Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets

 ,
, 
 ,
,  , , , and
, , , and 
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals, Housing, Experimental Design and Treatment
2.2. Chemical Analyses and In Vitro Digestibility Evaluation of Ctrl and Ch/Qu Diets
2.3. Effect of Ctrl and Ch/Qu Diet on Swine Intestinal Cell Viability
2.4. Collection of Fecal and Blood Samples
2.5. Zootechnical Evaluation
2.6. Total Phenolic Compounds, Urea and Ammonia in Feces
2.7. Blood Serum Analysis
2.8. Statistical Analysis
3. Results
3.1. Chemical Analyses and In Vitro Digestibility Evaluation of Experimental Ch/Qu and Ctrl Diets
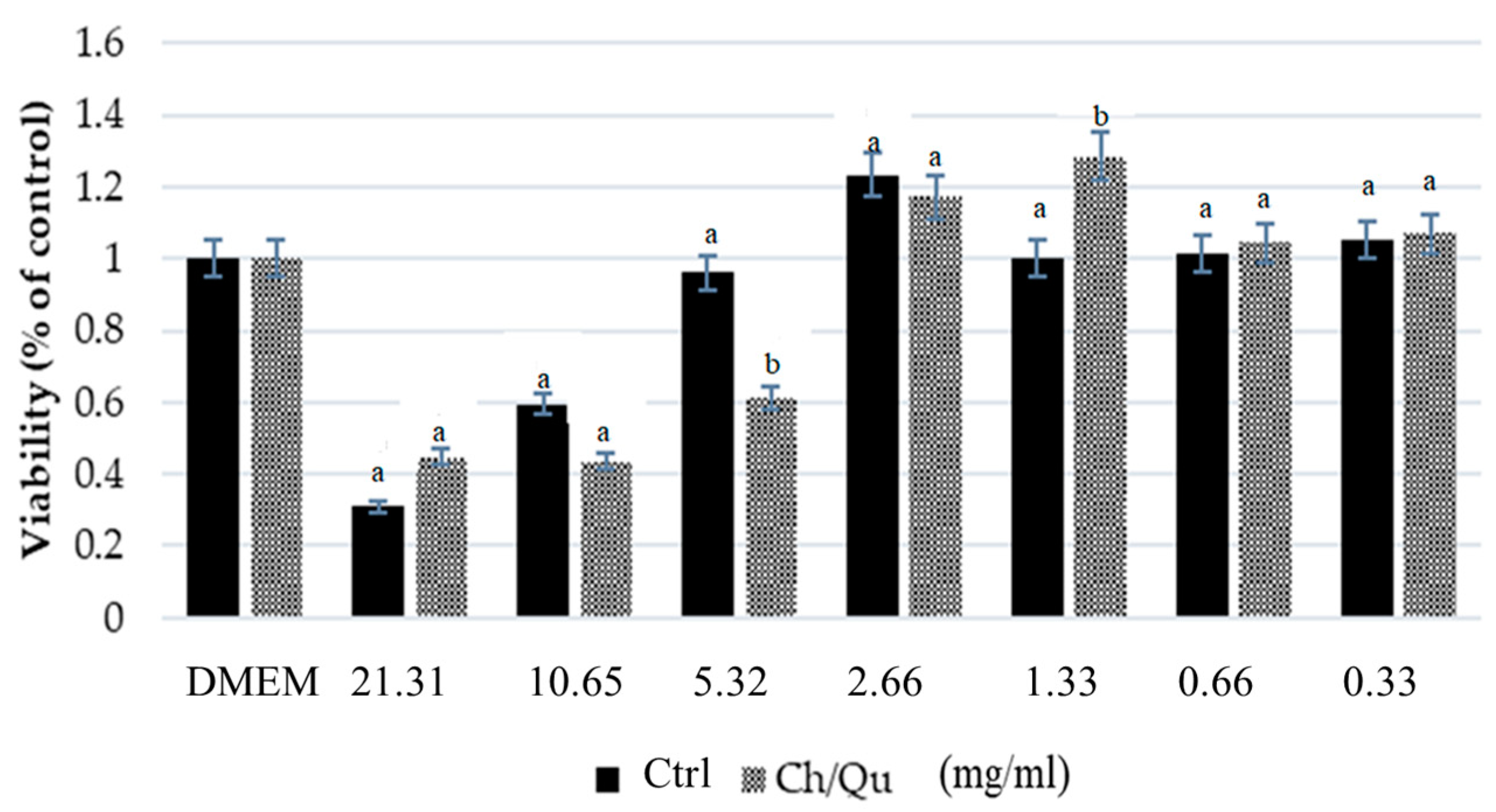
3.2. Swine Intestinal Cell Viability
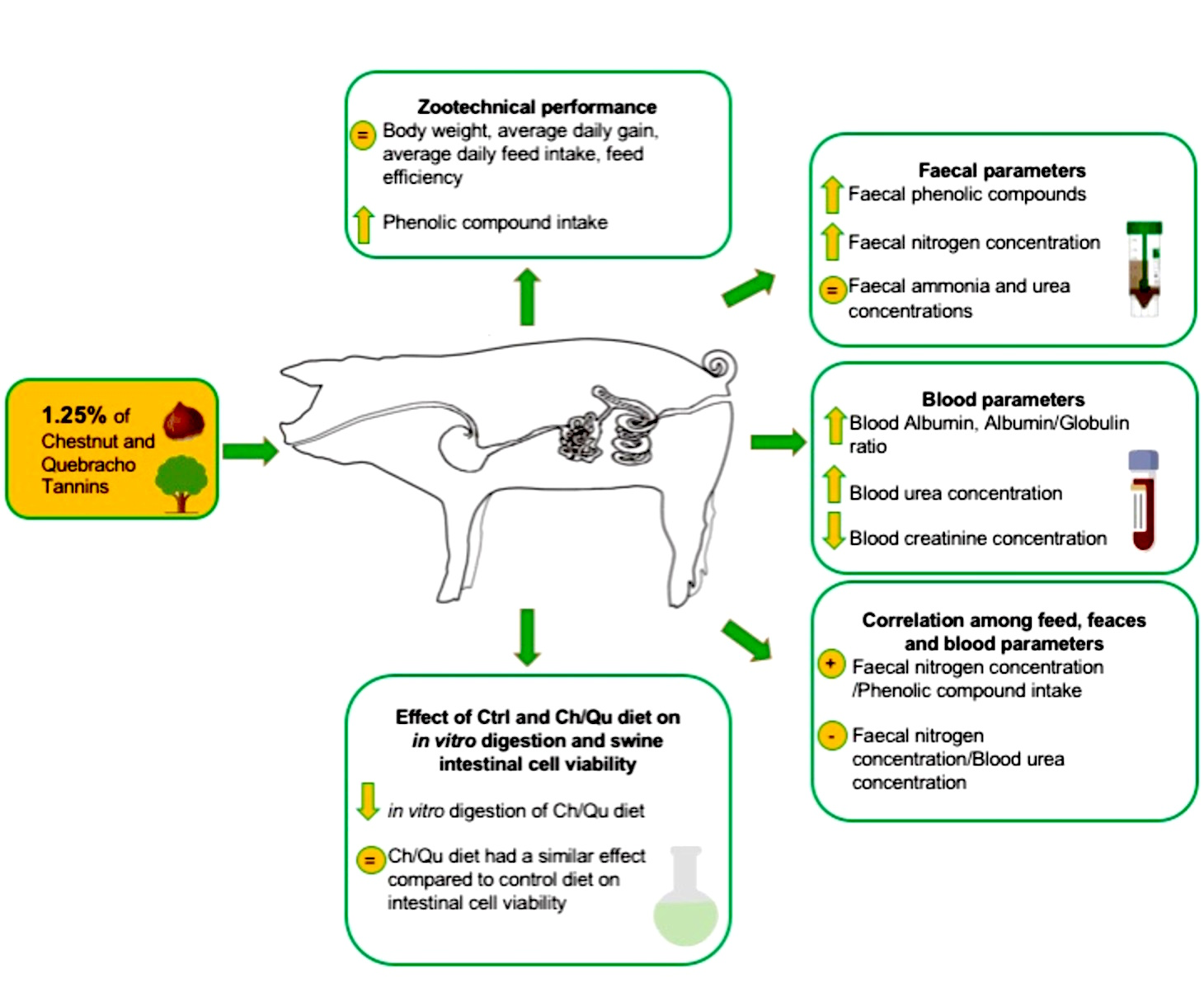
3.3. Zootechnical Performance
3.4. Blood Serum Metabolites
3.5. Influence of Dietary Treatment with Tannins on Feces
3.6. Influence of Dietary Treatment with Tannins on Correlation among Feed, Feces and Blood Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rossi, L.; Vagni, S.; Polidori, C.; Alborali, G.L.; Baldi, A.; Dell’Orto, V. Experimental induction of Escherichia coli diarrhoea in weaned piglets. J. Vet. Med. 2012, 2, 1–8. [Google Scholar] [CrossRef]
- Rossi, L.; Di Giancamillo, A.; Reggi, S.; Domeneghini, C.; Baldi, A.; Sala, V.; Dell’Orto, V.; Coddens, A.; Cox, E.; Fogher, C. Expression of verocytotoxic Escherichia coli antigens in tobacco seeds and evaluation of gut immunity after oral administration in mouse model. J. Vet. Sci. 2013, 14, 263–270. [Google Scholar] [CrossRef]
- Rossi, L.; Pinotti, L.; Agazzi, A.; Dell’Orto, V.; Baldi, A. Plant bioreactors for the antigenic hook-associated flgK protein expression. Ital. J. Anim. Sci. 2014, 13, 2939. [Google Scholar] [CrossRef]
- Rossi, L.; Dell’Orto, V.; Vagni, S.; Sala, V.; Reggi, S.; Baldi, A. Protective effect of oral administration of transgenic tobacco seeds against verocytotoxic Escherichia coli strain in piglets. Vet. Res. Commun. 2014, 38, 39–49. [Google Scholar] [CrossRef] [PubMed]
- EU Directive 1831/2003 of the European Parliament and of The Council on additives for use in animal nutrition. Off. J. Eur. Union 2003, 268, 29–43.
