Optimization of CASA-Mot Analysis of Donkey Sperm: Optimum Frame Rate and Values of Kinematic Variables for Different Counting Chamber and Fields




Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals Used and Ethics Statement
2.2. Semen Preparation
2.3. Semen Dilution
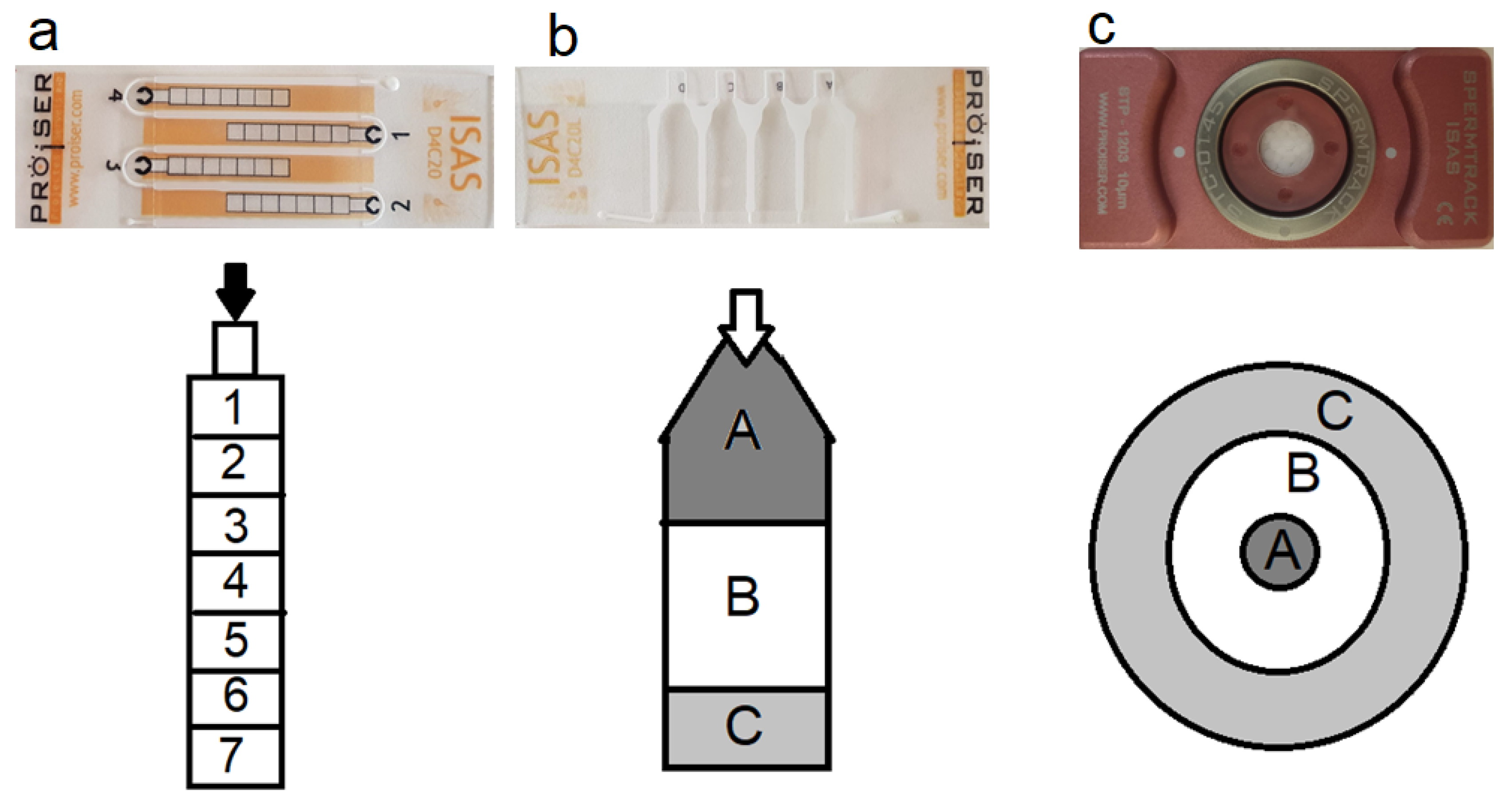
2.4. Counting Chambers and Loading Technique
2.5. CASA-Mot Analysis
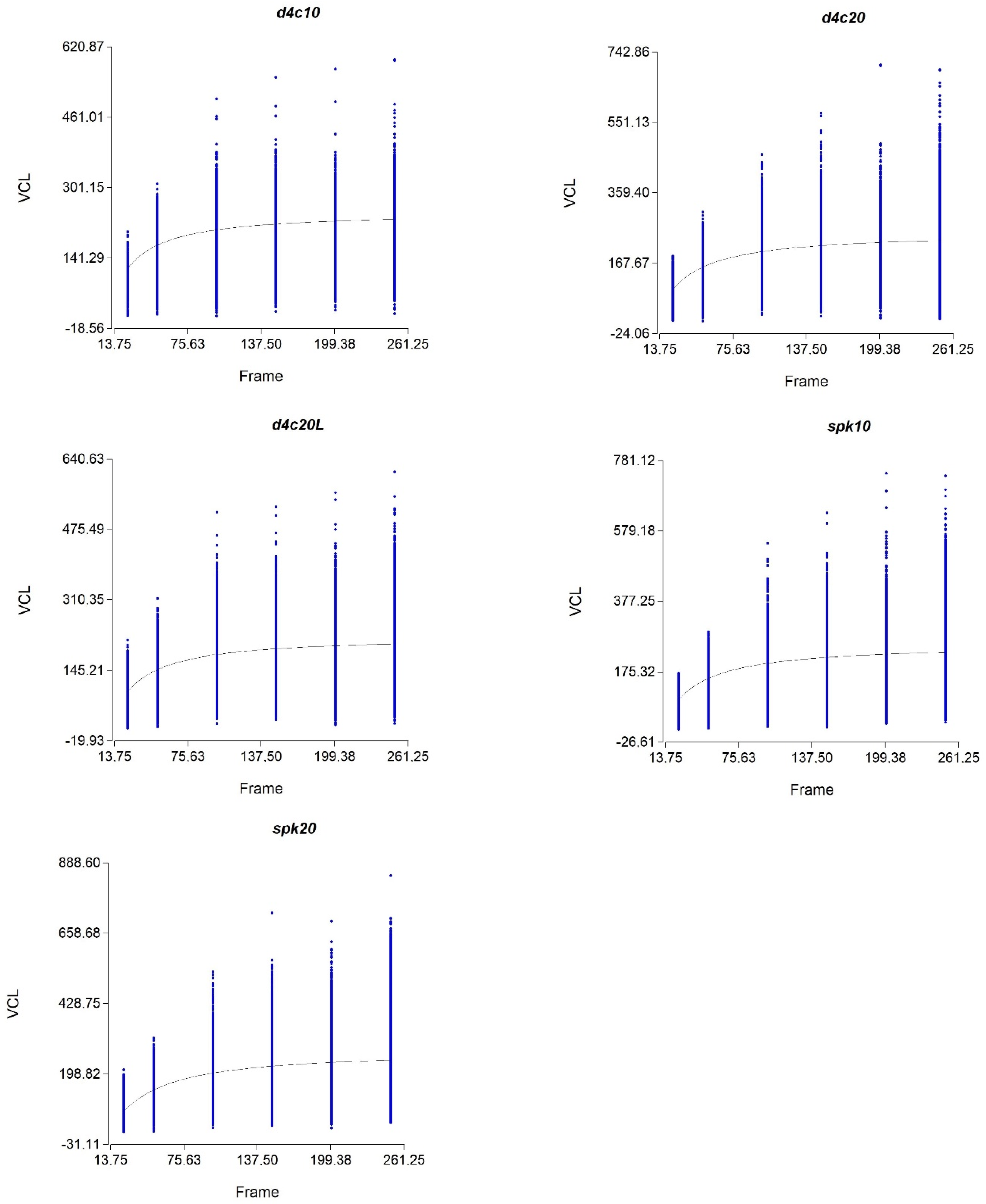
2.6. Calculating the Optimum Frame Rate
2.7. Statistical Analysis
3. Results
3.1. Optimum Frame Rate in Different Chambers
3.2. Effect of Chamber Type and Depth on Sperm Kinematic Parameters
3.3. Effect of the Capture Field Inside the Counting Chamber
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carneiro, G.F.; Lucena, J.E.C.; Barros, L.D.O. The Current Situation and Trend of the Donkey Industry in South America. J. Equine Vet. Sci. 2018, 65, 106–110. [Google Scholar] [CrossRef]
- Faccia, M.; D’Alessandro, A.G.; Summer, A.; Hailu, Y. Milk products from minor dairy species: A review. Animals 2020, 10, 1260. [Google Scholar] [CrossRef] [PubMed]
- Camillo, F.; Rota, A.; Biagini, L.; Tesi, M.; Fanelli, D.; Panzani, D. The Current Situation and Trend of Donkey Industry in Europe. J. Equine Vet. Sci. 2018, 65, 44–49. [Google Scholar] [CrossRef]
- Yılmaz, O.; Boztepe, S.; Ertuğrul, M. The domesticated donkey: III-economic importance, uncommon usages, reproduction traits, genetics, nutrition and health care. Can. J. Appl. Sci. 2012, 3, 320–338. [Google Scholar]
- Varner, D.D.D. Developments in stallion semen evaluation. Theriogenology 2008, 70, 448–462. [Google Scholar] [CrossRef]
- Giaretta, E.; Munerato, M.; Yeste, M.; Galeati, G.; Spinaci, M.; Tamanini, C.; Mari, G.; Bucci, D. Implementing an open-access CASA software for the assessment of stallion sperm motility: Relationship with other sperm quality parameters. Anim. Reprod. Sci. 2017, 176, 11–19. [Google Scholar] [CrossRef]
- Colenbrander, B.; Gadella, B.; Stout, T. The Predictive Value of Semen Analysis in the Evaluation of Stallion Fertility. Reprod. Domest. Anim. 2003, 38, 305–311. [Google Scholar] [CrossRef]
- McCue, P.M. Breeding Soundness Evaluation of the Stallion. In Equine Reproductive Procedures; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2014; pp. 319–324. [Google Scholar]
