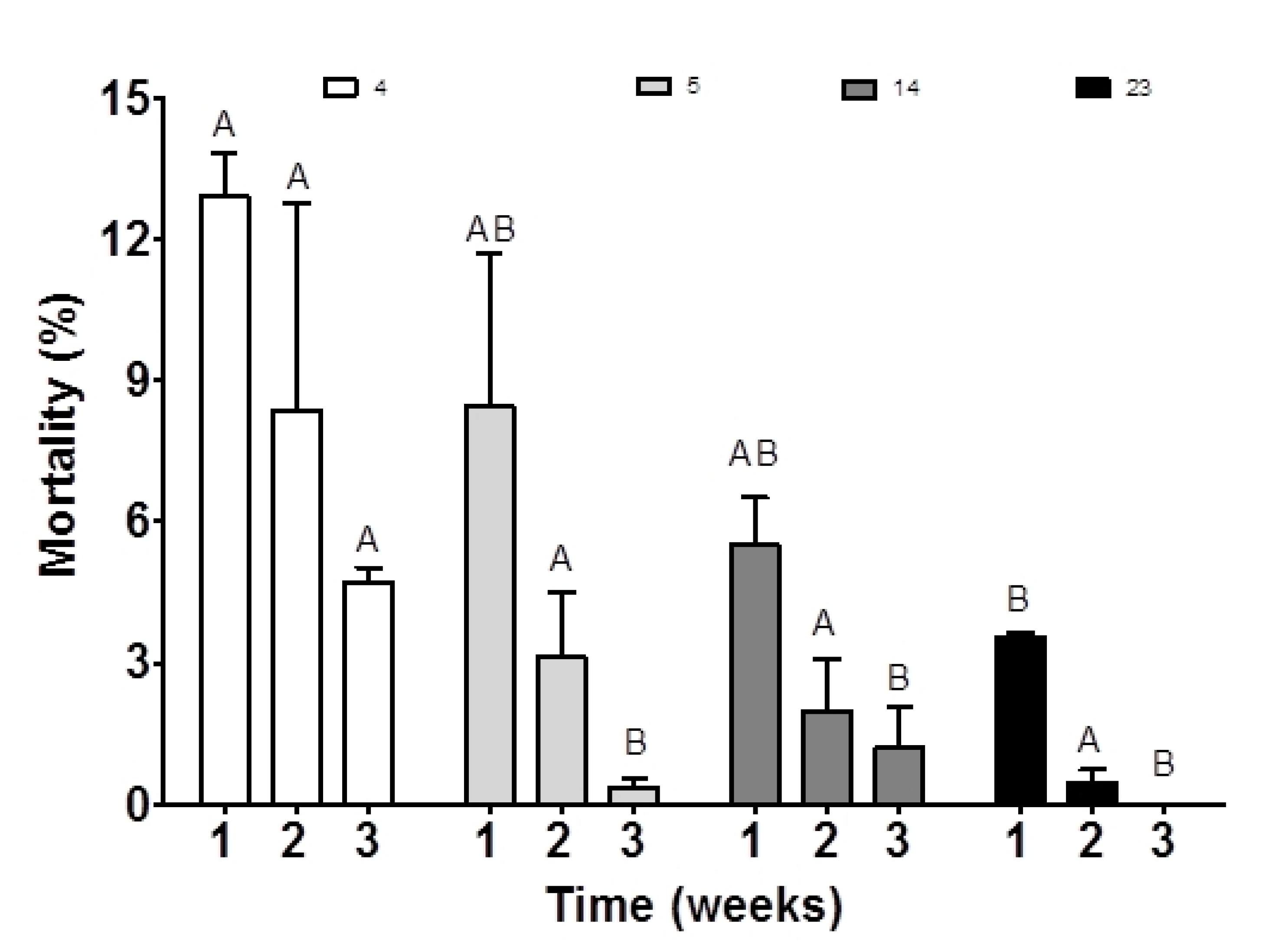
3.3. Effect of the ARA Diet on Larval Quality
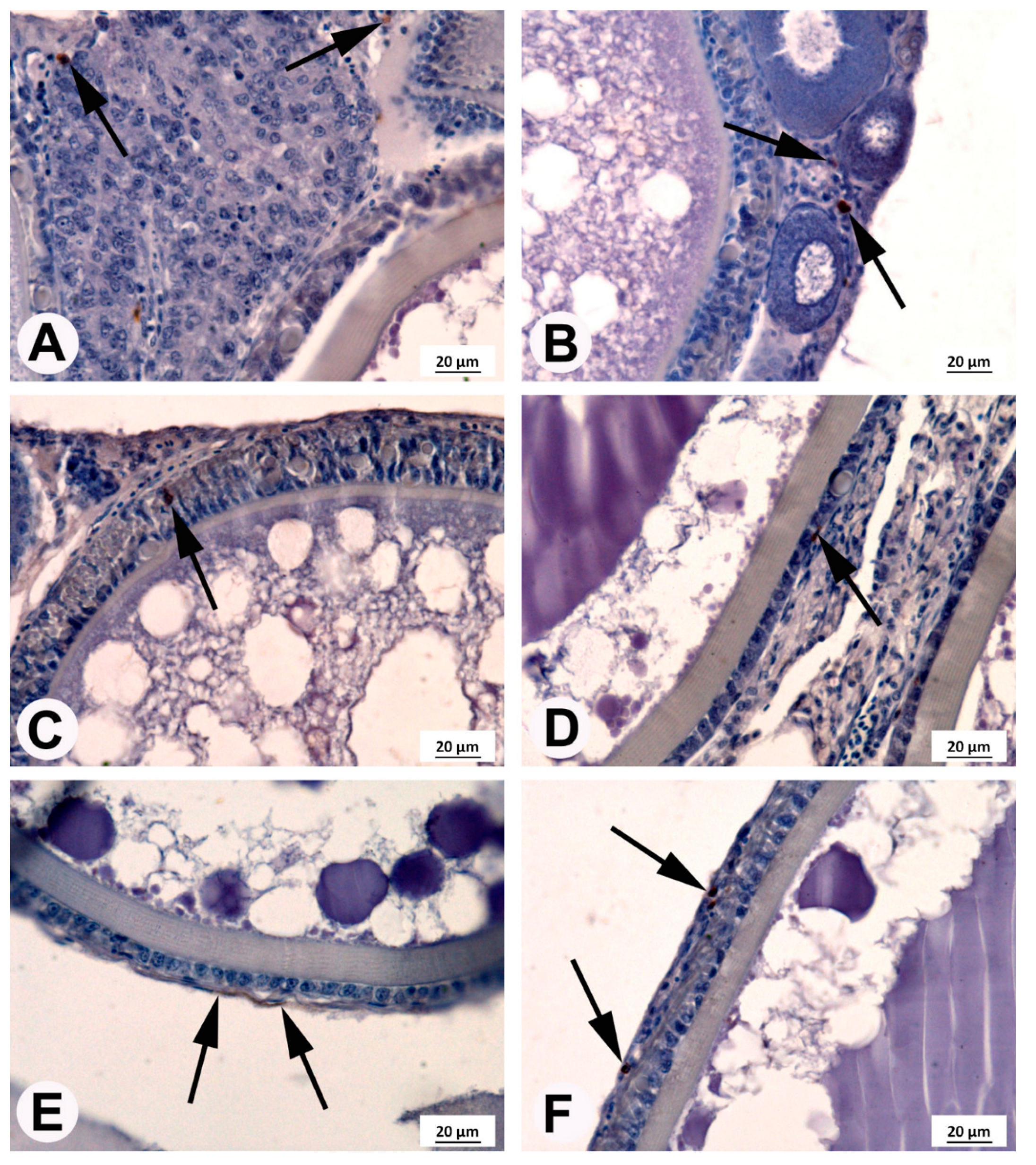
3.4. Immunohistochemical Localization of COX2
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Ballestrazzi, R.; Rainis, S.; Tulli, F.; Bracelli, A. The effect of dietary coconut oil on reproductive traits and egg fatty acid composition in rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2003, 11, 289–299. [Google Scholar] [CrossRef]
- Kowalska, A.; Zakęś, Z.; Jankowska, B.; Siwicki, A. Impact of diets with vegetable oils on the growth, histological structure of internal organs, biochemical blood parameters, and proximate composition of pikeperch Sander lucioperca (L.). Aquaculture 2010, 301, 69–77. [Google Scholar] [CrossRef]
- Kowalska, A.; Zakęś, Z.; Jankowska, B.; Siwicki, A. Substituting vegetable oil for fish oil in pikeperch diets: The impact on growth, internal organ histology, blood biochemical parameters, and proximate composition. Aquac. Nutr. 2011, 17, e148–e163. [Google Scholar] [CrossRef]
- Kowalska, A.; Zakęś, Z.; Siwicki, A.; Jankowska, B.; Jarmołowicz, S.; Demska-Zakęś, K. Impact of diets with different proportions of linseed and sunflower oils on the growth, liver histology, immunological and chemical blood parameters, and proximate composition of pikeperch Sander lucioperca (L.). Fish Physiol. Biochem. 2012, 38, 375–388. [Google Scholar] [CrossRef]
- Parpoura, A.C.R.; Alexis, M.N. Effect of different dietary oils in sea bass (Dicentrarchus labrax) nutrition. Aquac. Int. 2001, 9, 463–476. [Google Scholar] [CrossRef]
- Bell, J.G.; Sargent, J.R. Arachidonic acid in aquaculture feeds: Current status and future opportunities. Aquaculture 2003, 218, 491–499. [Google Scholar] [CrossRef]
- Miguel, A.H.; Reza, M.; Ibarra, A.M.; Wille, M.; Sorgeloos, P.; Soudant, P.; Palacios, E. Arachidonic acid (20:4n−6) effect on reproduction, immunology, and prostaglandin E2 levels in Crassostrea corteziensis (Hertlein, 1951). Aquaculture 2009, 294, 300–305. [Google Scholar]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Tocher, D.R.; Bell, J.G.; McGhee, F.; Dick, J.R.; Foneca–Madrigal, J. Effect of dietary lipid level and vegetable oil on fatty acid metabolism in Atlantic salmon (Salmo salar L.) over the whole production cycle. Fish Physiol. Biochem. 2003, 29, 193–209. [Google Scholar] [CrossRef]