- Cromwell, G. Why and how antibiotics are used in swine production. Anim. Biotechnol. 2002, 13, 7–27. [Google Scholar] [CrossRef] [PubMed]
- Hejna, M.; Gottardo, D.; Baldi, A.; Dell’Orto, V.; Cheli, F.; Zaninelli, M.; Rossi, L. Review: Nutritional ecology of heavy metals. Animal 2018, 12, 2156–2170. [Google Scholar] [CrossRef] [PubMed]
- Hejna, M.; Moscatelli, A.; Stroppa, N.; Onelli, E.; Pilu, S.; Baldi, A.; Rossi, L. Bioaccumulation of heavy metals from wastewater through a Typha latifolia and Thelypteris palustris phytoremediation system. Chemosphere 2020, 241, 125018. [Google Scholar] [CrossRef] [PubMed]
- Hejna, M.; Moscatelli, A.; Onelli, E.; Baldi, A.; Pilu, S.; Rossi, L. Evaluation of concentration of heavy metals in animal rearing system. Ital. J. Anim. Sci. 2019, 18, 1372–1384. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC), European Food Safety Authority (EFSA), & European Medicines Agency (EMA). ECDC/EFSA/EMA Second Joint Report on the Integrated Analysis of the Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals: Joint Interagency Antimicrobial Consumption and Resistance Analysis (JIACRA) Report. EFSA J. 2017, 15, e04872. [Google Scholar]
- Sotira, S.; Dell’Anno, M.; Caprarulo, V.; Hejna, M.; Pirrone, F.; Callegari, M.L.; Tucci, T.V.; Rossi, L. Effects of tributyrin supplementation on growth performance, insulin, blood metabolites and gut microbiota in weaned piglets. Animals 2020, 10, 726. [Google Scholar] [CrossRef]
- Dell’Anno, M.; Hejna, M.; Sotira, S.; Caprarulo, V.; Reggi, S.; Pilu, R.; Miragoli, F.; Callegari, M.L.; Panseri, S.; Rossi, L. Evaluation of leonardite as a feed additive on lipid metabolism and growth of weaned piglets. Anim. Feed Sci. Tech. 2020, 266, 114519. [Google Scholar] [CrossRef]
