Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Effects on Growth, Metabolism, and Body Composition of the Offspring

 , , , ,
, , , ,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethic Statement
2.2. Animals and Experimental Design
2.3. Assessment of Morphological and Homeostatic Features of Offspring during Early Postnatal Development
2.4. Assessment of Morphological and Homeostatic Features of Offspring during Juvenile Development
2.5. Evaluation of Body Composition and Organs Weights of Offspring at 60 and 180 Days-Old
2.6. Evaluation of the Oxidant/Antioxidant Status of the Offspring
2.7. Evaluation of the Metabolic Status of the Offspring
2.8. Evaluation of Fat Content and Fatty Acid Composition in Feed and Tissue Samples
2.9. Statistical Analysis
3. Results
3.1. Effects of Maternal Supplementation on Litter Characteristics and Prenatal Development of Offspring
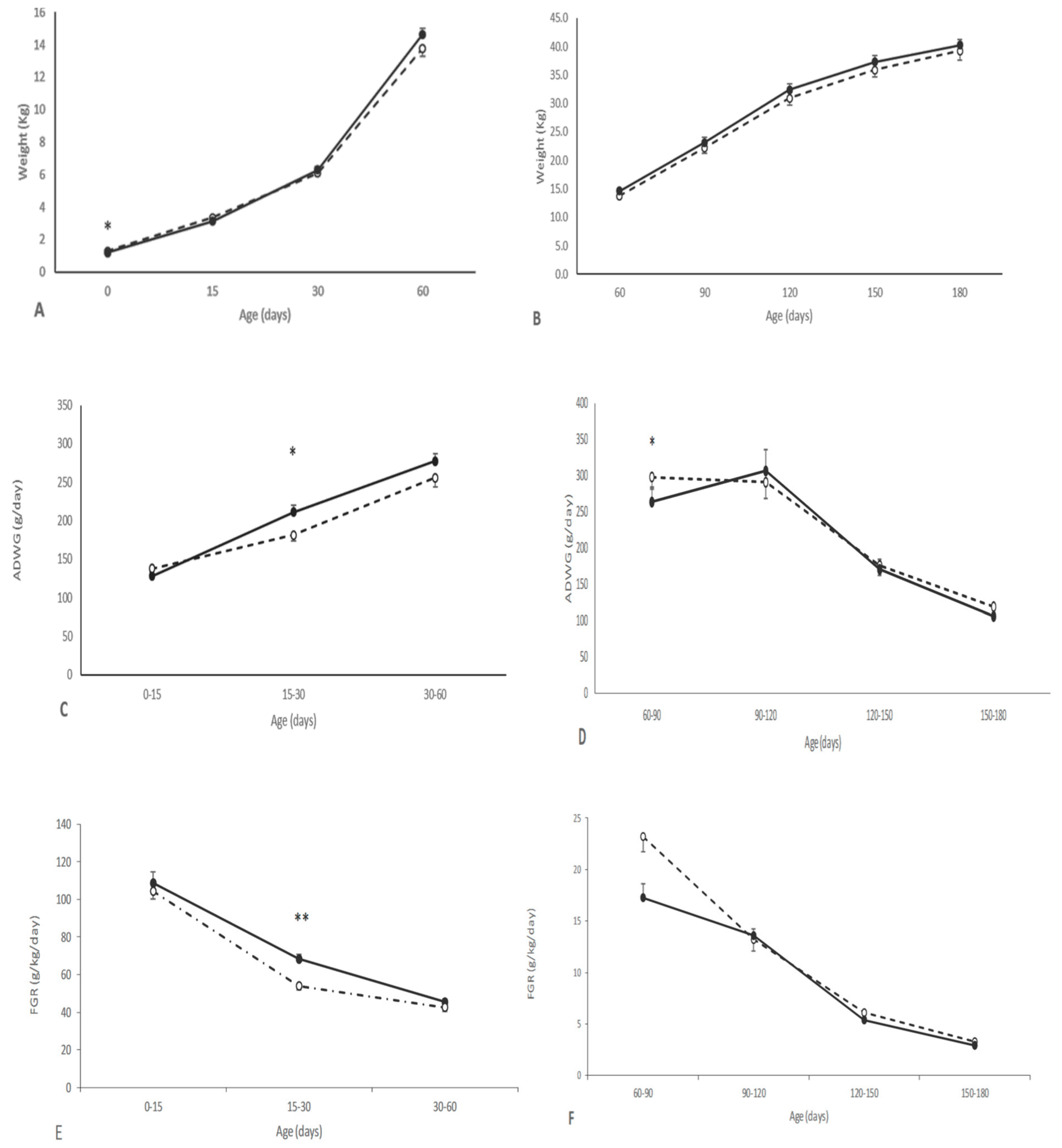
3.2. Effects of Maternal Supplementation on Postnatal Patterns of Growth and Development of Offspring
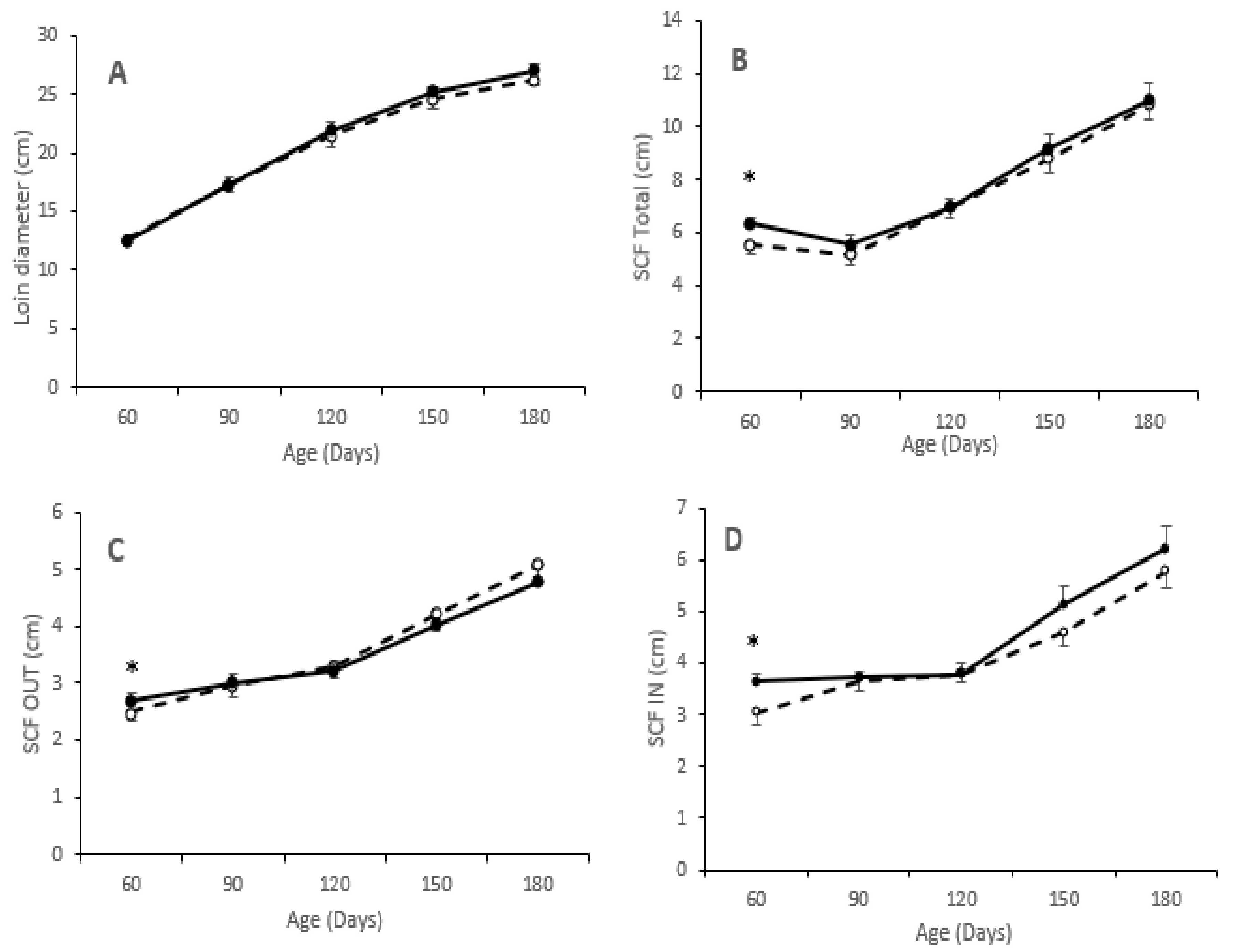
3.3. Effects of Maternal Supplementation on Body Composition and Adiposity of Offspring
3.4. Effects of Maternal Supplementation on the Antioxidant Capacity and Oxidative Stress of Offspring
3.5. Effects of Maternal Supplementation on the Metabolic Status of Offspring
3.6. Effects of Maternal Supplementation on Fatty Acid Composition of the Offspring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gonzalez-Bulnes, A.; Astiz, S.; Ovilo, C.; Lopez-Bote, C.J.; Torres-Rovira, L.; Barbero, A.; Ayuso, M.; Garcia-Contreras, C.; Vazquez-Gomez, M. Developmental Origins of Health and Disease in swine: Implications for animal production and biomedical research. Theriogenology 2016, 86, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Bernardini, E. Extra virgin olive oil’s polyphenols: Biological activities. Curr. Pharm. Des. 2011, 17, 786–804. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Barbero, A.; Pesantez, J.L.; Zinellu, A.; Berlinguer, F.; Gonzalez-Anover, P.; Gonzalez, J.; Encinas, T.; Torres-Rovira, L.; et al. Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Placental Gene Expression and Fetal Antioxidant Status, DNA-Methylation and Phenotype. Int. J. Mol. Sci. 2019, 20, 1187. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Pardo, Z.; Heras-Molina, A.; Encinas, T.; Torres-Rovira, L.; Astiz, S.; Nieto, R.; Ovilo, C.; Gonzalez-Bulnes, A.; et al. Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Hepatic Fat Accretion and Energy and Fatty Acids Profile of Fetal Tissues. Nutrients 2019, 11, 1534. