Evaluation of SCD, ACACA and FASN Mutations: Effects on Pork Quality and Other Production Traits in Pigs Selected Based on RNA-Seq Results

 , , , ,
, , , ,  , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Methods
2.2. RNA-Sequencing, Differentially Expressed Gene (DEG) and Variant Calling Analyses
2.3. Genotyping
2.4. Statistical Analyses
3. Results
3.1. Animals
3.2. Potential Genetic Marker Selection
3.3. ACACA, FASN, and SCD Mutation Frequency Analysis
3.4. Effect of FASN c.1328G > A Polymorphism on Pig Production Traits
3.5. Effect of ACACA c.-456T > C Polymorphism on Pig Production Traits
3.6. Effect of SCD c.*164A>G Polymorphism on Pig Production Traits
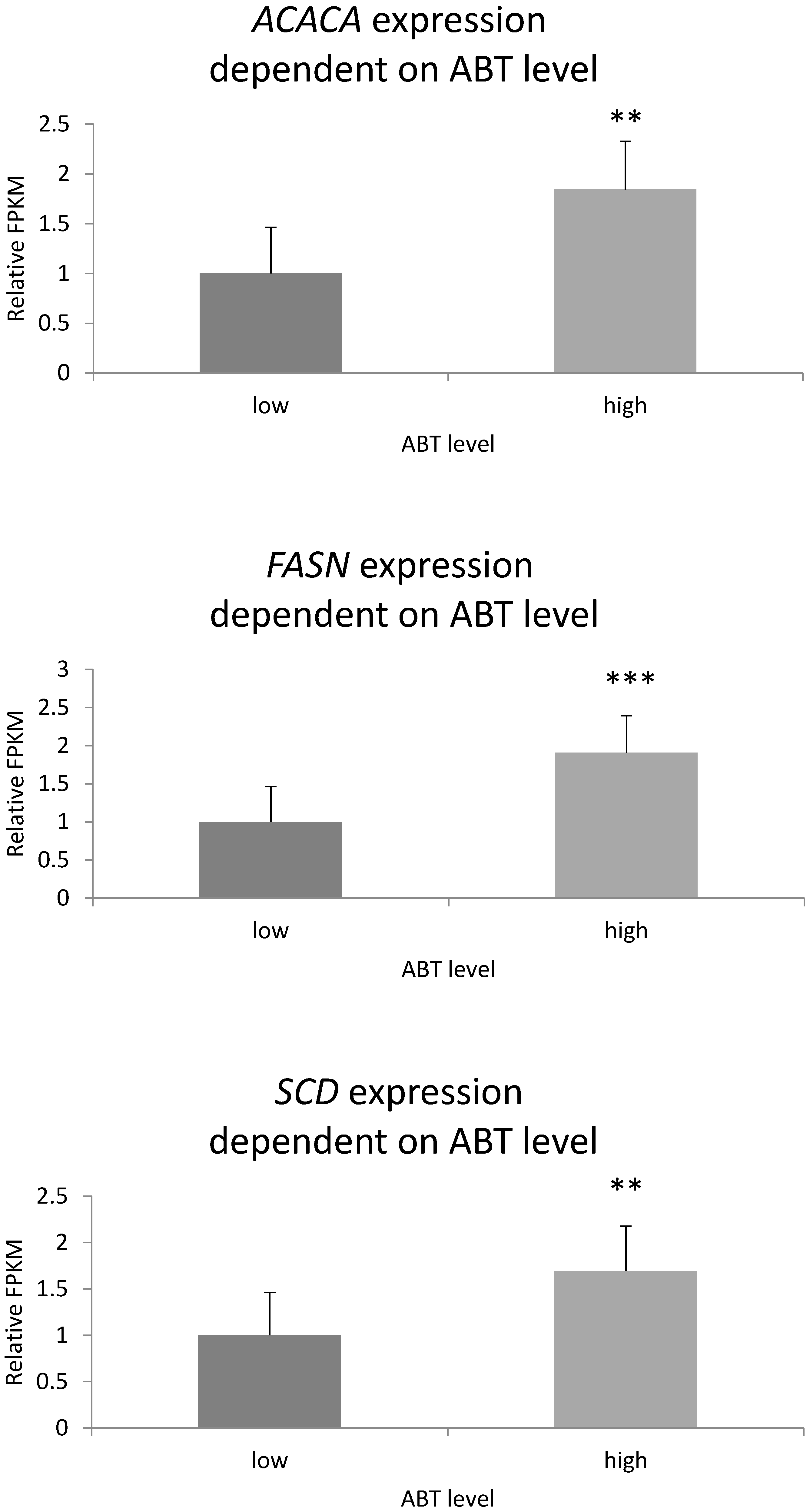
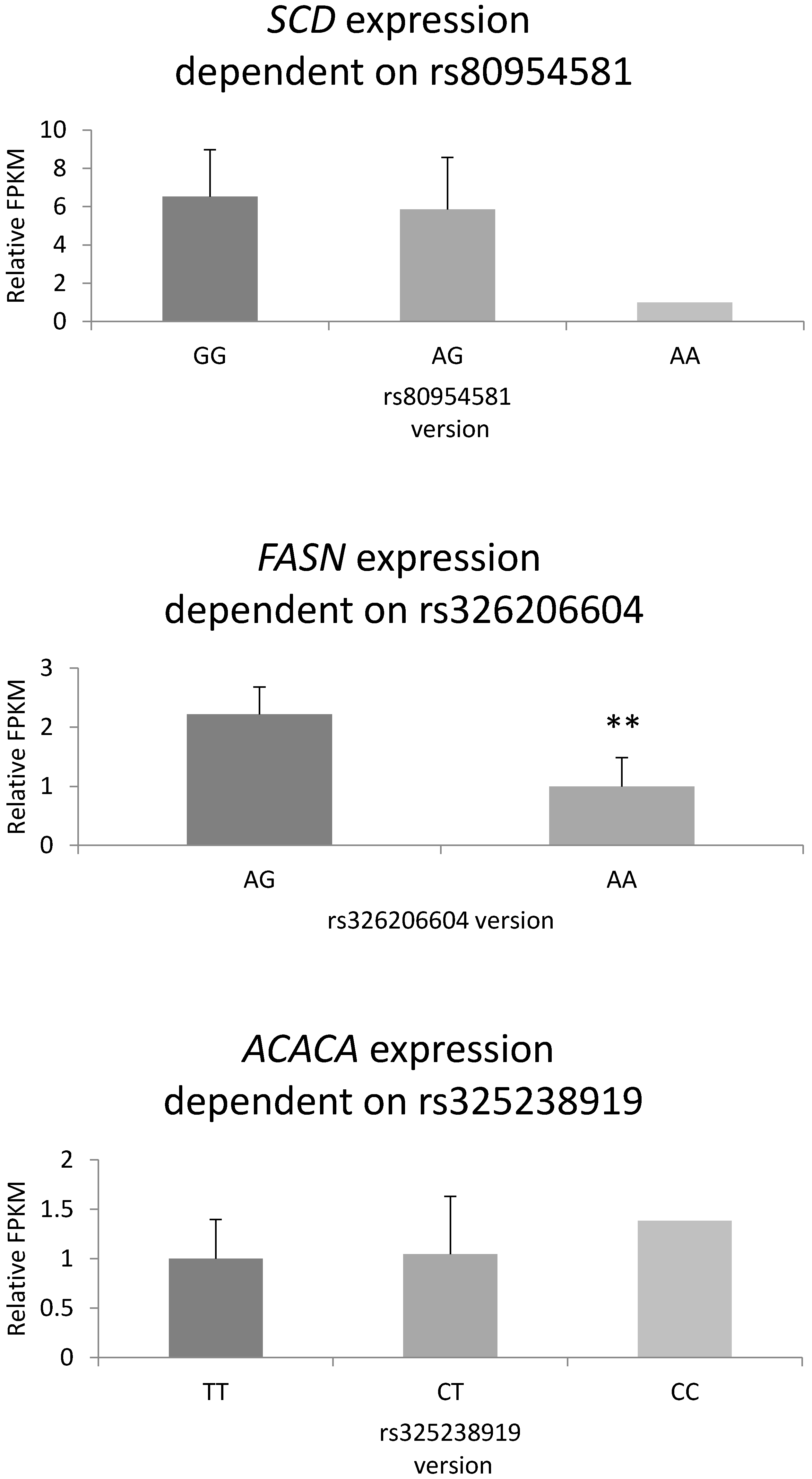
3.7. Gene Expression Level Depending on Genotype
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Różycki, M.; Tyra, M. Rules at evaluating the pigs in Pig Slaughter Testing Station. In State of Pig Breeding and Pig Evaluation Results; IZ PIB: Kraków, Poland, 2010; pp. 92–117. [Google Scholar]
- Ishii, K.; Arata, S.; Ohnishi, C.; Livestock, N.; Livestock, N.; Station, M. Estimates of genetic parameters for meat quality and carcass traits in Duroc pigs. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 11–16 February 2018; p. 408. [Google Scholar]
- Tomaschunas, M.; Zörb, R.; Fischer, J.; Köhn, E.; Hinrichs, J.; Busch-Stockfisch, M. Changes in sensory properties and consumer acceptance of reduced fat pork Lyon-style and liver sausages containing inulin and citrus fiber as fat replacers. Meat Sci. 2013, 95, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.S.; Zhu, L.G.; McKeith, F.K. Marbling effects on quality characteristics of pork loin chops: Consumer purchase intent, visual and sensory characteristics. Meat Sci. 2001, 59, 153–163. [Google Scholar] [CrossRef]
