Vultures and Livestock: The Where, When, and Why of Visits to Farms

 ,
,  ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Procedures
2.2. Characteristics of Farms
2.3. Use of Farms by Vultures
2.4. Statistical Analyses
- (1)
- FARM: number of different individual vultures present at each farm during each semester, controlling for the total number of individuals with available information for that period.
- (2)
- VULTURE: number of days each vulture visited each farm per semester, controlling for the total number of days with available information for that vulture during that semester. Since breeding activities may affect the use of particular feeding sources, we performed separate analyses on territorial (i.e., individuals showing territorial behaviour) and non-territorial vultures. For each individual, we quantified home ranges by using 95% kernel density estimates (KernelUD 95%, smoothing factor = 750, see [13] for details) and considered all farms located inside its home range as potential food sources.
2.4.1. Carcass Models
3. Results
3.1. General Patterns
3.2. Model Results
4. Discussion
Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Botha, A.J.; Andevski, J.; Bowden, C.G.; Gudka, M.; Safford, R.J.; Tavares, J.; Williams, N.P. Multi-Species Action Plan to Conserve African-Eurasian Vultures (Vulture MsAP); IUCU: Gland, Switzerland, 2017. [Google Scholar]
- Arrondo, E.; Cortés-Avizanda, A.; Donázar, J.A. Temporally unpredictable supplementary feeding may benefit endangered scavengers. Ibis 2015, 157, 648–651. [Google Scholar] [CrossRef]
- Cortés-Avizanda, A.; Jovani, R.; Carrete, M.; Donázar, J.A. Resource unpredictability promotes species diversity and coexistence in an avian scavenger guild: A field experiment. Ecology 2012, 93, 2570–2579. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, J.; Benhamou, S.; Riotte-Lambert, L.; Duriez, O. Assessing the risk for an obligate scavenger to be dependent on predictable feeding sources. Biol. Conserv. 2017, 215, 92–98. [Google Scholar] [CrossRef]
- Moleón, M.; Sánchez-Zapata, J.A.; Margalida, A.; Carrete, M.; Owen-Smith, N.; Donázar, J.A. Humans and scavengers: The evolution of interactions and ecosystem services. BioScience 2014, 64, 394–403. [Google Scholar] [CrossRef]
- Morales-Reyes, Z.; Pérez-García, J.M.; Moleón, M.; Botella, F.; Carrete, M.; Lazcano, C.; Moreno-Opo, R.; Margalida, A.; Donázar, J.A.; Sánchez-Zapata, J.A. Supplanting ecosystem services provided by scavengers raises greenhouse gas emissions. Sci. Rep. 2015, 5, 7811. [Google Scholar] [CrossRef]
- Donázar, J.A.; Margalida, A.; Campión, D. Vultures, Feeding Stations and Sanitary Legislation: A Conflict and Its Consequences from the Perspective of Conservation Biology; Munibe 29 (Suppl); Society of Sciences Aranzadi: San Sebastián, Spain, 2009; ISBN 9788493598662. [Google Scholar]
- Margalida, A.; Carrete, M.; Sánchez-Zapata, J.A.; Donázar, J.A. Good News for European Vultures. Science 2012, 335, 284. [Google Scholar] [CrossRef][Green Version]
- Arrondo, E.; Moleón, M.; Cortés-Avizanda, A.; Jiménez, J.; Beja, P.; Sánchez-Zapata, J.A.; Donázar, J.A. Invisible barriers: Differential sanitary regulations constrain vulture movements across country borders. Biol. Conserv. 2018, 219, 46–52. [Google Scholar] [CrossRef]
- Cortés-Avizanda, A.; Carrete, M.; Donázar, J.A. Managing supplementary feeding for avian scavengers: Guidelines for optimal design using ecological criteria. Biol. Conserv. 2010, 143, 1707–1715. [Google Scholar] [CrossRef]