- Nath, P.R.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S.; Zilberg, D. Dietary supplementation with the microalgae Parietochloris incisa increases survival and stress resistance in guppy (Poecilia reticulata) fry. Aquac. Nutr. 2012, 18, 167–180. [Google Scholar] [CrossRef]
- Wang, N.; Mandiki, S.N.M.; Henrotte, E.; Bouyahia, A.B.; Mairesse, G.; Rougeot, C.; Melard, C.; Kestemont, P. Effect of partial or total replacement of forage fish by a dry diet on quality of reproduction in pikeperch, Sander lucioperca. Aquac. Res. 2009, 40, 376–383. [Google Scholar] [CrossRef]
- Fujimori, C.; Ogiwara, K.; Hagiwara, A.; Rajapakse, S.; Kimura, A.; Takahashi, T. Expression of cyclooxygenase-2 and prostaglandin receptor EP4b mRNA in the ovary of the medaka fish, Oryzias latipes: Possible involvement in ovulation. Mol. Cell. Endocrinol. 2011, 332, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The Lipids. In Fish Nutrition; Halver, J.E., Hardy, R., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 182–246. [Google Scholar]
- Kinoshita, M.; Murata, K.; Naruse, K.; Tanaka, M. Medaka management. In Medaka. Biology, Management, and Experimental Protocols; Kinoshita, M., Murata, K., Naruse, K., Tanaka, M., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 31–65. [Google Scholar]
- Yang, H.; Tiersch, T.R. Sperm motility initiation and duration in a euryhaline fish, medaka (Oryzias latipes). Theriogenology 2009, 72, 386–392. [Google Scholar] [CrossRef]
- Folch, H.; Less, M.; Stanley, H.A. A simple method for isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497. [Google Scholar]
- Peisker, K. Rapid semi-micro method for methyl esters from triglycerides using chloroform, methanol, sulphuric acid. J. Am. Oil Chem. Soc. 1964, 11, 87–90. [Google Scholar] [CrossRef]
- Iwamatsu, T.; Ohta, T.; Oshima, E.; Sakai, N. Oogenesis in the medaka Oryzias latipes-stages of oocyte development. Zool. Sci. 1988, 5, 353–373. [Google Scholar]