- Weiss, J.; Gibis, M.; Schuh, V.; Salminen, H. Advances in ingredient and processing systems for meat and meat products. Meat Sci. 2010, 86, 196–213. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B. Influence of intramuscular fat content on the quality of pig meat—2. Consumer acceptability of m. longissimus lumborum. Meat Sci. 1999, 53, 67–72. [Google Scholar] [CrossRef]
- Font-i-Furnols, M.; Tous, N.; Esteve-Garcia, E.; Gispert, M. Do all the consumers accept marbling in the same way? The relationship between eating and visual acceptability of pork with different intramuscular fat content. Meat Sci. 2012, 91, 448–453. [Google Scholar] [CrossRef]
- Tyra, M.; Zak, G. Characteristics of the Polish Breeding Population of Pigs in Terms of Intramuscular Fat (IMF) Content of M. Longissimus Dorsi. Ann. Anim. Sci. 2010, 10, 241–248. [Google Scholar]
- Shi-Zheng, G.; Su-Mei, Z. Physiology, Affecting Factors and Strategies for Control of Pig Meat Intramuscular Fat. Recent Pat. Food Nutr. Agric. 2012, 1, 59–74. [Google Scholar]
- Bentley, G. The health effects of dietary unsaturated fatty acids. Nutr. Bull. 2007, 32, 82–84. [Google Scholar] [CrossRef]
- Voss, M.D.; Beha, A.; Tennagels, N.; Tschank, G.; Herling, A.W.; Quint, M.; Gerl, M.; Metz-Weidmann, C.; Haun, G.; Korn, M. Gene expression profiling in skeletal muscle of Zucker diabetic fatty rats: Implications for a role of stearoyl-CoA desaturase 1 in insulin resistance. Diabetologia 2005, 48, 2622–2630. [Google Scholar] [CrossRef]
- Maharani, D.; Park, H.B.; Lee, J.B.; Yoo, C.K.; Lim, H.T.; Han, S.H.; Lee, S.S.; Ko, M.S.; Cho, I.C.; Lee, J.H. Association of the gene encoding stearoyl-CoA desaturase (SCD) with fatty acid composition in an intercross population between Landrace and Korean native pigs. Mol. Biol. Rep. 2013, 40, 73–80. [Google Scholar] [CrossRef]
- Lim, K.S.; Kim, J.M.; Lee, E.A.; Choe, J.H.; Hong, K.C. A candidate single nucleotide polymorphism in the 3′ untranslated region of stearoyl-CoA desaturase gene for fatness quality and the gene expression in berkshire pigs. Asian-Australasian J. Anim. Sci. 2015, 28, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Henriquez-Rodriguez, E.; Bosch, L.; Tor, M.; Pena, R.N.; Estany, J. The effect of SCD and LEPR genetic polymorphisms on fat content and composition is maintained throughout fattening in Duroc pigs. Meat Sci. 2016, 121, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, D.; Quintanilla, R.; Varona, L.; Díaz, I.; Ramírez, O.; Pena, R.N.; Amills, M. Polymorphism of the pig acetyl-coenzyme A carboxylase α gene is associated with fatty acid composition in a Duroc commercial line. Anim. Genet. 2009, 40, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, M.; Nowacka-Woszuk, J.; Szydlowski, M.; Switonski, M. The ACACA and SREBF1 genes are promising markers for pig carcass and performance traits, but not for fatty acid content in the longissimus dorsi muscle and adipose tissue. Meat Sci. 2013, 95, 64–71. [Google Scholar] [CrossRef]
- Grzes, M.; Sadkowski, S.; Rzewuska, K.; Szydlowski, M.; Switonski, M. Pig fatness in relation to FASN and INSIG2 genes polymorphism and their transcript level. Mol. Biol. Rep. 2016, 43, 381–389. [Google Scholar] [CrossRef]
- Zappaterra, M.; Luise, D.; Zambonelli, P.; Mele, M.; Serra, A.; Costa, L.N.; Davoli, R. Association study between backfat fatty acid composition and SNPs in candidate genes highlights the effect of FASN polymorphism in large white pigs. Meat Sci. 2019, 156, 75–84. [Google Scholar] [CrossRef]
- Renaville, B.; Bacciu, N.; Lanzoni, M.; Corazzin, M.; Piasentier, E. Polymorphism of fat metabolism genes as candidate markers for meat quality and production traits in heavy pigs. Meat Sci. 2015, 110, 220–223. [Google Scholar] [CrossRef]
- Mitka, I.; Tyra, M.; Wojtylak-Jurkiewicz, E. Analiza efektów genów warunkujących poziom tłuszczu śródmięś niowego (IMF) u świń oraz możliwości ich zastosowania w pracach selekcyjnych—praca przeglądowa. Wiad. Zootech. 2016, 54, 150–159. [Google Scholar]
- Soxhlet, F. Die gewichtsanalytische Bestimmung des Milchfettes. Polytech. J. 1879, 232, 461–465. [Google Scholar]