- Onelli, E.; Moscatelli, A.; Gagliardi, A.; Zaninelli, M.; Bini, L.; Baldi, A.; Caccianiga, M.; Reggi, S.; Rossi, L. Retarded germination of Nicotiana tabacum seeds following insertion of exogenous DNA mimics the seed persistent behavior. PLoS ONE 2017, 12, e0187929. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, M.; Che, T.M.; Almeida, J.A.S.; Lee, J.J.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Dietary plant extracts alleviate diarrhea and alter immune responses of weaned pigs experimentally infected with a pathogenic Escherichia coli. J. Anim. Sci. 2013, 91, 5294–5306. [Google Scholar] [CrossRef] [PubMed]
- Dell’Anno, M.; Sotira, S.; Rebucci, R.; Reggi, S.; Castiglioni, B.; Rossi, L. In vitro evaluation of antimicrobial and antioxidant activities of algal extracts. Ital. J. Anim. Sci. 2020, 19, 103–113. [Google Scholar] [CrossRef]
- Reggi, S.; Giromini, C.; Dell’Anno, M.; Baldi, A.; Rebucci, R.; Rossi, L. In vitro digestion of chestnut and quebracho tannin extracts: Antimicrobial effect, antioxidant capacity and cytomodulatory activity in swine intestinal IPEC-J2 cells. Animals 2020, 10, 195. [Google Scholar] [CrossRef]
- Giromini, C.; Lovegrove, J.A.; Givens, D.I.; Rebucci, R.; Pinotti, L.; Maffioli, E.; Tedeschi, G.; Sundaram, T.S.; Baldi, A. In vitro-digested milk proteins: Evaluation of angiotensin-1-converting enzyme inhibitory and antioxidant activities, peptidomic profile, and mucin gene expression in HT29-MTX cells. J. Dairy Sci. 2019, 102, 10760–10771. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef]
- Cappai, M.G.; Wolf, P.; Pinna, W.; Kamphues, J. Dose-response relationship between dietary polyphenols from acorns and parotid gland hypertrophy in pigs. Food. Nutr. Sci. 2012, 3, 1261. [Google Scholar] [CrossRef]
- Hao, R.; Li, Q.; Zhao, J.; Li, H.; Wang, W.; Gao, J. Effects of grape seed procyanidins on growth performance, immune function and antioxidant capacity in weaned piglets. Livest. Sci. 2015, 178, 237–242. [Google Scholar] [CrossRef]
- Girard, M.; Bee, G. Invited review: Tannins as a potential alternative to antibiotics to prevent coliform diarrhea in weaned pigs. Animal 2020, 14, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Biagia, G.; Cipollini, I.; Paulicks, B.R.; Roth, F.X. Effect of tannins on growth performance and intestinal ecosystem in weaned piglets. Arch. Anim. Nutr. 2010, 64, 121–135. [Google Scholar] [CrossRef]