- Moreno-Opo, R.; Trujillano, A.; Arredondo, Á.; González, L.M.; Margalida, A. Manipulating size, amount and appearance of food inputs to optimize supplementary feeding programs for European vultures. Biol. Conserv. 2015, 181, 27–35. [Google Scholar] [CrossRef]
- Cortés-Avizanda, A.; Blanco, G.; DeVault, T.L.; Markandya, A.; Virani, M.Z.; Brandt, J.; Donázar, J.A. Supplementary feeding and endangered avian scavengers: Benefits, caveats, and controversies. Front. Ecol. Environ. 2016, 14, 191–199. [Google Scholar] [CrossRef]
- van Overveld, T.; García-Alfonso, M.; Dingemanse, N.J.; Bouten, W.; Gangoso, L.; de la Riva, M.; Serrano, D.; Donázar, J.A. Food predictability and social status drive individual resource specializations in a territorial vulture. Sci. Rep. 2018, 8, 15155. [Google Scholar] [CrossRef] [PubMed]
- Medina, F.M. Alimentación del alimoche, Neophron percnopterus (L.), en Fuerteventura, Islas Canarias (Aves, Accipitridae). Vieraea 1999, 27, 77–86. [Google Scholar]
- Gangoso, L.; Donázar, J.A.; Scholz, S.; Palacios, C.J.; Hiraldo, F. Contradiction in conservation of island ecosystems: Plants, introduced herbivores and avian scavengers in the Canary Islands. Biodivers. Conserv. 2006, 15, 2231–2248. [Google Scholar] [CrossRef]
- Blanco, G.; Cortés-Avizanda, A.; Frías, Ó.; Arrondo, E.; Donázar, J.A. Livestock farming practices modulate vulture diet-disease interactions. Glob. Ecol. Conserv. 2019, 17. [Google Scholar] [CrossRef]
- García-Alfonso, M.; Morales-Reyes, Z.; Gangoso, L.; Bouten, W.; Sánchez-Zapata, J.A.; Serrano, D.; Donázar, J.A. Probing into farmers’ perceptions of a globally endangered ecosystem service provider. Ambio 2018, 48, 900–912. [Google Scholar] [CrossRef] [PubMed]
- García-Heras, M.-S.; Cortés-Avizanda, A.; Donázar, J.A. Who are we feeding? Asymmetric individual use of surplus food resources in an insular population of the endangered Egyptian Vulture Neophron percnopterus. PLoS ONE 2013, 8, e80523. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Aguilar, A.; Jovani, R.; Melián, C.J.; Pradel, R.; Tella, J.L. Multi-event capture-recapture analysis reveals individual foraging specialization in a generalist species. Ecology 2015, 96, 1650–1660. [Google Scholar] [CrossRef]
- Krüger, S.; Reid, T.; Amar, A. Differential Range Use between Age Classes of Southern African Bearded Vultures Gypaetus barbatus. PLoS ONE 2014, 9, e114920. [Google Scholar] [CrossRef]
- Wolfson, D.W.; Fieberg, J.R.; Andersen, D.E. Juvenile Sandhill Cranes exhibit wider ranging and more exploratory movements than adults during the breeding season. Ibis 2020, 162, 556–562. [Google Scholar] [CrossRef]
- Gavashelishvili, A.; McGrady, M.J. Geographic information system-based modelling of vulture response to carcass appearance in the Caucasus. J. Zool. 2006, 269, 365–372. [Google Scholar] [CrossRef]
- Monsarrat, S.; Benhamou, S.; Sarrazin, F.; Bessa-Gomes, C.; Bouten, W.; Duriez, O. How Predictability of Feeding Patches Affects Home Range and Foraging Habitat Selection in Avian Social Scavengers? PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Delgado, O.; García Gallo, A.; Reyes Betancort, J.A. Estudio fitosociológico de la vegetación actual de Fuerteventura (Islas Canarias). Vieraea 2000, 28, 61–98. [Google Scholar]
- García-Martínez, A.; Olaizola, A.; Bernués, A. Trajectories of evolution and drivers of change in European mountain cattle farming systems. Animal 2009, 3, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Canarian Government Estadistica Ganadera. Available online: http://www.gobiernodecanarias.org/istac/ (accessed on 26 August 2018).