- Whitesell, K.; Stefanovski, D.; McDonnell, S.; Turner, R. Evaluation of the effect of laboratory methods on semen analysis and breeding soundness examination (BSE) classification in stallions. Theriogenology 2020, 142, 67–76. [Google Scholar] [CrossRef]
- Broekhuijse, M.L.W.J.; Šostarić, E.; Feitsma, H.; Gadella, B.M. Additional value of computer assisted semen analysis (CASA) compared to conventional motility assessments in pig artificial insemination. Theriogenology 2011, 76, 1473–1486. [Google Scholar] [CrossRef]
- Holt, W.V.; Cummins, J.M.; Soler, C. Computer-assisted sperm analysis and reproductive science; a gift for understanding gamete biology from multidisciplinary perspectives. Reprod. Fertil. Dev. 2018, 30, 3–5. [Google Scholar] [CrossRef]
- Yániz, J.L.; Silvestre, M.A.; Santolaria, P.; Soler, C. CASA-Mot in mammals: An update. Reprod. Fertil. Dev. 2018, 30, 799–809. [Google Scholar] [CrossRef]
- Canisso, I.F.; Panzani, D.; Miró, J.; Ellerbrock, R.E. Key Aspects of Donkey and Mule Reproduction. Vet. Clin. N. Am. Equine Pract. 2019, 35, 607–642. [Google Scholar] [CrossRef]
- Bompart, D.; García-Molina, A.; Valverde, A.; Caldeira, C.; Yániz, J.; De Murga, M.N.; Soler, C. CASA-Mot technology: How results are affected by the frame rate and counting chamber. Reprod. Fertil. Dev. 2018, 30, 810–819. [Google Scholar] [CrossRef]
- Contri, A.; Gloria, A.; Robbe, D.; De Amicis, I.; Carluccio, A. Characteristics of donkey spermatozoa along the length of the epididymis. Theriogenology 2012, 77, 166–173. [Google Scholar] [CrossRef]
- Quartuccio, M.; Marino, G.; Zanghì, A.; Garufi, G.; Cristarella, S. Testicular Volume and Daily Sperm Output in Ragusano Donkeys. J. Equine Vet. Sci. 2011, 31, 143–146. [Google Scholar] [CrossRef]
- Carluccio, A.; Panzani, S.; Contri, A.; Bronzo, V.; Robbe, D.; Veronesi, M.C. Influence of season on testicular morphometry and semen characteristics in Martina Franca jackasses. Theriogenology 2013, 79, 502–507. [Google Scholar] [CrossRef]
- Gacem, S.; Papas, M.; Catalan, J.; Miró, J. Examination of jackass (Equus asinus) accessory sex glands by B-mode ultrasound and of testicular artery blood flow by colour pulsed-wave Doppler ultrasound: Correlations with semen production. Reprod. Domest. Anim. 2020, 55, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.N.K.; Sayed, R.; Zayed, A.E.; Abdel-Hafeez, H.H. Morphological and Morphometric Study of the Development of Seminiferous Epithelium of Donkey (Equus asinus) from Birth to Maturity. J. Cytol. Histol. 2015, 6, 1. [Google Scholar]
