Single Layer Centrifugation Improves the Quality of Fresh Donkey Semen and Modifies the Sperm Ability to Interact with Polymorphonuclear Neutrophils

 ,
,  ,
,  , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Flow Cytometry
2.3.1. Evaluation of Sperm Membrane Integrity (SYBR14/PI)
2.3.2. Evaluation of Sperm Membrane Lipid Disorder (M540/YO-PRO-1)
2.3.3. Evaluation of Mitochondrial Membrane Potential (JC1)
2.3.4. Evaluation of Intracellular Calcium Levels (Fluo3/PI and Rhod5/YO-PRO-1)
2.3.5. Evaluation of Intracellular Reactive Oxygen Species (ROS) Levels: H2O2 and O2−● (H2DCFDA/PI and HE/YO-PRO-1)
2.4. Evaluation of Sperm Motility
2.5. Evaluation of Sperm–PMN Binding
2.6. Statistical Analysis
3. Results
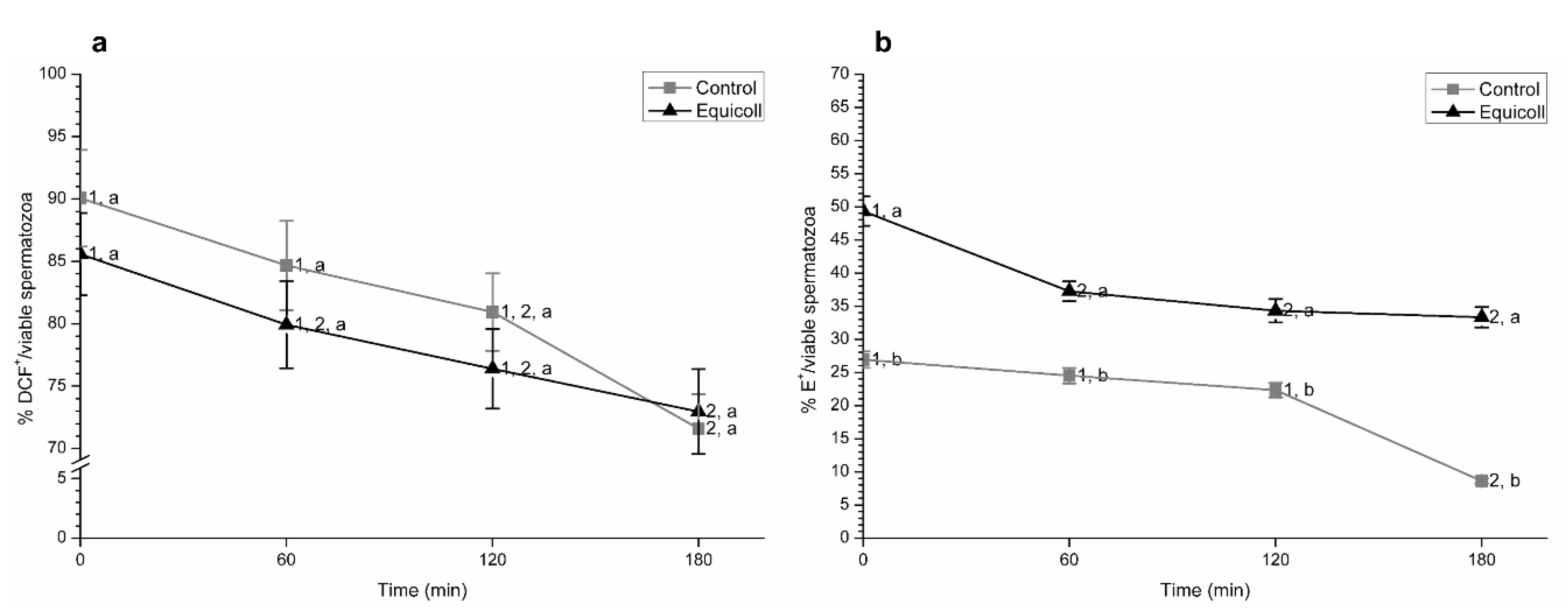
3.1. Integrity and Lipid Disorder of Sperm Plasma Membrane
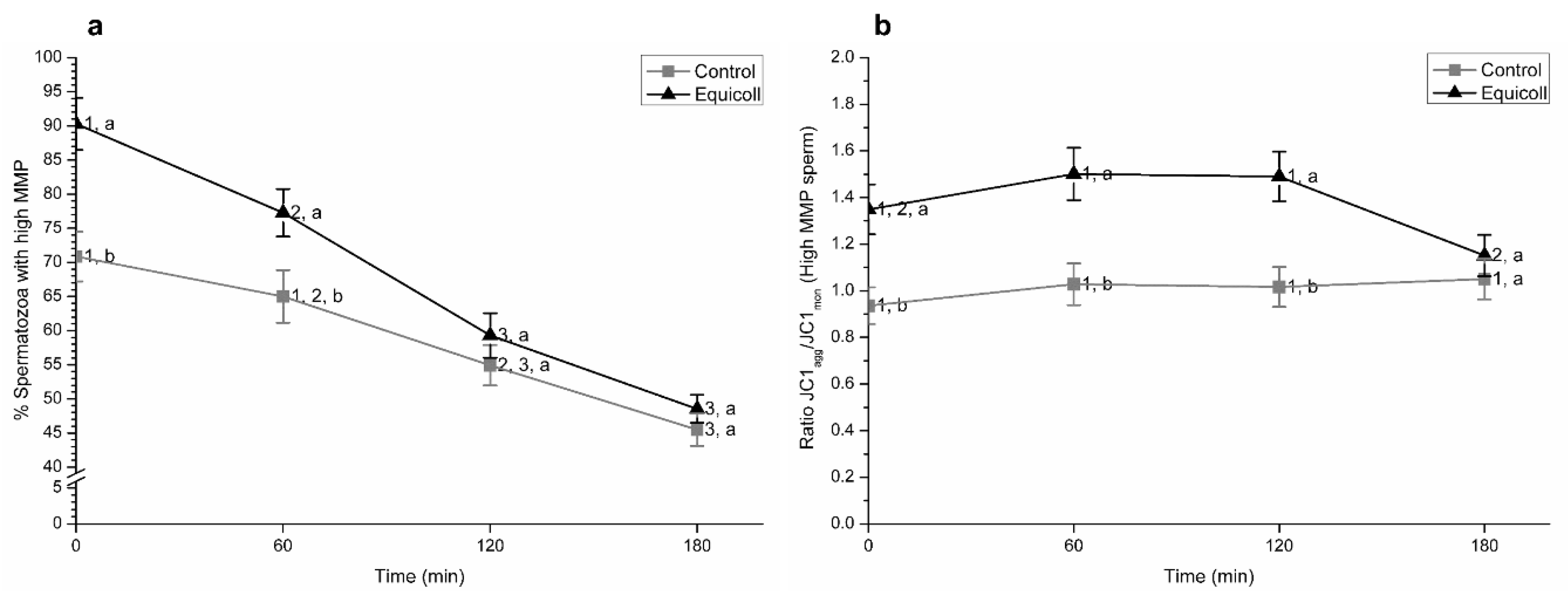
3.2. Mitochondrial Membrane Potential
3.3. Intracellular Calcium Levels
3.4. Intracellular Levels of Peroxides and Superoxides
3.5. Sperm Motility
3.6. Motile Sperm Subpopulations
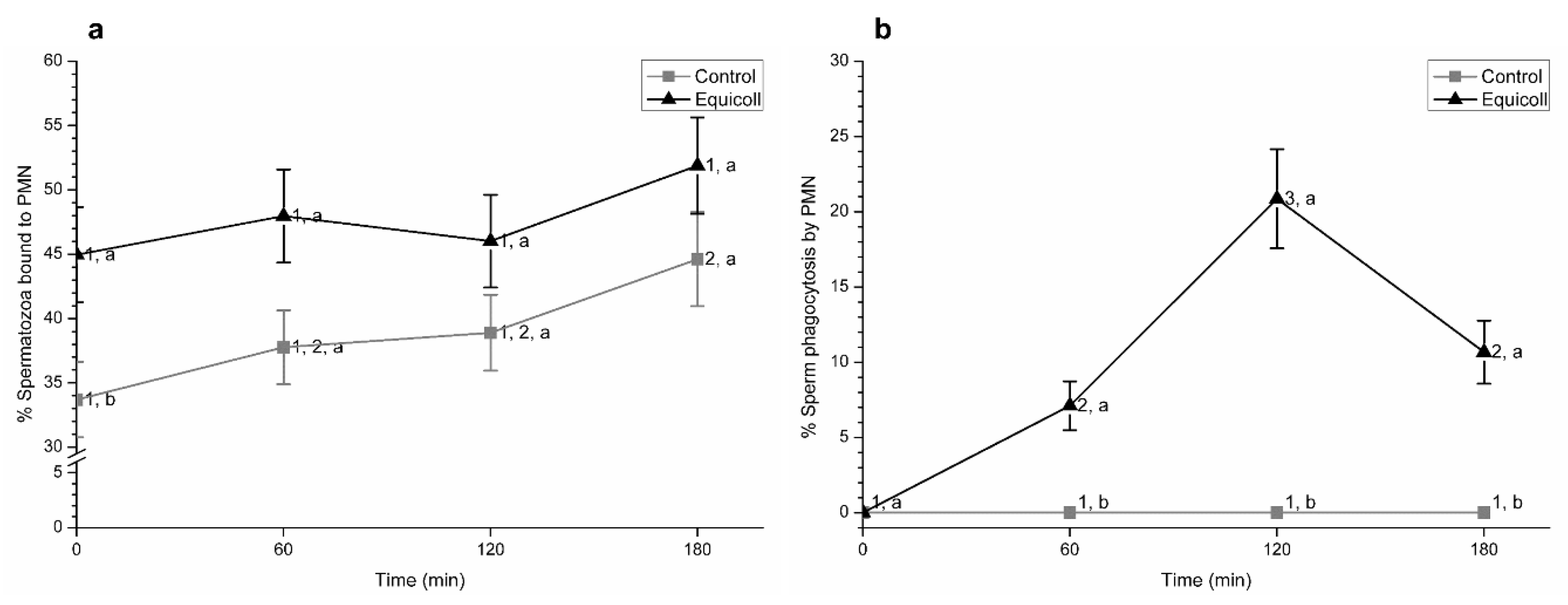
3.7. Sperm-PMN Interaction
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hinrichs, K. Assisted reproductive techniques in mares. Reprod. Domest. Anim. 2018, 53, 4–13. [Google Scholar] [CrossRef]
- Alvarenga, M.A.; Papa, F.O.; Neto, C.R. Advances in Stallion Semen Cryopreservation. Vet. Clin. Equine Pract. 2016, 32, 521–530. [Google Scholar] [CrossRef]
- Morrell, J.M.; Wallgren, M. Colloid centrifugation of boar semen. Reprod. Domest. Anim. 2011, 46, 18–22. [Google Scholar] [CrossRef]
- Oseguera-López, I.; Ruiz-Díaz, S.; Ramos-Ibeas, P.; Pérez-Cerezales, S. Novel Techniques of Sperm Selection for Improving IVF and ICSI Outcomes. Front. Cell Dev. Biol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M. Stallion Sperm Selection: Past, Present, and Future Trends. J. Equine Vet. Sci. 2012, 32, 436–440. [Google Scholar] [CrossRef]
- Roca, J.; Parrilla, I.; Gil, M.A.; Cuello, C.; Martinez, E.A.; Rodriguez-Martinez, H. Non-viable sperm in the ejaculate: Lethal escorts for contemporary viable sperm. Anim. Reprod. Sci. 2016, 169, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Kumaresan, A.; Johannisson, A. Practical implications of sperm selection techniques for improving reproduction. Anim. Reprod. 2017, 14, 572–580. [Google Scholar] [CrossRef]