- Donázar, J.A.; Palacios, C.J.; Gangoso, L.; Ceballos, O.; Gonzáles, M.J.; Hiraldo, F. Conservation status and limiting factors in the endangered population of Egyptian Vulture (Neophron percnopterus) in the Canary Islands. Biol. Conserv. 2002, 107, 89–97. [Google Scholar] [CrossRef]
- Agudo, R.; Carrete, M.; Alcaide, M.; Rico, C.; Hiraldo, F.; Donázar, J.A. Genetic diversity at neutral and adaptive loci determines individual fitness in a long-lived territorial bird. Proc. Biol. Sci. 2012, 279, 3241–3249. [Google Scholar] [CrossRef]
- Donázar, J.A.; Negro, J.J.; Palacios, C.J.; Gangoso, L.; Godoy, J.A.; Ceballos, O.; Hiraldo, F.; Capote, N. Description of a new subspecies of the Egyptian Vulture (Accipitridae: Neophron percnopterus) from the Canary Islands. J. Raptor Res. 2002, 36, 17–23. [Google Scholar]
- Donázar, J.A.; Barbosa, J.M.; García-Alfonso, M.; van Overveld, T.; Gangoso, L.; de la Riva, M. Too much is bad: Increasing numbers of livestock and conspecifics reduce body mass in an avian scavenger. Ecol. Appl. 2020, e02125. [Google Scholar] [CrossRef]
- Badia-Boher, J.A.; Sanz-Aguilar, A.; de la Riva, M.; Gangoso, L.; van Overveld, T.; García-Alfonso, M.; Luzardo, O.P.; Suarez-Pérez, A.; Donázar, J.A. Evaluating European LIFE conservation projects: Improvements in survival of an endangered vulture. J. Appl. Ecol. 2019, 56, 1210–1219. [Google Scholar] [CrossRef]
- Bouten, W.; Baaij, E.W.; Shamoun-Baranes, J.; Camphuysen, K.C.J. A flexible GPS tracking system for studying bird behaviour at multiple scales. J. Ornithol. 2013, 154, 571–580. [Google Scholar] [CrossRef]
- Sergio, F.; Tavecchia, G.; Tanferna, A.; López Jiménez, L.; Blas, J.; De Stephanis, R.; Marchant, T.A.; Kumar, N.; Hiraldo, F. No effect of satellite tagging on survival, recruitment, longevity, productivity and social dominance of a raptor, and the provisioning and condition of its offspring. J. Appl. Ecol. 2015, 52, 1665–1675. [Google Scholar] [CrossRef]
- MAGRAMA Spanish Ministry of Agriculture, Food and Environment. Available online: http://www.magrama.gog.es (accessed on 19 June 2015).
- Centro Nacional de Información Geofráfica [CNIG] Modelo Digital de Elevaciones 25. Available online: http://centrodedescargas.cnig.es/CentroDescargas/catalogo.do?Serie=MAPLI (accessed on 16 November 2016).
- Klaassen, R.H.G.; Schlaich, A.E.; Bouten, W.; Koks, B.J. Migrating Montagu’s harriers frequently interrupt daily flights in both Europe and Africa. J. Avian Biol. 2017, 48, 180–190. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Savelieve, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Gails, M., Krickeberg, K., Samet, J.M., Tsiatis, A., Wong, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; p. 574. ISBN 978-0-387-87457-9. [Google Scholar]
- Sugiura, N. Further analysis of the data by akaike’ s information criterion and the finite corrections. Commun. Stat. Theory Methods 1978, 7, 13–26. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Bolker, B.M. GLMM FAQ. Available online: https://bbolker.github.io/mixedmodels-misc/glmmFAQ.html (accessed on 15 April 2020).