- Caprarulo, V.; Giromini, C.; Rossi, L. Review: Chestnut and quebracho tannins in pig nutrition: A review of the effects on performance and intestinal health. Animal 2020, in press. [Google Scholar]
- Girard, M.; Hu, D.; Pradervand, N.; Neuenschwander, S.; Bee, G. Chestnut extract but not sodium salicylate decrease the severity of diarrhea and enterotoxigenic Escherichia coli F4 shedding in artificially infected piglets. BioRxiv 2019, 15, 0214267. [Google Scholar] [CrossRef] [PubMed]
- Bilic-Sobot, D.; Kubale, V.; Skrlep, M.; Candek-Potokar, M.; Povse, M.; Fazarinc, G.; Skorjanc, D. Effect of hydrolysable tannins on intestinal morphology, proliferation and apoptosis in entire male pigs. Arch. Anim. Nutr. 2016, 70, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Candek-Potokar, M.; Skrlep, M.; Lukac, N.; Zamaratskaia, G.; Povse, M.; Bolta, S.; Kubale, V.; Bee, G. Hydrolysable tannin fed to entire male pigs affects intestinal production, tissue deposition and hepatic clearance of skatole. Vet. J. 2015, 204, 162–167. [Google Scholar] [CrossRef]
- Brus, M.; Bavdek, S.V.; Skok, J.; Škorjanc, D. Impact of supplementing pig diet with tannins on histological characteristics of small intestine and growth performance of fattening pigs. Acta Agric. Slov. Suppl. 2013, 4, 135–138. [Google Scholar]
- Girard, M.; Thanner, S.; Pradervand, N.; Hu, D.; Ollagnier, C.; Bee, G. Hydrolysable chestnut tannins for reduction of postweaning diarrhea: Efficacy on an experimental ETEC F4 model. PLoS ONE 2018, 13, e0197878. [Google Scholar] [CrossRef]
- Tretola, M.; Maghin, F.; Silacci, P.; Ampuero, S.; Bee, G. Effect of supplementing hydrolysable tannins to a grower–finisher diet containing divergent PUFA levels on growth performance, boar taint levels in back fat and intestinal microbiota of entire males. Animals 2019, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- EU. Directive 2010/63/EU of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union Off. 2010, L276, 33–79. [Google Scholar]