- Tocher, D.R.; Bell, J.G.; Sargent, J.R. Production of eicosanoids derived from 20:4n-6 and 20:5n-3 in primary cultures of turbot (Scophthalmus maximus) brain astrocytes in response to platelet activating factor, substance P and interleukin-1 beta. Comp. Biochem. Physiol. Biochem. Mol. Biol. 1996, 15, 215–222. [Google Scholar] [CrossRef]
- Kowalska, A.; Kowalski, R.K. The effect of cyclooxygenase (COX) inhibitors on Japanese medaka (Oryzias latipes) reproduction parameters fed with high level of arachidonic acid (20:4 n-6). Aquac. Int. 2014, 22, 185–193. [Google Scholar] [CrossRef]
- Kowalska, A.; Kowalski, R.K.; Zakęś, Z. The effect of selective cyclooxygenase (COX) inhibitors on Japanese medaka (Oryzias latipes) reproduction parameters. World Acad. Sci. Eng. Technol. 2011, 77, 19–23. [Google Scholar]
- Gage, M.J.G.; Macfarlane, C.P.; Yeates, S.; Ward, R.G.; Searle, J.B.; Parker, G.A. Spermatozoal traits and sperm competition in Atlantic salmon: Relative sperm velocity is the primary determinant of fertilization success. Curr. Biol. 2004, 14, 44–47. [Google Scholar] [CrossRef]
- Liu, W.; Cao, D.; Oh, S.F.; Serhan, C.N.; Kulmacz, R.J. Divergent cyclooxygenase responses to fatty acid structure and peroxide level in fish and mammalian prostaglandin H synthases. FASEB J. 2006, 20, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Furne, M.; Holen, E.; Araujo, P.; Lie, K.; Moren, M. Cytokine gene expression and prostaglandin production in head kidney leukocytes isolated from Atlantic cod (Gadus morhua) added different levels of arachidonic acid and eicosapentaenoic acid. Fish. Shelfish Immunol. 2013, 34, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.P.; Havird, J.; Rose, R.; Hyndman, K.; Piermarini, P.; Evans, D.H. COX2 in a euryhaline teleost, Fundulus heteroclitus: Primary sequence, distribution, localization, and potential function in gills during salinity acclimation. J. Exp. Biol. 2006, 209, 1696–1708. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, A.; Siwicki, A.K.; Kowalski, R.K. Dietary resveratrol improves immunity but reduces reproduction of broodstock medaka Oryzias latipes (Temminck & Schlegel). Fish. Physiol. Biochem. 2017, 43, 27–37. [Google Scholar]