- Morrell, J.; Alsina, M.S.; Abraham, M.C.; Sjunnesson, Y. Practical applications of sperm selection techniques for improving reproductive efficiency. Anim. Reprod. 2016, 340–345. [Google Scholar] [CrossRef]
- Morrell, J.M.; Rodriguez-Martinez, H. Biomimetic Techniques for Improving Sperm Quality in Animal Breeding: A Review. Open Androl. J. 2009, 1, 1–9. [Google Scholar] [CrossRef]
- Morrell, J.; Dalin, A.M.; Rodriguez-Martinez, H. Comparison of density gradient and single layer centrifugation of stallion spermatozoa: Yield, motility and survival. Equine Vet. J. 2009, 41, 53–58. [Google Scholar] [CrossRef]
- Morrell, J.; Garcia, B.M.; Penã, F.J.; Johannisson, A. Processing stored stallion semen doses by Single Layer Centrifugation. Theriogenology 2011, 76, 1424–1432. [Google Scholar] [CrossRef]
- Ortiz, I.; Dorado, J.; Ramírez, L.; Morrell, J.; Acha, D.; Urbano, M.; Gálvez, M.J.; Carrasco, J.J.; Hidalgo, M. Effect of single layer centrifugation using Androcoll-E-Large on the sperm quality parameters of cooled-stored donkey semen doses. Animal 2014, 308–315. [Google Scholar] [CrossRef]
- Ortiz, I.; Dorado, J.; Morrell, J.M.; Crespo, F.; Gosálvez, J.; Gálvez, M.J.; Acha, D.; Hidalgo, M. Effect of single-layer centrifugation or washing on frozen–thawed donkey semen quality: Do they have the same effect regardless of the quality of the sample? Theriogenology 2015, 84, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Šterbenc, N.; Morrell, J.M.; Kosec, M.; Rath, D.; Klein, S.; Klinc, P. Single layer colloid centrifugation technique improves motility, viability and chromatin integrity of ram spermatozoa after thawing. Cryobiology 2019, 86, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Félez, I.; Castañeda-Sampedro, A.; Sánchez, D.I.; Fernández-Alegre, E.; Álvarez-Rodríguez, M.; Domínguez, J.C.; Morrell, J.M.; Martínez-Pastor, F. Effect of Single Layer Centrifugation Porcicoll (70%, 80% and 90%) or supplementation with reduced glutathione, seminal plasma and bovine serum albumin on frozen-thawed boar sperm. Anim. Reprod. Sci. 2017, 187, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Goodla, L.; Morrell, J.M.; Yusnizar, Y.; Stålhammar, H.; Johannisson, A. Quality of bull spermatozoa after preparation by single-layer centrifugation. J. Dairy Sci. 2014, 97, 2204–2212. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Rodríguez, M.; Álvarez, M.; Anel-López, L.; López-Urueña, E.; Manrique, P.; Borragán, S.; Morrell, J.M.; de Paz, P.; Anel, L. Effect of colloid (Androcoll-Bear, Percoll, and PureSperm) selection on the freezability of brown bear (Ursus arctos) sperm. Theriogenology 2016, 85, 1097–1105. [Google Scholar] [CrossRef]
- Anel-Lopez, L.; Ortega-Ferrusola, C.; Álvarez, M.; Borragán, S.; Chamorro, C.; Peña, F.J.; Morrell, J.; Anel, L.; de Paz, P. Improving sperm banking efficiency in endangered species through the use of a sperm selection method in brown bear (Ursus arctos) thawed sperm. BMC Vet. Res. 2017, 13, 200. [Google Scholar] [CrossRef]
- Martinez-Alborcia, M.J.; Morrell, J.M.; Parrilla, I.; Barranco, I.; Vázquez, J.M.; Martinez, E.A.; Roca, J. Improvement of boar sperm cryosurvival by using single-layer colloid centrifugation prior freezing. Theriogenology 2012, 78, 1117–1125. [Google Scholar] [CrossRef]
- Martinez-Alborcia, M.J.; Morrell, J.M.; Gil, M.A.; Barranco, I.; Maside, C.; Alkmin, D.V.; Parrilla, I.; Martinez, E.A.; Roca, J. Suitability and effectiveness of single layer centrifugation using Androcoll-P in the cryopreservation protocol for boar spermatozoa. Anim. Reprod. Sci. 2013, 140, 173–179. [Google Scholar] [CrossRef]
- Moore, A.I.; Squires, E.L.; Graham, J.K. Effect of seminal plasma on the cryopreservation of equine spermatozoa. Theriogenology 2005, 63, 2372–2381. [Google Scholar] [CrossRef] [PubMed]
- Barrier-Battut, I.; Bonnet, C.; Giraudo, A.; Dubois, C.; Caillaud, M.; Vidament, M. Removal of Seminal Plasma Enhances Membrane Stability on Fresh and Cooled Stallion Spermatozoa. Reprod. Domest. Anim. 2013, 71, 64–71. [Google Scholar] [CrossRef]
- Kruse, R.; Dutta, P.C.; Morrell, J. Colloid centrifugation removes seminal plasma and cholesterol from boar spermatozoa. Reprod. Fertil. Dev. 2011, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Lovaas, B.J.; Bird, S.L.; Lamb, G.C.; Rendahl, A.K.; Taube, P.C.; Foster, D.N. Species-specific interaction of seminal plasma on sperm-neutrophil binding. Anim. Reprod. Sci. 2009, 114, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Miró, J.; Vilés, K.; García, W.; Jordana, J.; Yeste, M. Effect of donkey seminal plasma on sperm movement and sperm-polymorphonuclear neutrophils attachment in vitro. Anim. Reprod. Sci. 2013, 140, 164–172. [Google Scholar] [CrossRef]