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Gomez, M.; Garcia-Contreras, C.; Torres-Rovira, L.; Pesantez, J.L.; Gonzalez-Anover, P.; Gomez-Fidalgo, E.; Sanchez-Sanchez, R.; Ovilo, C.; Isabel, B.; Astiz, S.; et al. Polyphenols and IUGR pregnancies: Maternal hydroxytyrosol supplementation improves prenatal and early-postnatal growth and metabolism of the offspring. PLoS ONE 2017, 12, e0177593. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Gomez, M.; Heras-Molina, A.; Garcia-Contreras, C.; Pesantez-Pacheco, J.L.; Torres-Rovira, L.; Martinez-Fernandez, B.; Gonzalez, J.; Encinas, T.; Astiz, S.; Ovilo, C.; et al. Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Postnatal Growth, Metabolism and Body Composition of the Offspring. Antioxidants 2019, 8, 535. [Google Scholar] [CrossRef]
- Jones, M.L.; Mark, P.J.; Waddell, B.J. Maternal dietary omega-3 fatty acids and placental function. Reproduction 2014, 147, R143–R152. [Google Scholar] [CrossRef]
- Leskanich, C.O.; Noble, R.C. The comparative roles of polyunsaturated fatty acids in pig neonatal development. Br. J. Nutr. 1999, 81, 87–106. [Google Scholar] [CrossRef]
- Greenberg, J.A.; Bell, S.J.; Ausdal, W.V. Omega-3 Fatty Acid supplementation during pregnancy. Rev. Obstet. Gynecol. 2008, 1, 162–169. [Google Scholar]
- Innis, S.M. Fatty acids and early human development. Early Hum. Dev. 2007, 83, 761–766. [Google Scholar] [CrossRef]
- Akerele, O.A.; Cheema, S.K. A balance of omega-3 and omega-6 polyunsaturated fatty acids is important in pregnancy. J. Nutr. Intermed. Metab. 2016, 5, 23–33. [Google Scholar] [CrossRef]
- Haggarty, P. Fatty acid supply to the human fetus. Annu. Rev. Nutr. 2010, 30, 237–255. [Google Scholar] [CrossRef] [PubMed]
- Hachey, D.L. Benefits and risks of modifying maternal fat intake in pregnancy and lactation. Am. J. Clin. Nutr. 1994, 59, 454S–464S. [Google Scholar] [CrossRef]
- Jump, D.B.; Clarke, S.D. Regulation of gene expression by dietary fat. Annu. Rev. Nutr. 1999, 19, 63–90. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.K. Fatty acids in foods and their health implications; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar] [CrossRef]
- Prostek, A.; Gajewska, M.; Kamola, D.; Bałasińska, B. The influence of EPA and DHA on markers of inflammation in 3T3-L1 cells at different stages of cellular maturation. Lipids Health Dis. 2014, 13, 3. [Google Scholar] [CrossRef]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Statti, G.A.; Menichini, F. Biological and pharmacological activities of iridoids: Recent developments. Mini. Rev. Med. Chem. 2008, 8, 399–420. [Google Scholar] [CrossRef]
- Enke, U.; Seyfarth, L.; Schleussner, E.; Markert, U.R. Impact of PUFA on early immune and fetal development. Br. J. Nutr. 2008, 100, 1158–1168. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzym. 1978, 52, 302–310. [Google Scholar] [CrossRef]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Lopez-Bote, C.; Rey, A.; Ruiz, J.; Isabel, B.; Sanz Arias, R. Effect of feeding diets high in monounsaturated fatty acids and α-tocopheryl acetate to rabbits on resulting carcass fatty acid profile and lipid oxidation. Anim. Sci. 1997, 64, 177–186. [Google Scholar] [CrossRef]