- Rota, A.; Puddu, B.; Sabatini, C.; Panzani, D.; Lainé, A.L.; Camillo, F. Reproductive parameters of donkey jacks undergoing puberty. Anim. Reprod. Sci. 2018, 192, 119–125. [Google Scholar] [CrossRef]
- Miró, J.; Flotats, A.; Rivera, M.; Ocaña, M.; Taberner, E.; Peña, A.; Rigau, T. OC3 Morphometry Characterisation of Catalan Donkey Spermatozoa and Identification of Sperm Morphometric Subpopulations. Reprod. Domest. Anim. 2006, 41, 103. [Google Scholar] [CrossRef]
- Miró, J.; Lobo, V.; Quintero-Moreno, A.; Medrano, A.; Peña, A.; Rigau, T. Sperm motility patterns and metabolism in Catalonian donkey semen. Theriogenology 2005, 63, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Kenney, R.M. Minimal contamination techniques for breeding mares: Techniques and priliminary findings. Proc. Am. Assoc. Equine Pract. 1975, 21, 327–336. [Google Scholar]
- Bompart, D.; Vázquez, R.F.; Gómez, R.; Valverde, A.; Roldán, E.R.S.; García-Molina, A.; Soler, C. Combined effects of type and depth of counting chamber, and rate of image frame capture, on bull sperm motility and kinematics. Anim. Reprod. Sci. 2019, 209, 106169. [Google Scholar] [CrossRef]
- Valverde, A.; Madrigal, M.; Caldeira, C.; Bompart, D.; de Murga, J.N.; Arnau, S.; Soler, C. Effect of frame rate capture frequency on sperm kinematic parameters and subpopulation structure definition in boars, analysed with a CASA-Mot system. Reprod. Domest. Anim. 2019, 54, 167–175. [Google Scholar] [CrossRef]
- Caldeira, C.; Hernández-Ibáñez, S.; Valverde, A.; Martin, P.; Herranz-Jusdado, J.G.; Gallego, V.; Asturiano, J.F.; Dzyuba, B.; Pšenička, M.; Soler, C. Standardization of sperm motility analysis by using CASA-Mot for Atlantic salmon (Salmo salar), European eel (Anguilla anguilla) and Siberian sturgeon (Acipenser baerii). Aquaculture 2019, 502, 223–231. [Google Scholar] [CrossRef]
- Del Gallego, R.; Sadeghi, S.; Blasco, E.; Soler, C.; Yániz, J.L.L.; Silvestre, M.A.A. Effect of chamber characteristics, loading and analysis time on motility and kinetic variables analysed with the CASA-mot system in goat sperm. Anim. Reprod. Sci. 2017, 177, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Basioura, A.; Tsousis, G.; Boscos, C.; Lymberopoulos, A.; Tsakmakidis, I. Method agreement between three different chambers for comparative boar semen computer-assisted sperm analysis. Reprod. Domest. Anim. 2019, 54, 41–45. [Google Scholar] [PubMed]