- Miró, J.; Marín, H.; Catalán, J.; Papas, M.; Gacem, S.; Yeste, M. Seminal plasma, sperm concentration, and sperm-PMN interaction in the donkey: An in vitro model to study endometrial inflammation at post-insemination. Int. J. Mol. Sci. 2020, 21, 3478. [Google Scholar] [CrossRef]
- Vilés, K.; Rabanal, R.; Rodríguez-Prado, M.; Miró, J. Influence of seminal plasma on leucocyte migration and amount of COX-2 protein in the jenny endometrium after insemination with frozen-thawed semen. Anim. Reprod. Sci. 2013, 143, 57–63. [Google Scholar] [CrossRef]
- Morrell, J.; Van Wienen, M.; Wallgren, M. Single Layer Centrifugation Can Be Scaled-Up Further to Process up to 150 mL Semen. ISRN Vet. Sci. 2011, 2011. [Google Scholar] [CrossRef]
- Petrunkina, A.M.; Waberski, D.; Bollwein, H.; Sieme, H. Identifying non-sperm particles during flow cytometric physiological assessment: A simple approach. Theriogenology 2010, 73, 995–1000. [Google Scholar] [CrossRef]
- Rathi, R.; Colenbrander, B.; Bevers, M.M.; Gadella, B.M. Evaluation of in vitro capacitation of stallion spermatozoa. Biol. Reprod. 2001, 65, 462–470. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Rivera Del Álamo, M.M.; Bonet, S.; Rigau, T.; Rodríguez-Gil, J.E. The increase in phosphorylation levels of serine residues of protein HSP70 during holding time at 17 °C is concomitant with a higher cryotolerance of boar spermatozoa. PLoS ONE 2014, 9, e90887. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ferrusola, C.; Sotillo-Galan, Y.; Varela-Fernandez, E.; Gallardo-Bolanos, J.M.; Muriel, A.; Gonzalez-Fernandez, L.; Tapia, J.A.; Pena, F.J. Detection of ‘Apoptosis-Like’ Changes During the Cryopreservation Process in Equine Sperm. J. Androl. 2007, 29, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Fernández-Novell, J.M.; Ramió-Lluch, L.; Estrada, E.; Rocha, L.G.