- Petersen, R.K.; Jørgensen, C.; Rustan, A.C.; Frøyland, L.; Muller-Decker, K.; Furstenberger, G.; Berge, R.K.; Kristiansen, K.; Madsen, L. Arachidonic acid-dependent inhibition of adipocyte differentiation requires PKA activity and is associated with sustained expression of cyclooxygenases. J. Lipid Res. 2003, 44, 2320–2330. [Google Scholar] [CrossRef]
- Hales, D.B.; Zhuge, Y.; Lagman, J.A.J.; Ansenberger, K.; Mahon, C.; Barua, A.; Luborsky, J.L.; Bahr, J.M. Cyclooxygenases expression and distribution in the normal ovary and their role in ovarian cancer in the domestic hen (Gallus domesticus). Endocrine 2008, 33, 235–244. [Google Scholar] [CrossRef]
- Richards, J.S.; Russell, D.L.; Ochsner, S.; Espey, L.L. Ovulation: New dimensions and new regulators of the inflammatory-like response. Annu. Rev. Physiol. 2002, 64, 69–92. [Google Scholar] [CrossRef]
- Dubois, R.N.; Abramson, S.B.; Crofford, L.; Gupta, R.A.; Simon, L.S.; Van De Putte, L.B.; Lipsky, P.E. Cyclooxygenase in biology and disease. FASEB J. 1998, 12, 1063–1073. [Google Scholar] [CrossRef]
- Martins, D.A.; Rocha, F.; Castanheira, F.; Mendes, A.; Pousão-Ferreira, P.; Bandarra, N.; Coutinho, J.; Morais, S.; Yúfera, M.; Conceição, L.E.; et al. Effects of dietary arachidonic acid on cortisol production and gene expression in stress response in Senegalese sole (Solea senegalensis) post-larvae. Fish Physiol. Biochem. 2013, 39, 1223–1238. [Google Scholar] [CrossRef]
- Martins, D.A.; Rocha, F.; Martínez-Rodríguez, G.; Bell, G.; Morais, S.; Castanheira, F.; Bandarra, N.; Coutinho, J.; Yúfera, M.; Conceição, L.E.C. Teleost fish larvae adapt to dietary arachidonic acid supply through modulation of the expression of lipid metabolism and stress response genes. Br. J. Nutr. 2011, 108, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.I.; Kim, S.H.; Sepich, D.; Buchanan, F.G.; Solnica-Krezel, L.; DuBois, R.N. Cyclooxygenase-1-derived PGE2 promotes cell motility via the G-protein-coupled EP4 receptor during vertebrate gastrulation. Genes Dev. 2006, 20, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Burdan, F. Comparison of developmental toxicity of selective and nonselective cyclooxygenase-2 inhibitors in CRL: (WI) WUBR Wistar rats—DFU and piroxicam study. Toxicology 2005, 211, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Walsh, L.P.; Reinhart, A.J.; Stocco, D.M. The role of arachidonic acid in steroidogenesis and steroidogenic acute regulatory (StAR) gene and protein expression. J. Biol. Chem. 2000, 275, 20204–20209. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Dyson, M.T.; Jo, Y.; Eubank, D.W.; Stocco, D.M. Involvement of 5-lipoxygenase metabolites of arachidonic acid in cyclic AMP-stimulated steroidogenesis and steroidogenic acute regulatory protein gene expression. J. Steroid Biochem. Mol. Biol. 2003, 85, 159–166. [Google Scholar] [CrossRef]
- Meves, H. Arachidonic acid and ion channels: An update. Br. J. Pharmacol. 2008, 155, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Gammanpila, M.; Akupitiyage, A.Y.; Bart, A.N. Evaluation of the effect of dietary vitamin C, E and Zinc supplementation on reproductive performance of Nile tilapia (Oreochromis niloticus). J. Aquacult. Sci. 2007, 12, 39–60. [Google Scholar] [CrossRef]
- Koven, W.; Van Anholt, R.D.; Lutsky, S.; Ben Atia, I.; Nixon, O.; Ron, B.; Tandler, A. The effect of dietary arachidonic acid on growth, survival, and cortisol levels in different-age gilthead seabream larvae (Sparus auratus) exposed to handling or daily salinity change. Aquaculture 2003, 228, 307–320. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids mg FA g−1 of total FA | Experimental Diets a | |||
|---|---|---|---|---|
| 4 | 5 | 14 | 23 | |
| C14:0 | 30.45 | 30.19 | 29.33 | 29.28 |
| C16:0 | 171.16 | 168.82 | 165.76 | 165.14 |
| Σ SFA b | 372.21 | 365.35 | 363.18 | 358.86 |
| C16:1 | 31.68 | 31.73 | 30.83 | 30.70 |
| C18:1 n-9 | 183.21 | 178.89 | 175.79 | 171.81 |
| Σ MUFA c | 275.08 | 271.80 | 267.32 | 264.19 |
| C18:2 n-6 | 219.24 | 226.33 | 219.19 | 222.62 |
| C18:3 n-3 | 24.27 | 24.91 | 24.19 | 24.34 |
| C20:4 n-6 | 4.37 | 5.16 | 13.69 | 22.89 |
| C20:4 n-3 | 3.65 | 3.74 | 3.67 | 3.75 |
| C20:5 n-3 | 37.26 | 38.48 | 37.91 | 38.49 |
| C22:5 n-3 | 8.99 | 9.00 | 8.99 | 9.15 |
| C22:6 n-3 | 46.52 | 46.48 | 49.96 | 47.45 |
| ΣPUFA d | 352.71 | 362.85 | 369.50 | 376.95 |
| Σ n-3 LC-PUFA e | 96.42 | 97.69 | 100.52 | 98.84 |
| EPA/ARA | 8.53 | 7.46 | 3.61 | 1.68 |
| DHA/ARA | 10.64 | 9.01 | 3.65 | 2.07 |
| EPA + DHA/ARA | 19.16 | 16.47 | 6.42 | 3.75 |
| Dietary Treatments (mg ARA−g−1 TFA) 1 | ||||
|---|---|---|---|---|