- Segura, J.; Lopez-Bote, C.J. A laboratory efficient method for intramuscular fat analysis. Food Chem. 2014, 145, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Antequera, T.; Andres, A.I.; Petron, M.J.; Muriel, E. Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods. Anal. Chim. Acta 2004, 520, 201–205. [Google Scholar] [CrossRef]
- Daza, A.; Latorre, M.A.; Olivares, A.; López Bote, C.J. The effects of male and female immunocastration on growth performances and carcass and meat quality of pigs intended for dry-cured ham production: A preliminary study. Livest. Sci. 2016, 190, 20–26. [Google Scholar] [CrossRef]
- Segura, J.; Escudero, R.; Romero de Avila, M.D.; Cambero, M.I.; Lopez-Bote, C.J. Effect of fatty acid composition and positional distribution within the triglyceride on selected physical properties of dry-cured ham subcutaneous fat. Meat Sci. 2015, 103, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, A.J.; Pamplona, R.; Buffenstein, R.; Buttemer, W.A. Life and death: Metabolic rate, membrane composition, and life span of animals. Physiol. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef] [PubMed]
- Hulver, M.W.; Berggren, J.R.; Carper, M.J.; Miyazaki, M.; Ntambi, J.M.; Hoffman, E.P.; Thyfault, J.P.; Stevens, R.; Dohm, G.L.; Houmard, J.A.; et al. Elevated stearoyl-CoA desaturase-1 expression in skeletal muscle contributes to abnormal fatty acid partitioning in obese humans. Cell Metab. 2005, 2, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, A.M. The fetal circulation and its response to stress. J. Dev. Physiol. 1984, 6, 11–19. [Google Scholar]
- Meher, A.; Randhir, K.; Mehendale, S.; Wagh, G.; Joshi, S. Maternal Fatty Acids and Their Association with Birth Outcome: A Prospective Study. PLoS ONE 2016, 11, e0147359. [Google Scholar] [CrossRef]
- Olsen, S.F.; Hansen, H.S.; Jensen, B. Fish oil versus arachis oil food supplementation in relation to pregnancy duration in rats. Prostaglandins Leukot. Essent. Fat. Acids 1990, 40, 255–260. [Google Scholar] [CrossRef]
- Rodríguez Francisco, M.; García García, R.M.; Arias Álvarez, M.; Formoso Rafferty, N.; Millán, P.; López Tello, J.; Lorenzo, P.L.; González Bulnes, A.; García Rebollar, P. A diet supplemented with n-3 polyunsaturated fatty acids influences the metabolic and endocrine response of rabbit does and their offspring. J. Anim. Sci. 2017, 95, 2690–2700. [Google Scholar]