- Hoogewijs, M.K.; De Vliegher, S.P.; Govaere, J.L.; De Schauwer, C.; De Kruif, A.; Van Soom, A. Influence of counting chamber type on CASA outcomes of equine semen analysis. Equine Vet. J. 2012, 44, 542–549. [Google Scholar] [CrossRef]
- Lenz, R.W.; Kjelland, M.E.; Vonderhaar, K.; Swannack, T.M.; Moreno, J.F. A comparison of bovine seminal quality assessments using different viewing chambers with a computer-assisted semen analyzer. J. Anim. Sci. 2011, 89, 383–388. [Google Scholar] [CrossRef]
- Valverde, A.; Arnau, S.; García-Molina, A.; Bompart, D.; Campos, M.; Roldán, E.; Soler, C. Dog sperm swimming parameters analysed by computer-assisted semen analysis of motility reveal major breed differences. Reprod. Domest. Anim. 2019, 54, 795–803. [Google Scholar] [CrossRef]
- Palacín, I.; Vicente-Fiel, S.; Santolaria, P.; Yániz, J.L. Standardization of CASA sperm motility assessment in the ram. Small Rumin. Res. 2013, 112, 128–135. [Google Scholar] [CrossRef]
- Gloria, A.; Carluccio, A.; Contri, A.; Wegher, L.; Valorz, C.; Robbe, D. The effect of the chamber on kinetic results in cryopreserved bull spermatozoa. Andrology 2013, 1, 879–885. [Google Scholar] [CrossRef]
- Suarez, S.S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Shalgi, R.; Smith, T.T.; Yanagimachi, R. A Quantitative Comparison of the Passage of Capacitated and Uncapacitated Hamster Spermatozoa through the Uterotubal Junction. Biol. Reprod. 1992, 46, 419–424. [Google Scholar] [CrossRef]
- Soler, C.; Picazo-Bueno, J.; Micó, V.; Valverde, A.; Bompart, D.; Blasco, F.J.; Álvarez, J.G.; García-Molina, A. Effect of counting chamber depth on the accuracy of lensless microscopy for the assessment of boar sperm motility. Reprod. Fertil. Dev. 2018, 30, 924–934. [Google Scholar] [CrossRef]
- Soler, C.; García, A.; Contell, J.; Segervall, J.; Sancho, M. Kinematics and subpopulations’ structure definition of blue fox (Alopex lagopus) sperm motility using the ISAS® V1 CASA system. Reprod. Domest. Anim. 2014, 49, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Douglas-Hamilton, D.H.; Smith, N.G.; Kuster, C.E.; Vermeiden, J.P.W.; Althouse, G.C. Capillary-loaded particle fluid dynamics: Effect on estimation of sperm concentration. J. Androl. 2005, 26, 115–122. [Google Scholar]