| 4 | 5 | 14 | 23 | |
| Broodstock | ||||
| Body weight (BW, g) | 0.45 ± 0.07 a | 0.41 ± 0.08 a | 0.42 ± 0.07 a | 0.43 ± 0.08 a |
| Body length (TL, cm) | 3.35 ± 0.29 a | 3.34 ± 0.24 a | 3.36 ± 0.19 a | 3.40 ± 0.36 a |
| Fertility (pcs. egg female−1) | 22.54 ± 5.76 a | 21.47 ± 4.10 a | 24.70 ± 6.33 a | 22.6 ± 4.12 a |
| Cyclooxygenase activity (COX1, U mg protein−1) | 10.2 ± 4.2 a | 11.8 ± 3.8 a | 14.5 ± 5.9 b | 13.8 ± 3.6 b |
| Size of oocyte (mm) | 1.20 ± 0.01 a | 1.21 ± 0.02 a | 1.22 ± 0.01 a | 1.21 ± 0.01 a |
| Embryo survival (%) | 62.01 ± 10.97 a | 68.36 ± 6.65 ab | 65.29 ± 3.24 ab | 83.17 ± 5.27 b |
| Hepatosomatic index (HSI, %) | 6.76 ± 1.05 a | 7.05 ± 1.16 a | 7.27 ± 1.75 a | 6.25 ± 1.26 a |
| Larvae | ||||
| Hatching rate (%) | 50.24 ± 5.36 a | 58.32 ± 5.42 ab | 75.16 ± 7.23 b | 78.58 ± 12.15 b |
| Initial body weight (BW, mg) | 0.40 ± 0.03 a | 0.45 ± 0.12 ab | 0.45 ± 0.21 ab | 0.66 ± 0.10 b |
| Initial body length (TL, mm) | 4.73 ± 0.04 a | 4.86 ± 0.09 a | 4.88 ± 0.03 a | 4.87 ± 0.09 a |
| Final body weight (BW, mg) | 2.80 ± 0.78 a | 4.59 ± 0.94 ab | 7.60 ± 1.13 c | 5.96 ± 0.59 bc |
| Final body length (TL, mm) | 7.98 ± 0.69 a | 8.97 ± 0.85 a | 8.75 ± 0.61 a | 8.49 ± 0.51 a |
| Total mortality (%) | 20.39 ± 5.39 b | 9.34 ± 4.78 a | 9.47 ± 2.04 a | 4.04 ± 1.93 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, A.; Kamaszewski, M.; Czarnowska-Kujawska, M.; Podlasz, P.; Kowalski, R.K. Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias latipes). Animals 2020, 10, 2174. https://doi.org/10.3390/ani10112174
Kowalska A, Kamaszewski M, Czarnowska-Kujawska M, Podlasz P, Kowalski RK. Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias latipes). Animals. 2020; 10(11):2174. https://doi.org/10.3390/ani10112174
Chicago/Turabian StyleKowalska, Agata, Maciej Kamaszewski, Marta Czarnowska-Kujawska, Piotr Podlasz, and Radosław K. Kowalski. 2020. "Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias latipes)" Animals 10, no. 11: 2174. https://doi.org/10.3390/ani10112174
APA StyleKowalska, A., Kamaszewski, M., Czarnowska-Kujawska, M., Podlasz, P., & Kowalski, R. K. (2020). Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias latipes). Animals, 10(11), 2174. https://doi.org/10.3390/ani10112174