- Imhoff-Kunsch, B.; Briggs, V.; Goldenberg, T.; Ramakrishnan, U. Effect of n-3 long-chain polyunsaturated fatty acid intake during pregnancy on maternal, infant, and child health outcomes: A systematic review. Paediatr. Perinat. Epidemiol. 2012, 26, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Salvig, J.D.; Lamont, R.F. Evidence regarding an effect of marine n-3 fatty acids on preterm birth: A systematic review and meta-analysis. Acta Obstet. Gynecol. Scand. 2011, 90, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Elmes, M.; Green, L.R.; Poore, K.; Newman, J.; Burrage, D.; Abayasekara, D.R.; Cheng, Z.; Hanson, M.A.; Wathes, D.C. Raised dietary n-6 polyunsaturated fatty acid intake increases 2-series prostaglandin production during labour in the ewe. J. Physiol. 2005, 562, 583–592. [Google Scholar] [CrossRef]
- Elmes, M.; Tew, P.; Cheng, Z.; Kirkup, S.E.; Abayasekara, D.R.; Calder, P.C.; Hanson, M.A.; Wathes, D.C.; Burdge, G.C. The effect of dietary supplementation with linoleic acid to late gestation ewes on the fatty acid composition of maternal and fetal plasma and tissues and the synthetic capacity of the placenta for 2-series prostaglandins. Biochim. Biophys. Acta 2004, 1686, 139–147. [Google Scholar] [CrossRef]
- Lavery, A.; Lawlor, P.G.; Miller, H.M.; Magowan, E. The Effect of Dietary Oil Type and Energy Intake in Lactating Sows on the Fatty Acid Profile of Colostrum and Milk, and Piglet Growth to Weaning. Animals 2019, 9, 1092. [Google Scholar] [CrossRef]
- Luo, W.L.; Luo, Z.; Xu, X.; Zhao, S.; Li, S.H.; Sho, T.; Yao, J.; Zhang, J.; Xu, W.N.; Xu, J.X. The Effect of Maternal Diet with Fish Oil on Oxidative Stress and Inflammatory Response in Sow and New-Born Piglets. Oxid. Med. Cell Longev. 2019, 2019, 6765803. [Google Scholar] [CrossRef]
- Smit, M.N.; Spencer, J.D.; Patterson, J.L.; Dyck, M.K.; Dixon, W.T.; Foxcroft, G.R. Effects of dietary enrichment with a marine oil-based n-3 LCPUFA supplement in sows with predicted birth weight phenotypes on birth litter quality and growth performance to weaning. Animal 2015, 9, 471–480. [Google Scholar] [CrossRef]
- López-Vergé, S.; Gasa, J.; Farré, M.; Coma, J.; Bonet, J.; Solà-Oriol, D. Potential risk factors related to pig body weight variability from birth to slaughter in commercial conditions. Transl. Anim. Sci. 2018, 2, 383–395. [Google Scholar] [CrossRef]
- Valenzuela, R.; Videla, L.A. The importance of the long-chain polyunsaturated fatty acid n-6/n-3 ratio in development of non-alcoholic fatty liver associated with obesity. Food Funct. 2011, 2, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Polyunsaturated fatty acids and inflammatory processes: New twists in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A. Dietary n-3 PUFA and CVD: A review of the evidence. Proc. Nutr. Soc. 2014, 73, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Coletta, J.M.; Bell, S.J.; Roman, A.S. Omega-3 Fatty acids and pregnancy. Rev. Obstet. Gynecol. 2010, 3, 163–171. [Google Scholar]