- Grau, R.; Hamm, R. Eine einfache Methode zur Bestimmung der Wasserbindung im Muskel. Naturwissenschaften 1953, 40, 29–30. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Lonergan, S.M. Mechanisms of water-holding capacity of meat: The role of postmortem biochemical and structural changes. Meat Sci. 2005, 71, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Piórkowska, K.; Żukowski, K.; Tyra, M.; Szyndler-Nędza, M.; Szulc, K.; Skrzypczak, E.; Ropka-Molik, K. The Pituitary Transcriptional Response Related to Feed Conversion in Pigs. Genes (Basel) 2019, 10, 712. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Farré, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Roselló, L.; Albà, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef]
- Chang, T.H.; Huang, H.Y.; Hsu, J.B.K.; Weng, S.L.; Horng, J.T.; Huang, H. Da An enhanced computational platform for investigating the roles of regulatory RNA and for identifying functional RNA motifs. BMC Bioinform. 2013, 14 (Suppl. 2), S4. [Google Scholar] [CrossRef]
- Maharani, D.; Jo, C.-R.; Jeon, J.-T.; Lee, J.-H. Quantitative Trait Loci and Candidate Genes Affecting Fatty Acid Composition in Cattle and Pig. Korean J. Food Sci. Anim. Resour. 2011, 31, 325–338. [Google Scholar] [CrossRef][Green Version]
- Muñoz, M.; Bozzi, R.; García, F.; Núñez, Y.; Geraci, C.; Crovetti, A.; García-Casco, J.; Alves, E.; Škrlep, M.; Charneca, R.; et al. Diversity across major and candidate genes in European local pig breeds. PLoS ONE 2018, 13, 1–30. [Google Scholar] [CrossRef]
- Deaton, A.M.; Sulem, P.; Nioi, P.; Benonisdottir, S.; Ward, L.D.; Davidsson, O.B.; Lao, S.; Helgadottir, A.; Fan, F.; Jensson, B.O.; et al. A rare missense variant in NR1H4 associates with lower cholesterol levels. Commun. Biol. 2018, 1, 14. [Google Scholar] [CrossRef]
- Tempfli, K.; Kiss, B.; Szalai, K.; Simon, Z.; Pongrácz, L.; Bali Papp, Á. Differential expression of six genes in fat-type Hungarian Mangalica and other pigs. Arch. Anim. Breed. 2016, 59, 259–265. [Google Scholar] [CrossRef]