- Valverde, A.; Areán, H.; Fernández, A.; Bompart, D.; García-Molina, A.; López-Viana, J.; Soler, C. Combined effect of type and capture area of counting chamber and diluent on Holstein bull sperm kinematics. Andrologia 2018, 51, 1–10. [Google Scholar] [CrossRef]
- Dresdner, R.D.; Katz, D.F. Relationships of Mammalian Sperm Motility and Morphology to Hydrodynamic Aspects of Cell Function1. Biol. Reprod. 1981, 25, 920–930. [Google Scholar] [CrossRef]
- Spiropoulos, J. Computerized semen analysis (CASA): Effect of semen concentration and chamber depth on measurements. Arch. Androl. 2001, 46, 37–42. [Google Scholar] [CrossRef]
- Rijsselaere, T.; Van Soom, A.; Maes, D. Effect of technical settings on canine semen motility parameters measured by the Hamilton-Thorne analyzer. Theriogenology 2003, 60, 1553–1568. [Google Scholar] [CrossRef]
- Contri, A.; Valorz, C.; Faustini, M.; Wegher, L.; Carluccio, A. Effect of semen preparation on casa motility results in cryopreserved bull spermatozoa. Theriogenology 2010, 74, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Makler, A. Sealed mini-chamber of variable depth for direct observation and extended evaluation of sperm motility under the influence of various gases. Hum. Reprod. 1991, 6, 1275–1278. [Google Scholar] [CrossRef]
- Buss, T.; Aurich, J.; Aurich, C. Evaluation of a portable device for assessment of motility in stallion semen. Reprod. Domest. Anim. 2019, 54, 514–519. [Google Scholar] [CrossRef]
- Iguer-ouada, M.; Verstegen, J.P. Evaluation of the “Hamilton thorn computer-based automated system” for dog semen analysis. Theriogenology 2019, 55, 733–749. [Google Scholar] [CrossRef]
- Hayden, S.S.; Blanchard, T.L.; Brinsko, S.P.; Varner, D.D.; Hinrichs, K.; Love, C.C. Theriogenology The “dilution effect” in stallion sperm. Theriogenology 2015, 83, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Varner, D.D.; Blanchard, T.L.; Love, C.L.; Garcia, M.C.; Kenney, R.M. Effects of semen fractionation and dilution ratio on equine spermatozoal motility parameters. Theriogenology 1987, 28, 709–723. [Google Scholar] [CrossRef]
- Makler, A.; Deutch, M.; Vilensky, A.; Palti, Y. Factors affecting sperm motility VIII. Velocity and survival of human spermatozoa as related to temperatures above zero. Int. J. Androl. 1981, 4, 559–569. [Google Scholar] [CrossRef]
- Mortimer, D.; Goel, N.; Shu, M.A. Evaluation of the CellSoft automated semen analysis system in a routine laboratory setting. Fertil. Steril. 1988, 50, 960–968. [Google Scholar] [CrossRef]