- Gelman, A.; Hill, J. Linear regression: Before and after fitting the model. In Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2007; Chapter 4; pp. 53–77. ISBN 9780521867061. [Google Scholar]
- Graham, M.H. Statistical confronting multicollinearity in ecological. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Arnold, T.W. Uninformative parameters and model selection using Akaike’s information criterion. J. Wildl. Manag. 2010, 74, 1175–1178. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2002; Volume 172, p. 488. ISBN 978-0-387-22456-5. [Google Scholar]
- Crawley, M.J. Statistical Computing: An Introduction to Data Analysis Using S-Plus; Wiley: New York, NY, USA, 2002. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Bjørnstad, O.N.; Falck, W. Nonparametric spatial covariance functions: Estimation and testing. Environ. Ecol. Stat. 2001, 8, 53–70. [Google Scholar] [CrossRef]
- Moreira, F.; Martins, R.C.; Catry, I.; D’Amico, M. Drivers of power line use by white storks: A case study of birds nesting on anthropogenic structures. J. Appl. Ecol. 2018. [Google Scholar] [CrossRef]
- R Core Team R. A Language and Environment For Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package, version 2.0-4. 2016. Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 15 November 2020).
- Barton, K. MuMIn: Multi-Model Inference. R Package, version 1.15.6. 2016; Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 November 2020).
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Brown, J.A.; Jimenez-Valverde, A.; Real, R. modEvA: Model Evaluation and Analysis. R Package, version 1.3.2. 2016; Available online: https://CRAN.R-project.org/package=modEvA (accessed on 15 November 2020).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package, version 0.9-69-3. 2018. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 15 November 2020).
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Modell. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Nakagawa, S.; Johnson, P.C.D.; Schielzeth, H. The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J. R. Soc. Interface 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Barton, K. MuMIn: Multi-Model Inference. R Package, version 1.43.6. 2019; Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 November 2020).