- Ananthakrishnan, A.N.; Khalili, H.; Konijeti, G.G.; Higuchi, L.M.; de Silva, P.; Fuchs, C.S.; Willett, W.C.; Richter, J.M.; Chan, A.T. Long-term intake of dietary fat and risk of ulcerative colitis and Crohn’s disease. Gut 2014, 63, 776–784. [Google Scholar] [CrossRef]
- Rombaldi Bernardi, J.; de Souza Escobar, R.; Ferreira, C.F.; Pelufo Silveira, P. Fetal and neonatal levels of omega-3: Effects on neurodevelopment, nutrition, and growth. Sci. World J. 2012, 2012, 202473. [Google Scholar] [CrossRef]
- Gonzalez-Bulnes, A.; Astiz, S.; Ovilo, C.; Lopez-Bote, C.J.; Sanchez-Sanchez, R.; Perez-Solana, M.L.; Torres-Rovira, L.; Ayuso, M.; Gonzalez, J. Early-postnatal changes in adiposity and lipids profile by transgenerational developmental programming in swine with obesity/leptin resistance. J. Endocrinol. 2014, 223, M17–M29. [Google Scholar] [CrossRef]
- Hemant, P.; Lindsay, B. Stearoyl-CoA Desaturase: A Vital Checkpoint in the Development and Progression of Obesity. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 217–231. [Google Scholar] [CrossRef]
- Barbero, A.; Astiz, S.; Lopez-Bote, C.J.; Perez-Solana, M.L.; Ayuso, M.; Garcia-Real, I.; Gonzalez-Bulnes, A. Maternal malnutrition and offspring sex determine juvenile obesity and metabolic disorders in a swine model of leptin resistance. PLoS ONE 2013, 8, e78424. [Google Scholar] [CrossRef]
- Lee, J.M.; Lee, H.; Kang, S.; Park, W.J. Fatty Acid Desaturases, Polyunsaturated Fatty Acid Regulation, and Biotechnological Advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Lefaucheur, L.; Block, J.; Stabenow, B.; Pfuhl, R.; Otten, W.; Metges, C.C.; Kalbe, C. Limited and excess protein intake of pregnant gilts differently affects body composition and cellularity of skeletal muscle and subcutaneous adipose tissue of newborn and weanling piglets. Eur. J. Nutr. 2012, 51, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, C.; Stabenow, B.; Pfuhl, R.; Block, J.; Nurnberg, G.; Otten, W.; Metges, C.C.; Kalbe, C. Effects of limited and excess protein intakes of pregnant gilts on carcass quality and cellular properties of skeletal muscle and subcutaneous adipose tissue in fattening pigs. J. Anim. Sci. 2012, 90, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Patience, J.F.; Rossoni-Serão, M.C.; Gutiérrez, N.A. A review of feed efficiency in swine: Biology and application. J. Anim. Sci. Biotechnol. 2015, 6. [Google Scholar] [CrossRef]
- Smit, M.N.; Patterson, J.L.; Webel, S.K.; Spencer, J.D.; Cameron, A.C.; Dyck, M.K.; Dixon, W.T.; Foxcroft, G.R. Responses to n-3 fatty acid (LCPUFA) supplementation of gestating gilts, and lactating and weaned sows. Animal 2013, 7, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Roden, M.; Price, T.B.; Perseghin, G.; Petersen, K.F.; Rothman, D.L.; Cline, G.W.; Shulman, G.I. Mechanism of free fatty acid-induced insulin resistance in humans. J. Clin. Invest. 1996, 97, 2859–2865. [Google Scholar] [CrossRef]