- Neuwinger, J.; Knuth, U.A.; Nieschlag, E. Evaluation of the Hamilton–Thorn 2030 motility analyser for routine semen analysis in an infertility clinic. Int. J. Androl. 1990, 13, 100–109. [Google Scholar] [CrossRef]
- Verstegen, J.; Iguer-Ouada, M.; Onclin, K. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology 2002, 57, 149–179. [Google Scholar] [CrossRef]
- Rota, A.; Magelli, C.; Panzani, D.; Camillo, F. Effect of extender, centrifugation and removal of seminal plasma on cooled-preserved Amiata donkey spermatozoa. Theriogenology 2008, 69, 176–185. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Chamber Type | Chamber | n | α | SEMα | β | SEMβ | VCLα | VCL25 | VCL50 | VCL100 | VCL150 | VCL200 | VCL250 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Disposable | |||||||||||||
| D4C10 | 14,451 | 247.6 | 0.97 | 18.43 | 0.31 | 229.79 | 118.46 | 171.26 | 205.92 | 218.97 | 225.8 | 230.00 | |
| D4C20 | 64,732 | 252.96 | 0.62 | 23.64 | 0.25 | 230.30 | 98.26 | 157.65 | 199.7 | 216.07 | 224.75 | 230.13 | |
| D4C20L | 76,812 | 225.71 | 0.49 | 21.18 | 0.18 | 207.72 | 96.74 | 147.76 | 182.62 | 195.98 | 203.02 | 207.37 | |
| Reusable | |||||||||||||
| Spk10 | 105,679 | 255.01 | 0.55 | 24.23 | 0.19 | 231.89 | 96.74 | 157.07 | 200.13 | 216.97 | 225.91 | 231.45 | |
| Spk20 | 172,918 | 278.46 | 0.46 | 31.80 | 0.15 | 248.36 | 78.04 | 147.41 | 202.60 | 225.26 | 237.52 | 245.20 |
| Chambers | Disposable | Reusable | |||
|---|---|---|---|---|---|
| D4C | D4C | D4CL | Spk | Spk | |
| Depth | 10 µm | 20 µm | 20 µm | 10 µm | 20 µm |
| VCL | 227.91 ± 1.19 d | 238.16 ± 0.83 c | 223.77 ± 0.77 d | 260.47 ± 0.78 b | 268.07 ± 0.54 a |
| VSL | 70.19 ± 0.67 c,d | 71.16 ± 0.45 c | 68.93 ± 0.42 d | 83.57 ± 0.45 a | 80.46 ± 0.3 b |
| VAP | 181.57 ± 0.8 a | 177.18 ± 0.52 b | 168.15 ± 0.5 c | 183.8 ± 0.48 a | 181.66 ± 0.33 a |
| LIN | 29.82 ± 0.27 a,b | 28.3 ± 0.18 c | 29.32 ± 0.19 b | 30.23 ± 0.18 a | 28.46 ± 0.12 a,b |
| STR | 37.67 ± 0.38 d | 38.09 ± 0.25 d | 39.06 ± 0.24 c | 42.61 ± 0.23 a | 41.47 ± 0.16 b |
| WOB | 79.1 ± 0.19 a | 73.72 ± 0.13 c | 74.39 ± 0.15 b | 70.19 ± 0.13 d | 67.5 ± 0.09 e |
| ALH | 1.17 ± 0.01 e | 1.26 ± 0.01 c | 1.22 ± 0.01 d | 1.39 ± 0.01 b | 1.44 ± 0.0049 a |
| BCF | 42.58 ± 0.2 a | 40.35 ± 0.14 c | 38 ± 0.14 d | 40.79 ± 0.12 b | 39.96 ± 0.08 c |
| Chamber | VCL | VSL | VAP | LIN | STR | WOB | ALH | BCF |