- EU. Commission Directive 2010/6/EU of 9 February 2010 amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council as regards mercury, free gossypol, nitrites and Mowrah, Bassia, Madhuca. Off. J. the Eur. Union 2010, 37, 29–32. [Google Scholar]
- National Research Council, Nutrient Requirements of Swine; National Academies Press: Washington DC, USA, 2012.
- EU Directive 2008/120/EC of 18 December 2008 laying down minimum standards for the protection of pigs. Off. J. Eur. Union 2008, 316, 36–38.
- EU Commission Regulation 152/2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union 2009, 54, 1–130.
- International Organization for Standardization. ISO-24333: Cereal and Cereal Products—Sampling; International Organization for Standardization: Geneva, Switzerland, 2009. [Google Scholar]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Attard, E.A. Rapid microtitre plate Folin-Ciocalteu method for the assessment of polyphenols. Open Life Sci. 2013, 8, 48–53. [Google Scholar] [CrossRef]
- Regmi, P.R.; Ferguson, N.S.; Zijlstra, R.T. In vitro digestibility techniques to predict apparent total tract energy digestibility of wheat in grower pigs. J. Anim. Sci. 2009, 87, 3620–3629. [Google Scholar] [CrossRef]
- Castrica, M.; Rebucci, R.; Giromini, C.; Tretola, M.; Cattaneo, D.; Baldi, A. Total phenolic content and antioxidant capacity of agri-food waste and by-products. Ital. J. Anim. Sci. 2019, 18, 336–341. [Google Scholar] [CrossRef]
- SAS/STAT®. 9.4 User’s Guide, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Klem, T.B.; Bleken, E.; Morberg, H.; Thoresen, S.I.; Framstad, T. Hematologic and biochemical reference intervals for Norwegian crossbreed grower pigs. Vet. Clin. Pathol. 2010, 39, 221–226. [Google Scholar] [CrossRef]
- Friendship, R.M.; Lumsden, J.H.; McMillan, I.; Wilson, M.R. Hematology and biochemistry reference values for Ontario swine. Can. J. Comp. Med. 1984, 48, 390. [Google Scholar] [CrossRef]
- IZSLER, Parametri di Chimica Clinica: Valori Osservati in Suini di Diversa Età. Available online: https://www.izsler.it/pls/izs_bs/v3_s2ew_consultazione.mostra_pagina?id_pagina=1494 (accessed on 1 October 2020).