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Concha, I.I.; Ramírez, A.; Rodríguez-Gil, J.E. Intracellular calcium movements of boar spermatozoa during “in vitro” capacitation and subsequent acrosome exocytosis follow a multiple-storage place, extracellular calcium-dependent model. Andrology 2015, 3, 729–747. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.A.P.; Mairet, B.; Miller, N.G.A. Flow cytometric studies of bicarbonate-mediated Ca2+ influx in boar sperm populations. Mol. Reprod. Dev. 1993, 35, 197–208. [Google Scholar] [CrossRef]
- Kadirvel, G.; Kumar, S.; Kumaresan, A.; Kathiravan, P. Capacitation status of fresh and frozen-thawed buffalo spermatozoa in relation to cholesterol level, membrane fluidity and intracellular calcium. Anim. Reprod. Sci. 2009, 116, 244–253. [Google Scholar] [CrossRef]
- Morrell, J.M.; Winblad, C.; Georgakas, A.; Stuhtmann, G.; Humblot, P.; Johannisson, A. Reactive oxygen species in stallion semen can be affected by season and colloid centrifugation. Anim. Reprod. Sci. 2013, 140, 62–69. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Welch, G.R. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry1. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef]
- Yeste, M.; Bonet, S.; Rodríguez-Gil, J.E.; Rivera Del Álamo, M.M. Evaluation of sperm motility with CASA-Mot: Which factors may influence our measurements? Reprod. Fertil. Dev. 2018, 30, 789–798. [Google Scholar] [CrossRef]
- Luna, C.; Yeste, M.; Rivera Del Alamo, M.M.; Domingo, J.; Casao, A.; Rodriguez-Gil, J.E.; Pérez-Pé, R.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. Effect of seminal plasma proteins on the motile sperm subpopulations in ram ejaculates. Reprod. Fertil. Dev. 2017, 29, 394–405. [Google Scholar] [CrossRef]
- Miró, J.; Lobo, V.; Quintero-Moreno, A.; Medrano, A.; Peña, A.; Rigau, T. Sperm motility patterns and metabolism in Catalonian donkey semen. Theriogenology 2005, 63, 1706–1716. [Google Scholar] [CrossRef]
- Ortiz, I.; Dorado, J.; Acha, D.; Gálvez, M.J.; Urbano, M.A.; Hidalgo, M. Colloid single-layer centrifugation improves post-thaw donkey (Equus asinus) sperm quality and is related to ejaculate freezability. Reprod. Fertil. Dev. 2015, 27, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, T.; Lopes, G.; Pinto, M.; Silva, E.; Miranda, C.; Correia, M.J.; Damásio, L.; Thompson, G.; Rocha, A. Colloid centrifugation