|---|---|---|---|---|---|---|---|---|
| D4C10 | ||||||||
| 1 | 237.0 ± 3.3 a | 71.96 ± 1.69 b | 186.6 ± 2.71 a | 28.94 ± 0.56 b,c | 37.37 ± 0.71 b,c | 77.73 ± 0.39 b | 1.20 ± 0.01 a | 41.26 ± 0.67 a |
| 2 | 226.9 ± 3.1 a,b,c | 66.70 ± 1.59 b | 180.39 ± 2.56 a,b | 28.03 ± 0.53 c | 35.51 ± 0.67 c | 78.79 ± 0.37 a | 1.18 ± 0.01 a | 41.62 ± 0.63 a |
| 3 | 218.56 ± 4.22 c,d | 66.62 ± 2.16 b | 177.59 ± 3.47 a,b,c | 29.63 ± 0.72 b,c | 36.63 ± 0.91 b,c | 80.53 ± 0.5 a | 1.14 ± 0.01 a | 42.02 ± 0.86 a |
| 4 | 233.54 ± 3.11 a,b | 71.18 ± 1.6b | 183.96 ± 2.56 a,b | 29.66 ± 0.53 b | 37.63 ± 0.67 b | 78.58 ± 0.37 a,b | 1.2 ± 0.01 a | 41.99 ± 0.63 a |
| 5 | 218.26 ± 4.22 c,d | 67.45 ± 2.16 b | 177.22 ± 3.47 a,b,c | 30.46 ± 0.72 a,b | 37.76 ± 0.91 b | 80.63 ± 0.5 a | 1.14 ± 0.01 a | 42.34 ± 0.86 a |
| 6 | 212.65 ± 4.09 d | 69.66 ± 2.1 b | 170.19 ± 3.37 c | 31.97 ± 0.7 a | 40.27 ± 0.88 a | 79.41 ± 0.49 a | 1.12 ± 0.01 a | 40.91 ± 0.83 a |
| 7 | 252.96 ± 16.8 a | 93.72 ± 8.61 a | 198.38 ± 13.83 a | 35.64 ± 2.87 a | 45.5 ± 3.61 a | 78.06 ± 1.99 a,b | 1.23 ± 0.06 a | 43.78 ± 3.42 a |
| D4C20 | ||||||||
| 1 | 238.94 ± 2.64 b | 68.96 ± 1.32 c | 171.01 ± 2.04 b | 26.99 ± 0.4 c | 37.94 ± 0.51 b | 70.54 ± 0.27 c | 1.29 ± 0.01 b | 38.41 ± 0.48 c |
| 2 | 215.94 ± 2.58 c | 65.06 ± 1.29 d | 162.94 ± 1.99 c | 28.2 ± 0.39 b | 37.66 ± 0.5 b | 74.28 ± 0.26 a | 1.17 ± 0.01 d | 36.79 ± 0.46 d |
| 3 | 241.81 ± 2.51 b | 70.26 ± 1.26 b,c | 180.81±1.94 a | 27.7 ± 0.38 b,c | 37 ± 0.48 b | 74.31 ± 0.26 a | 1.27 ± 0.01 b | 40.52 ± 0.45 b |
| 4 | 243.05 ± 2.38 b | 72.97 ± 1.19 b | 182.02±1.84 a | 28.35 ± 0.36 b | 37.88 ± 0.46 b | 74.47 ± 0.24 a | 1.27 ± 0.01 b | 40.88 ± 0.43 b |
| 5 | 244.19 ± 2.71 b | 73.77 ± 1.36 b | 182.1±2.09 a | 28.7 ± 0.4 b | 38.39± 0.52 b | 73.99 ± 0.28 a | 1.28 ± 0.01 b | 39.7 ± 0.49 b,c |
| 6 | 253.55 ± 3.51 a | 78.47 ± 1.76 a | 184.84±2.72 a | 30.13 ± 0.53 a | 41.1 ± 0.68 a | 72.75 ± 0.36 b | 1.33 ± 0.01 a | 41.51 ± 0.63 a |
| 7 | 222.69 ± 3.91 c | 63.82 ± 1.96 d | 169.34 ± 3.02 b,c | 26.61 ± 0.59 c | 34.95 ± 0.75 c | 74.77 ± 0.4 a | 1.22 ± 0.01 c | 38.54 ± 0.71 c |
| Capture Fields | VCL | VSL | VAP | LIN | STR | WOB | ALH | BCF |
|---|---|---|---|---|---|---|---|---|
| A | 227.53 ± 1.89 a | 70.55 ± 0.95 a | 169.65 ± 1.49 a | 29.54 ± 0.3 a | 39.71 ± 0.38 a | 74.03 ± 0.19 b | 1.24 ± 0.01 a | 38.6 ± 0.37 a |