- Harrison, X.A. A comparison of observation-level randomeffect and Beta-Binomial models for modelling overdispersion in Binomial data in ecology & evolution. PeerJ 2015, 2015. [Google Scholar] [CrossRef]
- Morant Etxebarria, J.; López-López, P.; Zuberogoitia Arroyo, I. Parental investment asymmetries of a globally endangered scavenger: Unravelling the role of gender, weather conditions and stage of the nesting cycle. Bird Study 2019, 66, 329–341. [Google Scholar] [CrossRef]
- Newton, I. Population Ecology of Raptors; A&C Black: London, UK, 1979; ISBN 978-0856610233. [Google Scholar]
- Shaffer, S.A.; Weimerskirch, H.; Costa, D.P. Functional significance of sexual dimorphism in Wandering Albatrosses, Diomedea exulans. Funct. Ecol. 2001, 15, 203–210. [Google Scholar] [CrossRef]
- Ceballos, O.; Donázar, J.A. Actividad, uso del espacio y cuidado parental en una pareja de alimoches (Neophron percnopterus) durante el periodo de dependencia de los pollos. Ecología 1988, 2, 275–291. [Google Scholar]
- Van Overveld, T.; Gangoso, L.; García-Alfonso, M.; Bouten, W.; de la Riva, M.; Donázar, J.A. Seasonal grouping dynamics in a territorial vulture: Ecological drivers and social consequences. Behav. Ecol. Sociobiol. 2020, 74. [Google Scholar] [CrossRef]
- Donázar, J.A.; Ceballos, O.; Cortés-Avizanda, A. Tourism in protected areas: Disentangling road and traffic effects on intra-guild scavenging processes. Sci. Total Environ. 2018, 630, 600–608. [Google Scholar] [CrossRef]
- Bautista, L.M.; García, J.T.; Calmaestra, R.G.; Palacín, C.; Martín, C.A.; Morales, M.B.; Bonal, R.; Viñuela, J. Effect of weekend road traffic on the use of space by raptors. Conserv. Biol. 2004, 18, 726–732. [Google Scholar] [CrossRef]
- Tejera, G.; Rodríguez, B.; Armas, C.; Rodríguez, A. Wildlife-vehicle collisions in Lanzarote biosphere reserve, Canary Islands. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
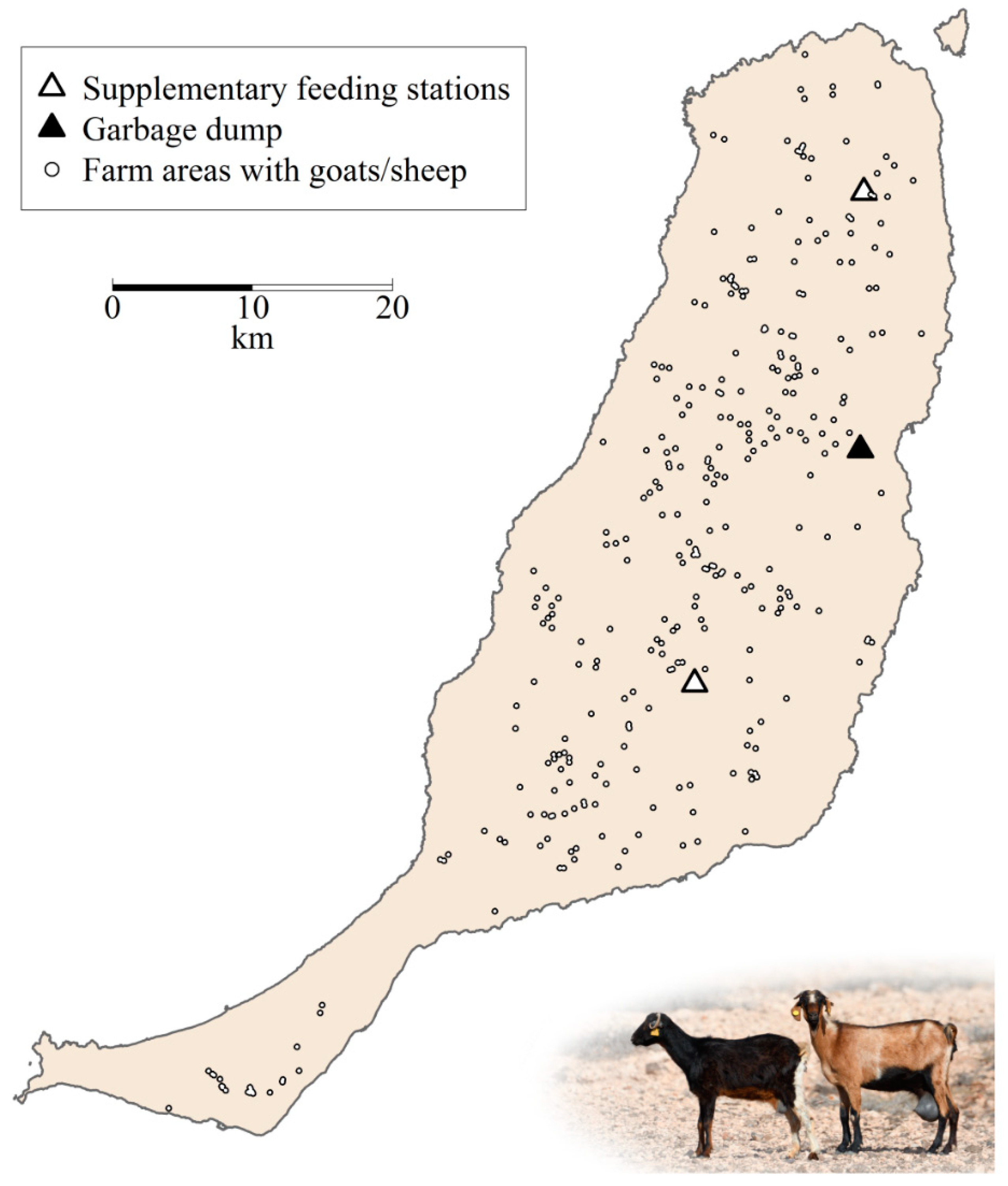{kind=link}
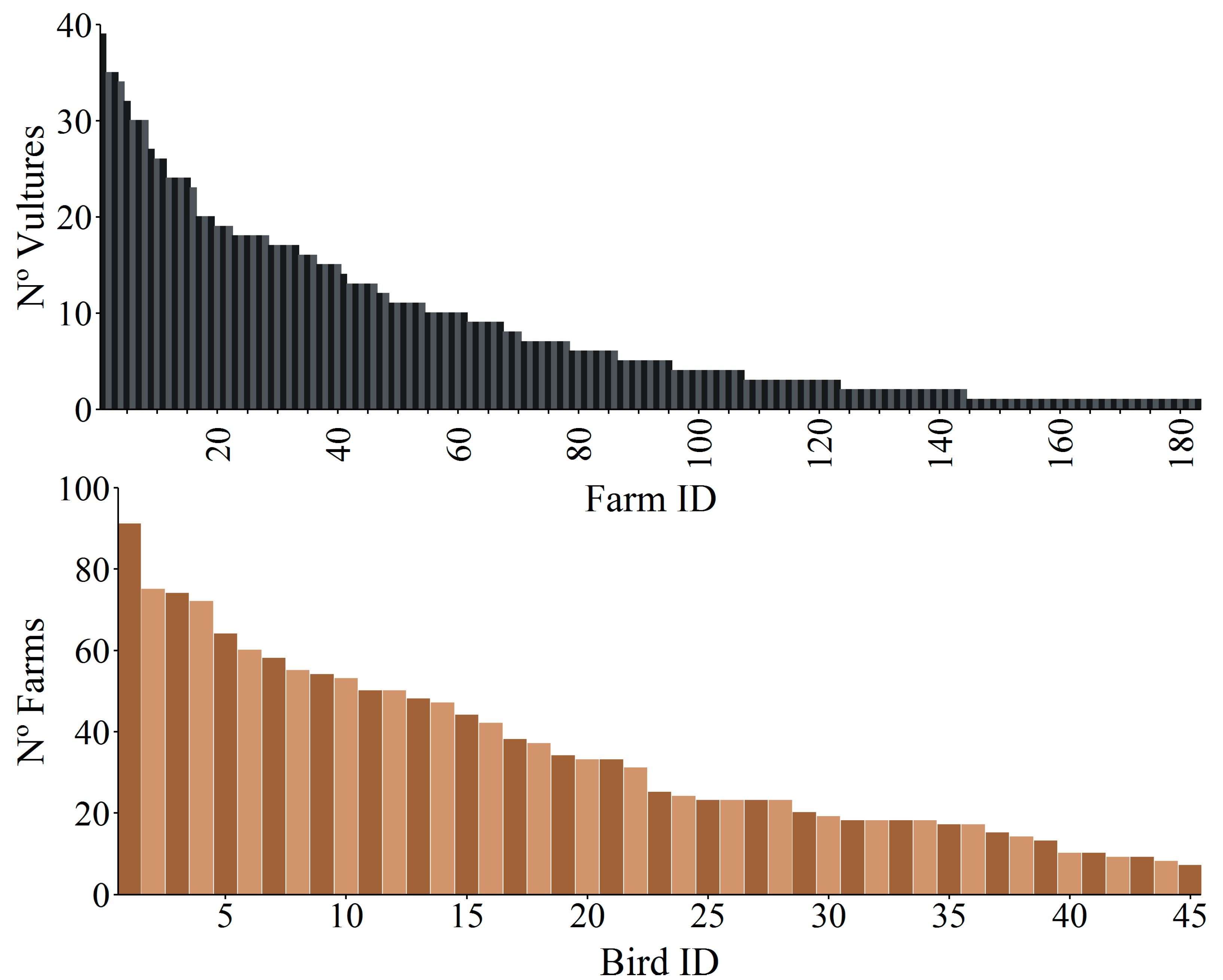{kind=link}
{kind=link}
| Drivers | Variable | Predictions | References | |
|---|---|---|---|---|
| Territorial | Non-Territorial | |||
| Individual characteristics | Sex | Males will forage more than females on farms, because females heavily rely on supplementary feeding stations. | [13,18] | |