- Segovia, S.A.; Vickers, M.H.; Zhang, X.D.; Gray, C.; Reynolds, C.M. Maternal supplementation with conjugated linoleic acid in the setting of diet-induced obesity normalises the inflammatory phenotype in mothers and reverses metabolic dysfunction and impaired insulin sensitivity in offspring. J. Nutr. Biochem. 2015, 26, 1448–1457. [Google Scholar] [CrossRef]
- Flachs, P.; Rossmeisl, M.; Bryhn, M.; Kopecky, J. Cellular and molecular effects of n-3 polyunsaturated fatty acids on adipose tissue biology and metabolism. Clin. Sci. 2009, 116, 1–16. [Google Scholar] [CrossRef]
- Torres-Rovira, L.; Gonzalez-Anover, P.; Astiz, S.; Caro, A.; Lopez-Bote, C.; Ovilo, C.; Pallares, P.; Perez-Solana, M.L.; Sanchez-Sanchez, R.; Gonzalez-Bulnes, A. Effect of an obesogenic diet during the juvenile period on growth pattern, fatness and metabolic, cardiovascular and reproductive features of swine with obesity/leptin resistance. Endocr. Metab. Immune Disord. Drug Targets 2013, 13, 143–151. [Google Scholar] [CrossRef]
- Torres-Rovira, L.; Astiz, S.; Caro, A.; Lopez-Bote, C.; Ovilo, C.; Pallares, P.; Perez-Solana, M.L.; Sanchez-Sanchez, R.; Gonzalez-Bulnes, A. Diet-induced swine model with obesity/leptin resistance for the study of metabolic syndrome and type 2 diabetes. Sci. World J. 2012, 2012, 510149. [Google Scholar] [CrossRef]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Torres-Rovira, L.; Gonzalez, J.; Porrini, E.; Gonzalez-Colaço, M.; Isabel, B.; Astiz, S.; Gonzalez-Bulnes, A. Characterization of ageing- and diet-related swine models of sarcopenia and sarcopenic obesity. Int. J. Mol. Sci. 2018, 19, 823. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, R.; González-Bulnes, A.; Garcia-Contreras, C.; Rodriguez-Rodriguez, A.E.; Astiz, S.; Vazquez-Gomez, M.; Pesantez, J.L.; Isabel, B.; Salido-Ruiz, E.; González, J.; et al. The Iberian pig fed with high-fat diet: A model of renal disease in obesity and metabolic syndrome. Int. J. Obes. 2020, 44, 457–465. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredients (g/Kg) | Group | |
|---|---|---|
| C | T | |
| Barley | 234.0 | 299.0 |
| Wheat | 300.1 | 192.5 |
| Wheat bran | 245.5 | 300.0 |
| Cookie Flour | 100.0 | 0.0 |
| Beet pulp | 50.0 | 50.0 |
| Sunflower flour (28%) | 31.5 | 68.0 |
| Soybean oil | 7.0 | 0.0 |
| Linseed oil | 0.0 | 40.0 |
| Sepiolite | 0.0 | 18.1 |
| Salt | 5.0 | 5.0 |
| Phosfate Monocalcium | 2.5 | 2.5 |
| Calcium carbonate | 16.5 | 17.5 |
| L—Lysine (500g/kg) | 4.2 | 3.8 |
| L-Threonine | 0.7 | 0.6 |
| Mineral and vitamin Premix | 3.0 | 3.0 |
| Days | Parameter | C | T | ||||
|---|---|---|---|---|---|---|---|