| B | 219.63 ± 1.65 b | 65.43 ± 0.83 b | 164.74 ± 1.3 b | 28.28 ± 0.26 b | 37.83 ± 0.33 b | 74.12 ± 0.17 b | 1.21 ± 0.01 b | 37.68 ± 0.32 a |
| C | 222.51 ± 1.5 b | 69.13 ± 0.76 a | 167.8 ± 1.19 a,b | 29.52 ± 0.24 a | 39.1 ± 0.3 a | 74.58 ± 0.16 a | 1.21 ± 0.01 b | 37.91 ± 0.29 a |
| Chamber and Capture Field | VCL | VSL | VAP | LIN | STR | WOB | ALH | BCF |
|---|---|---|---|---|---|---|---|---|
| SpK10 | ||||||||
| A | 274.11 ± 2.31 a | 88.37 ± 1.28 a | 188.94 ± 1.69 a | 30.33 ± 0.36 a,b | 43.8 ± 0.48 a | 68.53 ± 0.22 b | 1.45 ± 0.01 a | 41.35 ± 0.39 a |
| B | 257.82 ± 1.24 b | 81.41 ± 0.69 b | 182.23 ± 0.91 b | 29.6 ± 0.19 b | 41.61 ± 0.26 b | 70.3 ± 0.12 a | 1.38 ± 0.0045 b | 39.84 ± 0.21 b |
| C | 257.6 ± 1.63 b | 83.48 ± 0.91 b | 182.8 ± 1.2 b | 30.69 ± 0.25 a | 43.07 ± 0.34 a | 70.53 ± 0.15 a | 1.37 ± 0.01 b | 40.3 ± 0.28 b |
| SpK20 | ||||||||
| A | 271.45 ± 1.76 a | 82.66 ± 0.93 a | 177.73 ± 1.25 b | 28.08 ± 0.26 b | 42.47 ± 0.34 a | 64.84 ± 0.15 c | 1.47 ± 0.01 a | 38.54 ± 0.29 b |
| B | 271.84 ± 1.11 a | 81.69 ± 0.59 a | 183.81 ± 0.79 a | 28.71 ± 0.16 a | 41.89 ± 0.22 a | 67.51 ± 0.1 b | 1.45 ± 0.0041 b | 39.42 ± 0.18 a |
| C | 263.56 ± 1.05 b | 77.72 ± 0.56 b | 180.56 ± 0.74 b | 27.94 ± 0.15 b | 40.27 ± 0.21 b | 68.18 ± 0.09 a | 1.42 ± 0.0038 c | 39.54 ± 0.17 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gacem, S.; Catalán, J.; Valverde, A.; Soler, C.; Miró, J. Optimization of CASA-Mot Analysis of Donkey Sperm: Optimum Frame Rate and Values of Kinematic Variables for Different Counting Chamber and Fields. Animals 2020, 10, 1993. https://doi.org/10.3390/ani10111993
Gacem S, Catalán J, Valverde A, Soler C, Miró J. Optimization of CASA-Mot Analysis of Donkey Sperm: Optimum Frame Rate and Values of Kinematic Variables for Different Counting Chamber and Fields. Animals. 2020; 10(11):1993. https://doi.org/10.3390/ani10111993
Chicago/Turabian StyleGacem, Sabrina, Jaime Catalán, Anthony Valverde, Carles Soler, and Jordi Miró. 2020. "Optimization of CASA-Mot Analysis of Donkey Sperm: Optimum Frame Rate and Values of Kinematic Variables for Different Counting Chamber and Fields" Animals 10, no. 11: 1993. https://doi.org/10.3390/ani10111993
APA StyleGacem, S., Catalán, J., Valverde, A., Soler, C., & Miró, J. (2020). Optimization of CASA-Mot Analysis of Donkey Sperm: Optimum Frame Rate and Values of Kinematic Variables for Different Counting Chamber and Fields. Animals, 10(11), 1993. https://doi.org/10.3390/ani10111993