| Age | The probability of visiting a farm will be affected by individuals’ age due to differences in foraging strategies, skills, environmental knowledge, and movement patterns. | [13,19,20,21] | ||
| Success | The probability of visiting a farm increases with successful breeding because of increased food demands. | - | ||
| AreaK95 | The probability of visiting a farm increases with smaller home ranges, as the birds would concentrate searching effort in fewer and better-known areas. | |||
| Main food resources provided by farms | Goats Sheep | Farms with more resources (number of heads of livestock) would receive more visits. | ||
| Carcass | Farms with more probability of abandoning livestock carcasses would receive more visits. | |||
| Temporal constraints | Breeding | Farms will receive fewer visits during the breeding season due to spatial constraints derived from territory attendance (territorial birds) and prospecting behavior (non-territorial birds). | [18] | |
| Spatial constraints relative to farms | Dist Terr | Visits increase with distance to the nearest occupied territory because owners defend trophic resources in areas close to their nests. | [18] | |
| Dist Road | Visits increase with distance to roads and urban areas because of less human disturbance. | [22] | ||
| Dist Urb | ||||
| Dist Nest | Visits decrease with distance to the nest because of central-place foraging restraints. | - | [4] | |
| Dist K50 | Visits decrease with distance to core areas because of central-place foraging restraints. | [18] | ||
| Dist HPFP | Visits decrease with distance to highly predictable feeding places (HPFP) because these places offer high amounts of food resources also acting as social meeting points. Therefore, individuals concentrate their activity around these points. | [13,23] | ||
| Variable | Description |
|---|---|
| Fixed Factors | |
| Dist HPFP a,b,c | For each farm, distance from the centroid to the nearest HPFP (highly predictable feeding place): dump or supplementary feeding stations. |
| Dist Urb a,b,c | For each farm, distance from the centroid to the nearest urban area. |
| Dist Road a,b,c | For each farm, distance from the centroid to the nearest road. |
| Dist Terr b,c | For each farm, individual, and semester, distance from the centroid of the farm to the nearest nest (different from their own nest in the case of territorial). |