- Mueller-Harvey, I. Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Galassi, G.; Mason, F.; Rapetti, L.; Crovetto, G.; Spanghero, M. Digestibility and metabolic utilisation of diets containing chestnut tannins and their effects on growth and slaughter traits of heavy pigs. Ital. J. Anim. Sci. 2019, 18, 746–753. [Google Scholar] [CrossRef]
- Chapman, P.M. Defining hormesis: Comments on Calabrese and Baldwin. Hum. Exp. Toxicol. 2002, 21, 99–101. [Google Scholar] [CrossRef]
- Bee, G.; Silacci, P.; Ampuero-Kragten, S.; Candek-Potokar, M.; Wealleans, A.; Litten-Brown, J.; Salminen, J.; Mueller-Harvey, I. Hydrolysable tannin-based diet rich in gallotannins has a minimal impact on pig performance but significantly reduces salivary and bulbourethral gland size. Animal 2017, 11, 1617–1625. [Google Scholar] [CrossRef]
- Williams, A.; Krych, L.; Ahmad, H.; Nejsum, P.; Skovgaard, K.; Nielsen, D.; Thamsborg, S. A polyphenol-enriched diet and Ascaris suum infection modulate mucosal immune responses and gut microbiota composition in pigs. PLoS ONE 2017, 12, e0186546. [Google Scholar] [CrossRef]
- Carstensen, L.; Ersbøll, A.K.; Jensen, K.H.; Nielsen, J.P. Escherichia coli post-weaning diarrhea occurrence in piglets with monitored exposure to creep feed. Vet. Microbiol. 2005, 110, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Laine, T.M.; Lyytikäinen, T.; Yliaho, M.; Anttila, M. Risk factors for post-weaning diarrhoea on piglet producing farms in Finland. Acta Vet. Scand. 2008, 50, 21. [Google Scholar] [CrossRef] [PubMed]
- Etim, N.N.; Williams, M.E.; Akpabio, U.; Offiong, E.E.A. Haematological parameters and factors affecting their values. Agric. Sci. 2014, 2, 37–47. [Google Scholar] [CrossRef]
- Chedea, V.; Palade, L.; Pelmus, R.; Dragomir, C.; Taranu, I. Red grape pomace rich in polyphenols diet increases the antioxidant status in key organs kidneys, liver, and spleen of piglets. Animals 2019, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Abarghuei, M.J.; Rouzbehan, Y.; Salem, A.Z.M.; Zamiri, M.J. Nitrogen balance, blood metabolites and milk fatty acid composition of dairy cows fed pomegranate-peel extract. Livest. Sci. 2014, 164, 72–80. [Google Scholar] [CrossRef]
- Chen, G.J.; Song, S.D.; Wang, B.X.; Zhang, Z.F.; Peng, Z.L.; Guo, C.H.; Zhong, J.C.; Wang, Y. Effects of forage: Concentrate ratio on growth performance, ruminal fermentation and blood metabolites in housing-feeding yaks. Asian Austral. J. Anim. 2015, 28, 1736. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Lefter, N.A.; Gheorghe, A.; Untea, A.; Ropotă, M.; Grigore, D.-M.; Varzaru, I.; Toma, S.M. Evaluation of performance, nitrogen metabolism and tissue composition in barrows fed an n-3 PUFA-rich diet. Animals 2019, 9, 234. [Google Scholar] [CrossRef]