of fresh stallion semen before cryopreservation decreased microorganism load of frozen-thawed semen without affecting seminal kinetics. Theriogenology 2015, 83, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Varela, E.; Rey, J.; Plaza, E.; Muñoz de Propios, P.; Ortiz-Rodríguez, J.M.; Álvarez, M.; Anel-López, L.; Anel, L.; De Paz, P.; Gil, M.C.; et al. How does the microbial load affect the quality of equine cool-stored semen? Theriogenology 2018, 114, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Stuhtmann, G.; Meurling, S.; Lundgren, A.; Winblad, C.; Macias Garcia, B.; Johannisson, A. Sperm yield after single layer centrifugation with Androcoll-E is related to the potential fertility of the original ejaculate. Theriogenology 2014, 81, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Richter, J.; Martinsson, G.; Stuhtmann, G.; Hoogewijs, M.; Roels, K.; Dalin, A.M. Pregnancy rates after artificial insemination with cooled stallion spermatozoa either with or without Single Layer Centrifugation. Theriogenology 2014, 82, 1102–1105. [Google Scholar] [CrossRef] [PubMed]
- Yulnawati, Y.; Abraham, M.; Laskowski, D.; Johannisson, A.; Morrell, J. Changes in Bull Sperm Kinematics after Single Layer Centrifugation. Reprod. Domest. Anim. 2014, 49, 954–956. [Google Scholar] [CrossRef]
- Morrell, J.M. Effect of colloid centrifugation on boar sperm quality during storage and function in in vitro fertilization. Theriogenology 2019, 137, 122–126. [Google Scholar] [CrossRef]
- Beydola, T.; Sharma, R.; Agarwal, A. Sperm preparation and selection techniques. In Medical and Surgical Management of Male Infertility, 1st ed.; Rizk, B., Aziz, N., Agarwal, A., Sabanegh, E., Eds.; Jaypee Brothers Medical Publishers: New Delhi, India, 2014; pp. 244–251. [Google Scholar]
- Björkgren, I.; Sipilä, P. The impact of epididymal proteins on sperm function. Reproduction 2019, 158, R155–R167. [Google Scholar] [CrossRef]
- Recuero, S.; Fernandez-Fuertes, B.; Bonet, S.; Barranco, I.; Yeste, M. Potential of seminal plasma to improve the fertility of frozen-thawed boar spermatozoa. Theriogenology 2019, 137, 36–42. [Google Scholar] [CrossRef]
- Troedsson, M.H.T. Breeding-Induced Endometritis in Mares. Vet. Clin. North. Am. Equine Pract. 2006, 22, 705–712. [Google Scholar] [CrossRef]