| Mean | Female | Male | Mean | Female | Male | ||
| 60 | Number of piglets | 18 | 7 | 11 | 17 | 8 | 9 |
| FRAP (µmol/mL) | 18.5 ± 1.18 | 17.2 ± 0.49 | 19.3 ± 1.91 | 21.3 ± 2.01 | 17.4 a ± 2.00 | 24.8 b ± 2.98 | |
| MDA (µmol/L) | 75.3 ± 5.43 | 81.4 ± 7.20 | 71.4 ± 7.65 | 65.4 ± 3.09 | 70.7 ± 4.66 | 60.8 ± 3.65 | |
| 180 | Number of piglets | 17 | 7 | 10 | 20 | 10 | 10 |
| FRAP (µmol/mL) | 38.1 ± 3.33 | 37.9 ± 7.18 | 38.3 ± 3.53 | 42.6 ± 4.74 | 49.2 ± 6.61 | 36.8 ± 6.61 | |
| MDA (µmol/L) | 27.2 ± 2.41 | 27.5 ± 4.76 | 27.1 ± 2.63 | 29.6 ± 1.96 | 31.4 ± 3.02 | 27.6 ± 2.42 | |
| Metabolites | 60 Days | 120 Days | 150 Days | 180 Days | ||||
|---|---|---|---|---|---|---|---|---|
| C | T | C | T | C | T | C | T | |
| GLU (MG/DL) | 100 ± 4.95 | 99.0 ± 5.63 | 97.4 ± 2.64 | 92.1 ± 2.26 | 105 ± 4.14 | 97.3 ± 2.12 | 99.3 ± 2.87 | 99.4 ± 3.31 |
| FRU (MG/DL) | 309 a ± 14.5 | 254 b ± 16.4 | 250 c ± 4.71 | 231 d ± 6.17 | 234 ± 5.47 | 225 ± 4.95 | 235 ± 4.84 | 222 ± 5.04 |
| CHO (MG/DL) | 107 ± 5.41 | 96.0 ± 3.79 | 111 a ± 2.85 | 101 b ± 1.88 | 109 c ± 2.33 | 99.5 d ± 2.30 | 106 e ± 2.98 | 97.0 f ± 3.13 |
| HDL (MG/DL) | 31.1 ± 1.45 | 28.8 ± 1.44 | 42.4 a ± 0.95 | 39.3 b ± 0.68 | 43.8 c ± 1.40 | 39.0 d ± 0.89 | 38.2 e ± 1.13 | 35.3 f ± 1.11 |
| LDL (MG/DL) | 78.2 a ± 5.10 | 63.2 b ± 2.98 | 66.4 c ± 2.60 | 59.7 d ± 1.97 | 64.7 e ± 3.01 | 57.2 f ± 1.96 | 73.2 g ± 3.23 | 64.8 h ± 2.47 |
| TG (MG/DL) | 50.3 a ± 5.81 | 69.8 b ± 4.74 | 63.8 ± 4.89 | 59.9 ± 4.14 | 52.4 ± 2.55 | 46.7 ± 2.77 | 46.3 ± 2.83 | 49.9 ± 3.51 |
| NEFA (MMOL/L) | 0.42 ± 0.02 | 0.43 ± 0.04 | 0.49 a ± 0.02 | 0.51 b ± 0.02 | 0.37 ± 0.02 | 0.38 ± 0.02 | 0.57 ± 0.05 | 0.52 ± 0.04 |
| UREA (MG/DL) | 25.3 ± 1.77 | 24.4 ± 1.58 | 26.3 a ± 1.92 | 21.6 b ± 0.99 | 29.1 ± 1.86 | 28.8 ± 2.17 | 29.0 ± 2.42 | 27.9 ± 3.14 |
| HP (MG/DL) | 55.6 ± 3.25 | 46.8 ± 4.74 | 37.7 ± 2.69 | 35.8 ± 2.44 | 42.9 ± 4.56 | 47.2 ± 3.77 | 44.4 ± 3.69 | 51.0 ± 4.20 |
| LAC (MG/DL) | 48.8 ± 5.15 | 44.6 ± 5.55 | 30.1 ± 2.64 | 21.6 ± 3.61 | 44.3 ± 6.95 | 36.9 ± 4.79 | 52.6 ± 4.74 | 49.0 ± 4.62 |
| BHB (MMOL/L) | 0.35 ± 0.03 | 0.32 ± 0.04 | 0.20 ± 0.02 | 0.25 ± 0.04 | 0.32 ± 0.03 | 0.30 ± 0.03 | 0.32 ± 0.03 | 0.29 ± 0.03 |
| Tissue | Fraction | Variable | C | T |
|---|---|---|---|---|
| SCF | Out | ∑n3 (g/100g FA) | 2.09 a ± 0.05 | 2.24 b ± 0.06 |
| ∑n6/∑n3 | 10.6 e ± 0.11 | 10.0 f ± 0.09 | ||
| DN6 | 0.007 c ± 0.00 | 0.009 d ± 0.000 | ||
| DI | 6.55 a ± 0.33 | 5.66 b ± 0.18 | ||
| In | ∑n6/∑n3 | 10.8 e ± 0.14 | 10.0 f ± 0.13 | |
| DN3 | 0.021 a ± 0.000 | 0.022 b ± 0.000 | ||
| DN6 | 0.007 e ± 0.000 | 0.008 f ± 0.000 | ||
| LD | Neutral | ∑n6/∑n3 | 9.86 e ± 0.11 | 9.18 f ± 0.10 |
| Polar | PUFA (g/100g FA) | 43.7 a ± 0.38 | 45.2 b ± 0.45 | |
| ∑n6 | 40.5 a ± 0.37 | 41.8 b ± 0.45 | ||
| BF | Neutral | ∑n6/∑n3 | 10.6 a ± 0.19 | 9.78 b ± 0.15 |