- Whang, K.; Easter, R. Blood urea nitrogen as an index of feed efficiency and lean growth potential in growing-finishing swine. Asian-Asian Austral. J. Anim. 2000, 13, 811–816. [Google Scholar] [CrossRef]
- Bravo, L.; Abia, R.; Eastwood, M.A.; Saura-Calixtol, F. Degradation of polyphenols (catechin and tannic acid) in the rat intestinal tract. Effect on coloic fermentation and faecal output. Br. J. Nutr. 1994, 71, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Choy, Y.; Quifer-Rada, P.; Holstege, D.; Frese, S.; Calvert, C.; Mills, D.; Lamuela-Raventos, R.; Waterhouse, A. Phenolic metabolites and substantial microbiome changes in pig feces by ingesting grape seed proanthocyanidins. Food Funct. 2014, 5, 2298–2308. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Jeong, J.; Kim, D.; Lee, Y.; Kwon, H.; Kim, Y. Influence of rapeseed meal on growth performance, blood profiles, nutrient digestibility and economic benefit of growing-finishing pigs. Asian Austral. J. Anim. 2015, 28, 1345–1353. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Ingredients 1,2,3,4,5, as % of Fed Basis | Ctrl | Ch/Qu |
|---|---|---|
| Barley meal | 25.15 | 25.00 |
| Wheat meal | 19.41 | 19.07 |
| Corn meal | 14.03 | 13.50 |
| Corn flakes | 4.85 | 4.80 |
| Soybean meal | 4.65 | 4.60 |
| Soybean protein | 4.11 | 4.10 |
| Bakery meal | 4.00 | 4.00 |
| Dextrose monohydrate | 3.50 | 3.50 |
| Wheat middlings | 4.32 | 4.30 |
| Fermented milk product | 3.00 | 3.00 |
| Fish meal | 2.50 | 2.50 |
| Milk whey powder | 2.50 | 2.50 |
| Coconut oil | 1.00 | 1.00 |
| Soy oil | 1.00 | 1.00 |
| Plasma, meal | 1.00 | 1.00 |
| Dicalcium phosphate | 0.85 | 0.80 |
| Animal fats, lard | 0.70 | 0.70 |
| L-Lysine | 0.50 | 0.50 |
| Acidity regulators 5 | 1.00 | 1.00 |
| Benzoic acid | 0.40 | 0.40 |
| L-Threonine | 0.34 | 0.34 |
| DL-Methionine | 0.35 | 0.35 |
| Sodium chloride | 0.26 | 0.24 |
| Vitamins | 0.24 | 0.24 |
| L-Valine (96.5%) | 0.14 | 0.14 |
| L-Tryptophan | 0.08 | 0.05 |
| Copper sulfate | 0.04 | 0.04 |
| Ch/Qu Tannins 6 | - | 1.25 |
| Principal Nutrients | Experimental Diets 1 | Tannin Additive | |
|---|---|---|---|
| Ctrl | Ch/Qu | Tannins 2 | |
| Moisture, % | 9.54 | 9.66 | 6.28 |
| Crude protein, % | 17.40 | 17.72 | 3.06 |
| Crude fiber, % | 3.27 | 3.45 | nd 3 |
| Crude fat, % | 4.47 | 4.94 | nd 3 |
| Ash, % | 5.32 | 5.71 | 1.29 |
| Phenolic compound, g TAE/kg | 0.79 ± 0.03 | 2.91 ± 0.06 | 715.05 ± 51.02 |
| Growth Performance | Treatments | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| Ctrl | Ch/Qu | Treatment | Day | Treatment × Day | ||
| Phenolic compound intake, g/d | 0.656 a | 2.341 b | 0.15 | <0.01 | <0.01 | <0.01 |
| ADFI, kg/d | 0.623 | 0.599 | 0.02 | 0.258 | <0.01 | 0.241 |
| BW, kg | 13.86 | 13.64 | 0.64 | 0.819 | <0.01 | 0.987 |
| ADG, kg/d | 0.292 | 0.284 | 0.02 | 0.729 | <0.01 | 0.651 |
| Feed:Gain, d | 1.98 | 2.04 | 0.07 | 0.510 | 0.040 | 0.140 |
| Blood 1 | Treatments 2 | SEM | p-Value | Reference Range | |