- Doty, A.; Buhi, W.C.; Benson, S.; Scoggin, K.E.; Pozor, M.; Macpherson, M.; Mutz, M.; Troedsson, M.H.T. Equine CRISP3 modulates interaction between spermatozoa and polymorphonuclear neutrophils. Biol. Reprod. 2011, 85, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Foster, D.N.; Troedsson, M.H.T. Equine seminal plasma reduces sperm binding to polymorphonuclear neutrophils (PMNs) and improves the fertility of fresh semen inseminated into inflamed uteri. Reproduction 2004. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Foster, D.N. Seminal DNase Frees Spermatozoa Entangled in Neutrophil Extracellular Traps. Biol. Reprod. 2005, 73, 1174–1181. [Google Scholar] [CrossRef]
- Alghamdi, A.S.; Funnell, B.J.; Bird, S.L.; Lamb, G.C.; Rendahl, A.K.; Taube, P.C.; Foster, D.N. Comparative studies on bull and stallion seminal DNase activity and interaction with semen extender and spermatozoa. Anim. Reprod. Sci. 2010, 121, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Fichtner, T.; Kotarski, F.; Gärtner, U.; Conejeros, I.; Hermosilla, C.; Wrenzycki, C.; Taubert, A. Bovine sperm samples induce different NET phenotypes in a NADPH oxidase-, PAD4-, and Ca-dependent process. Biol. Reprod. 2020, 102, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Thiam, H.R.; Wong, S.L.; Wagner, D.D.; Waterman, C.M. Cellular Mechanisms of NETosis. Annu. Rev. Cell Dev. Biol. 2020, 36, 191–218. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinematic Parameters | 0 min | 60 min | 120 min | 180 min | ||||
|---|---|---|---|---|---|---|---|---|
| Control | Equicoll | Control | Equicoll | Control | Equicoll | Control | Equicoll | |
| VCL | 155.0 ± 7.8 a | 138.4 ± 9.3 a | 143.4 ± 12.2 a | 102.5 ± 11.2 a | 136.3 ± 14.6 a | 84.8 ± 5.4 b | 108.5 ± 10.7 a | 67.7 ± 4.7 a |
| VSL | 115.3 ± 6.0 a | 96.8 ± 6.9 a | 112.4 ± 5.3 a | 80.6 ± 6.0 b | 109.1 ± 9.0 a | 68.5 ± 4.2 b | 106.9 ± 3.8 a | 53.9 ± 4.2 b |
| VAP | 141.6 ± 5.7 a | 121.9 ± 7.6 a | 130.0 ± 9.5 a | 91.9 ± 9.2 b | 124.7 ± 12.0 a | 76.3 ± 5.2 b | 100.1 ± 9.1 a | 58.9 ± 4.5 b |
| LIN | 74.7 ± 3.4 a | 70.2 ± 2.3 a | 79.6 ± 3.8 a | 79.8 ± 3.1 a | 81.6 ± 4.1 a | 81.0 ± 1.2 a | 84.5 ± 2.3 a | 79.5 ± 1.2 a |