| Polar | DN6 | 0.32 a ± 0.04 | 0.35 b ± 0.04 | |
| Liver | Polar | ∑n3 (g/100g FA) | 2.14 a ± 0.16 | 2.55 b ± 0.09 |
| Tissue | Fraction | Variable | C | T |
|---|---|---|---|---|
| SCF | Out | ∑n3 UI DI | 1.19 a ± 0.03 0.88 a ± 0.01 5.11 c ± 0.11 | 1.10 b ± 0.03 0.86 b ± 0.01 4.64 d ± 0.10 |
| D9 | 0.61 c ± 0.003 | 0.60 d ± 0.004 | ||
| DN6 | 0.016 a ± 0.00 | 0.013 b ± 0.00 | ||
| In | SFA PUFA MUFA/SFA | 34.3 c ± 0.41 15.7 a ± 0.54 1.41 c ± 0.02 | 35.9 d ± 0.34 14.6 b ± 0.36 1.34 d ± 0.02 | |
| ∑n6/∑n3 | 13.7 a ± 0.20 | 15.1 b ± 0.31 | ||
| ∑n3 UI DI | 1.07 c ± 0.04 0.84 c ± 0.01 3.96 c ± 0.08 | 0.92 d ± 0.03 0.81 d ± 0.01 3.55 d ± 0.07 | ||
| VCF | ∑n3 UI DI | 0.95 a ± 0.03 0.73 a ± 0.01 2.11 a ± 0.05 | 0.91 b ± 0.03 0.72 b ± 0.01 1.99 b ± 0.03 | |
| LD | Neutral | ∑n6 ∑n6/∑n3 UI | 9.48 a ± 0.49 10.75 a ± 0.29 0.77 a ± 0.01 | 8.17 b ± 0.41 9.96 b ± 0.20 0.74 b ± 0.01 |
| BF | Polar | DI | 1.12 a ± 0.03 | 1.20 b ± 0.05 |
| Liver | Neutral | PUFA ∑n6 DN6 | 8.96 a ± 0.92 7.24 a ± 0.85 0.38 c ± 0.01 | 8.67 b ± 0.50 6.61 b ± 0.46 0.34 d ± 0.01 |
| Polar | SFA MUFA PUFA MUFA/SFA ∑n3 ∑n6 UI DI D9 | 62.0 a ± 0.64 23.2 a ± 0.50 14.9 c ± 0.67 0.37 c ± 0.01 1.68 e ± 0.08 13.2 a ± 0.60 0.69 e ± 0.02 0.63 a ± 0.02 0.25 c ± 0.01 | 63.8 b ± 0.42 21.7 b ± 0.32 14.5 d ± 0.44 0.34 d ± 0.01 1.55 f ± 0.04 12.9 b ± 0.41 0.65 f ± 0.02 0.56 b ± 0.01 0.23 d ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heras-Molina, A.; Pesantez-Pacheco, J.L.; Astiz, S.; Garcia-Contreras, C.; Vazquez-Gomez, M.; Encinas, T.; Óvilo, C.; Isabel, B.; Gonzalez-Bulnes, A. Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Effects on Growth, Metabolism, and Body Composition of the Offspring. Animals 2020, 10, 1946. https://doi.org/10.3390/ani10111946
Heras-Molina A, Pesantez-Pacheco JL, Astiz S, Garcia-Contreras C, Vazquez-Gomez M, Encinas T, Óvilo C, Isabel B, Gonzalez-Bulnes A. Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Effects on Growth, Metabolism, and Body Composition of the Offspring. Animals. 2020; 10(11):1946. https://doi.org/10.3390/ani10111946
Chicago/Turabian StyleHeras-Molina, Ana, José Luis Pesantez-Pacheco, Susana Astiz, Consolación Garcia-Contreras, Marta Vazquez-Gomez, Teresa Encinas, Cristina Óvilo, Beatriz Isabel, and Antonio Gonzalez-Bulnes. 2020. "Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Effects on Growth, Metabolism, and Body Composition of the Offspring" Animals 10, no. 11: 1946. https://doi.org/10.3390/ani10111946
APA StyleHeras-Molina, A., Pesantez-Pacheco, J. L., Astiz, S., Garcia-Contreras, C., Vazquez-Gomez, M., Encinas, T., Óvilo, C., Isabel, B., & Gonzalez-Bulnes, A. (2020). Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Effects on Growth, Metabolism, and Body Composition of the Offspring. Animals, 10(11), 1946. https://doi.org/10.3390/ani10111946