|---|---|---|---|---|---|
| Ctrl | Ch/Qu | Treatment | |||
| Total protein content, g/L | 52.88 | 52.18 | 1.89 | 0.798 | 44–74 [41,42,43] |
| Albumin, g/L | 19.30 a | 23.05 b | 0.88 | 0.006 | 19–39 [42] |
| Globulin, g/L | 33.58 | 29.80 | 1.55 | 0.053 | 19–41 [41,43] |
| A/G ratio | 0.58 a | 0.82 b | 0.04 | 0.001 | 0.50–2.2 [41,42,43] |
| ALT-GPT, IU/L | 38.33 | 33.82 | 2.51 | 0.218 | 36–117 [43] |
| AST-GOT, IU/L | 54.17 | 48.09 | 3.41 | 0.221 | 21–98 [42] |
| ALP, UI/L | 165.67 | 186.00 | 12.58 | 0.266 | 46–341 [43] |
| Glucose, mmol/L | 5.00 | 5.05 | 0.25 | 0.878 | 3.5–8.6 [41] |
| Urea, mmol/L | 2.18 a | 0.95 b | 0.19 | < 0.001 | 0.90–8.89 [41,42] |
| Creatinine, μmol/L | 78.92 a | 54.82 b | 4.18 | 0.001 | 67–172 [42] |
| Total bilirubin, umol/L | 1.98 | 1.67 | 0.13 | 0.107 | 0.9–3.4 [42] |
| Total cholesterol, mmol/L | 2.51 | 2.38 | 0.13 | 0.479 | 1.3–4.2 [41,42,43] |
| Triglycerides, mmol/L | 0.68 | 0.58 | 0.06 | 0.259 | 0.3–2.7 [41] |
| HDL, mmol/L | 0.77 | 0.75 | 0.03 | 0.667 | - |
| LDL, mmol/L | 1.60 | 1.52 | 0.10 | 0.532 | - |
| Calcium, mmol/L | 2.28 | 2.29 | 0.06 | 0.923 | 2.02–3.21 [42] |
| Phosphorus, mmol/L | 3.05 | 3.02 | 0.08 | 0.774 | 1.46–3.45 [42] |
| Magnesium, mmol/L | 0.85 | 0.80 | 0.02 | 0.214 | 0.9–1.2 [41] |
| Feces | Treatments 2 | SEM | p-Value | |
|---|---|---|---|---|
| Ctrl | Ch/Qu | Treatment | ||
| Phenolic compound, g TAE/kg | 0.34 a | 0.50 b | 0.05 | 0.047 |
| Nitrogen, g/kg 3 | 36.02 a | 41.35 b | 1.00 | 0.002 |
| Ammonia, g/100 g | 2.26 | 3.20 | 1.89 | 0.684 |
| Urea, g/100 g | 0.62 | 0.99 | 0.23 | 0.235 |
| Feces | Parameter | Spearman r | p-Value |
|---|---|---|---|
| Fecal nitrogen, g/kg DM | Blood urea, mmol/L | −0.86 | 0.014 |
| Phenolic compound intake, g/d | 0.89 | 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caprarulo, V.; Hejna, M.; Giromini, C.; Liu, Y.; Dell’Anno, M.; Sotira, S.; Reggi, S.; Sgoifo-Rossi, C.A.; Callegari, M.L.; Rossi, L. Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets. Animals 2020, 10, 1945. https://doi.org/10.3390/ani10111945
Caprarulo V, Hejna M, Giromini C, Liu Y, Dell’Anno M, Sotira S, Reggi S, Sgoifo-Rossi CA, Callegari ML, Rossi L. Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets. Animals. 2020; 10(11):1945. https://doi.org/10.3390/ani10111945
Chicago/Turabian StyleCaprarulo, Valentina, Monika Hejna, Carlotta Giromini, Yanhong Liu, Matteo Dell’Anno, Stefania Sotira, Serena Reggi, Carlo Angelo Sgoifo-Rossi, Maria Luisa Callegari, and Luciana Rossi. 2020. "Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets" Animals 10, no. 11: 1945. https://doi.org/10.3390/ani10111945
APA StyleCaprarulo, V., Hejna, M., Giromini, C., Liu, Y., Dell’Anno, M., Sotira, S., Reggi, S., Sgoifo-Rossi, C. A., Callegari, M. L., & Rossi, L. (2020). Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets. Animals, 10(11), 1945. https://doi.org/10.3390/ani10111945