| STR | 81.4 ± 2.6 a | 79.4 ± 2.5 a | 87.2 ± 3.0 a | 88.6 ± 2.5 a | 88.4 ± 3.0 a | 90.1 ± 1.9 a | 91.3 ± 1.6 a | 91.5 ± 0.8 a |
| WOP | 91.6 ± 1.7 a | 88.2 ± 1.0 a | 91.1 ± 1.6 a | 90.0 ± 1.1 a | 92.2 ± 1.6 a | 89.8 ± 0.7 a | 92.4 ± 1.4 a | 86.8 ± 1.2 b |
| ALH | 3.2 ± 0.4 a | 3.1 ± 0.3 a | 3.0 ± 0.4 a | 2.4 ± 0.2 a | 3.0 ± 0.4 a | 2.1 ± 0.1 a | 2.5 ± 0.3 a | 2.1 ± 0.1 a |
| BCF | 9.5 ± 0.8 a | 10.1 ± 1.1 a | 10.0 ± 10.9 a | 9.9 ± 0.6 a | 10.1 ± 1.0 a | 9.1 ± 0.2 a | 8.7 ± 0.1 a | 9.7 ± 0.3 a |
| Kinematic Parameters | SP1 | SP2 | SP3 | SP4 |
|---|---|---|---|---|
| n | 8422 | 8133 | 6152 | 1692 |
| VCL | 165.8 ± 0.2 | 103.8 ± 0.3 | 120.1 ± 0.1 | 0.6 ± 0.1 |
| VSL | 132.8 ± 0.3 | 90.7 ± 0.3 | 46.4 ± 0.3 | 2.0 ± 0.1 |
| VAP | 150.5 ± 0.2 | 97.5 ± 0.3 | 94.4 ± 0.6 | 0.3 ± 0.0 |
| LIN | 80.2 ± 0.1 | 87.3 ± 0.1 | 40.6 ± 0.2 | 0.3 ± 0.1 |
| STR | 88.2 ± 0.1 | 93.2 ± 0.1 | 53.2 ± 0.3 | 0.67 ± 0.1 |
| WOB | 90.6 ± 0.1 | 93.4 ± 0.1 | 767.0 ± 0.1 | 1.0 ± 0.2 |
| ALH | 3.6 ± 0.0 | 2.1 ± 0.0 | 3.6 ± 0.0 | 0.1 ± 0.0 |
| BCF | 11.0 ± 0.0 | 8.4 ± 0.0 | 7.8 ± 0.0 | 0.1 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papas, M.; Catalán, J.; Recuero, S.; Morrell, J.M.; Yeste, M.; Miró, J. Single Layer Centrifugation Improves the Quality of Fresh Donkey Semen and Modifies the Sperm Ability to Interact with Polymorphonuclear Neutrophils. Animals 2020, 10, 2128. https://doi.org/10.3390/ani10112128
Papas M, Catalán J, Recuero S, Morrell JM, Yeste M, Miró J. Single Layer Centrifugation Improves the Quality of Fresh Donkey Semen and Modifies the Sperm Ability to Interact with Polymorphonuclear Neutrophils. Animals. 2020; 10(11):2128. https://doi.org/10.3390/ani10112128
Chicago/Turabian StylePapas, Marion, Jaime Catalán, Sandra Recuero, Jane M. Morrell, Marc Yeste, and Jordi Miró. 2020. "Single Layer Centrifugation Improves the Quality of Fresh Donkey Semen and Modifies the Sperm Ability to Interact with Polymorphonuclear Neutrophils" Animals 10, no. 11: 2128. https://doi.org/10.3390/ani10112128
APA StylePapas, M., Catalán, J., Recuero, S., Morrell, J. M., Yeste, M., & Miró, J. (2020). Single Layer Centrifugation Improves the Quality of Fresh Donkey Semen and Modifies the Sperm Ability to Interact with Polymorphonuclear Neutrophils. Animals, 10(11), 2128. https://doi.org/10.3390/ani10112128