| Dist K50 b,c | For each farm, individual, and semester, distance from the centroid of the farm to the nearest core area defined by the KernelUD 50%. |
| Goat Sheep a,b,c | For each farm and semester, the number of goats and sheep on the basis of livestock censuses (see Methods). |
| Carcass a,b,c | For each farm, answer to the question about whether they abandon or not carcasses (affirmative answer = 1, negative = 0). When no information was available, it was based on the probability resulting from a predictive model performed (see Methods 2.4.1). |
| AreaK95 a,b,c | For each individual and semester, the size of the area defined by KernelUD 95% (km2). |
| Breeding a,b,c | For each semester, breeding season (1) or not (0). |
| Sex b,c | For each individual, male (M) or female (F). |
| Age b,c | For each individual and semester, age measured in years, calculated as the year of the semester minus year of birth. |
| Dist Nest b | For each farm, breeding individual, and semester, distance from the centroid of the farm to the nest. |
| Success b | For each individual and year, having at least one fledgling (1) or none (0) during the breeding season of the year. |
| Random Factors | |
| Semester ID a,b,c | Period of six months defined as 1st semester or 2nd semester for each year from 2nd semester of 2013 to 2nd semester of 2016. |
| Farm ID a,b,c | Farm identity, defined by one or more farms and the area around them with a threshold distance of 180 meters. |
| Bird ID b,c | Individual identifier of each vulture. |
| N.farms HR | |||||
| Territorial | Non-territ. | ||||
| Breeding | Non-Breed. | Breeding | Non-Breed. | ||
| Differences between sexes | W | 123.5 | 170 | 57 | 210 |
| p | 0.665 | 1.000 | <0.01 | <0.01 | |
| N.farms Visited | |||||
| Territorial | Non-territ. | ||||
| Breeding | Non-Breed. | Breeding | Non-Breed. | ||
| Differences between sexes | W | 111 | 136.5 | 104 | 217 |
| p | 0.374 | 0.312 | 0.004 | <0.001 | |
| Male | Female | Male | Female | ||
| Differences between seasons | W | 152 | 185 | 203 | 95.5 |
| p | 0.574 | 0.657 | <0.001 | <0.001 | |
| Variable | Estimate | Std. Error | 7.5% | 92.5% | RI |
|---|---|---|---|---|---|
| FARM | |||||
| (Intercept) | −4.41 | 0.19 | −4.68 | −4.13 | |
| Goat Sheep | 0.44 | 0.07 | 0.35 | 0.54 | |
| Carcass | 0.56 | 0.14 | 0.36 | 0.75 | |
| Dist Road | 0.60 | 0.13 | 0.41 | 0.79 | |
| Dist HPFP | −0.86 | 0.14 | −1.06 | −0.66 | |
| Breeding | 0.49 | 0.21 | 0.19 | 0.79 | |
| Breeding:Dist HPFP | 0.31 | 0.07 | 0.22 | 0.40 | |
| Territorial VULTURES | |||||
| (Intercept) | −7.06 | 0.34 | −7.54 | −6.57 | |
| Sex | 0.43 | 0.35 | 0.24 | 0.98 | 0.70 |
| AreaK95 | −0.45 | 0.04 | −0.51 | −0.39 | 1.00 |
| Breeding | 0.04 | 0.13 | 0.03 | 0.56 | 0.15 |
| Dist Terr | 0.80 | 0.06 | 0.72 | 0.88 | 1.00 |
| Dist HPFP | 0.27 | 0.19 | 0.12 | 0.56 | 0.80 |
| Dist Road | 0.45 | 0.25 | 0.23 | 0.80 | 0.87 |
| Goat Sheep | 0.35 | 0.09 | 0.22 | 0.48 | 1.00 |
| Carcass | 0.01 | 0.06 | 0.06 | 0.70 | 0.02 |
| Dist K50 | −1.33 | 0.05 | −1.39 | −1.26 | 1.00 |
| Age | −0.12 | 0.15 | −0.33 | 0.09 | 1.00 |
| Age:Dist K50 | −0.79 | 0.05 | −0.86 | −0.71 | 1.00 |
| Non-territorial VULTURES | |||||
| (Intercept) | −7.13 | 0.21 | −7.43 | −6.82 | |
| Age | 0.03 | 0.04 | 0.00 | 0.12 | 0.51 |
| AreaK95 | −0.57 | 0.03 | −0.61 | −0.53 | 1.00 |
| Dist Road | 1.24 | 0.15 | 1.02 | 1.45 | 1.00 |
| Dist K50 | −0.68 | 0.02 | −0.71 | −0.65 | 1.00 |
| Goat Sheep | 0.66 | 0.04 | 0.61 | 0.71 | 1.00 |
| Carcass | 0.17 | 0.19 | 0.04 | 0.53 | 0.62 |
| Dist Terr | 0.21 | 0.05 | 0.14 | 0.28 | 1.00 |
| Breeding | 0.14 | 0.08 | 0.02 | 0.25 | 1.00 |
| Breeding:Dist Terr | 0.28 | 0.02 | 0.25 | 0.31 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Alfonso, M.; van Overveld, T.; Gangoso, L.; Serrano, D.; Donázar, J.A. Vultures and Livestock: The Where, When, and Why of Visits to Farms. Animals 2020, 10, 2127. https://doi.org/10.3390/ani10112127
García-Alfonso M, van Overveld T, Gangoso L, Serrano D, Donázar JA. Vultures and Livestock: The Where, When, and Why of Visits to Farms. Animals. 2020; 10(11):2127. https://doi.org/10.3390/ani10112127
Chicago/Turabian StyleGarcía-Alfonso, Marina, Thijs van Overveld, Laura Gangoso, David Serrano, and José A. Donázar. 2020. "Vultures and Livestock: The Where, When, and Why of Visits to Farms" Animals 10, no. 11: 2127. https://doi.org/10.3390/ani10112127
APA StyleGarcía-Alfonso, M., van Overveld, T., Gangoso, L., Serrano, D., & Donázar, J. A. (2020). Vultures and Livestock: The Where, When, and Why of Visits to Farms. Animals, 10(11), 2127. https://doi.org/10.3390/ani10112127