- De Souza, F.R.P.; Chiquitelli, M.G.; da Fonseca, L.F.S.; Cardoso, D.F.; da Silva Fonseca, P.D.; de Camargo, G.M.F.; Gil, F.M.M.; Boligon, A.A.; Tonhati, H.; Mercadante, M.E.Z.; et al. Associations of FASN gene polymorphisms with economical traits in Nellore cattle (Bos primigenius indicus). Mol. Biol. Rep. 2012, 39, 10097–10104. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Gui, L.; Khan, R.; Schreurs, N.M.; Xiaoyu, W.; Wu, S.; Mei, C.; Wang, L.; Ma, X.; Wei, D.; et al. Association between FASN gene polymorphisms ultrasound carcass traits and intramuscular fat in Qinchuan cattle. Gene 2018, 645, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.C.; Carpenter, Z.L. Eating quality of meat animal products and their fat content. In Fat Content and Composition of Animal Products; National Academy of Sciences: Washington, DC, USA, 1976; pp. 147–182. ISBN 978-0-309-02440-2. [Google Scholar]
- Monroy, M.; Prasher, S.; Ngadi, M.O.; Wang, N.; Karimi, Y. Pork meat quality classification using Visible/Near-Infrared spectroscopic data. Biosyst. Eng. 2010, 107, 271–276. [Google Scholar] [CrossRef]
- Karpiesiuk, K.; Kozera, W.; Bugnacka, D.; Falkowski, J. Wpływ warunków chowu tuczników na jakość mięsa i profil kwasów tłuszczowych w mięsniu najdłuższym grzbietu. Żywn. Nauka Technol. Jakość 2013, 3, 39–50. [Google Scholar]
- Strzyżewski, T.; Bilska, A.; Krysztofiak, K. Zaleznośc pomiędzy wartością pH mięsa. Nauka Przyr. Technol. 2008, 2, 129–161. [Google Scholar]
- Reddy Gangidi, R.; Proctor, A. Meat and Meat Products. In Infrared Spectroscopy for Food Quality Analysis and Control; Academic Press: Cambridge, MA, USA, 2009; ISBN 9780123741363. [Google Scholar]
- Watanabe, G.; Motoyama, M.; Nakajima, I.; Sasaki, K. Relationship between water-holding capacity and intramuscular fat content in Japanese commercial pork loin. Asian-Australasian J. Anim. Sci. 2018, 31, 914–918. [Google Scholar] [CrossRef]
- Sieczkowska, H.; Andrzejczuk, A.; Zybert, A.; Krzęcio-Nieczyporuk, E.; Antosik, K.; Tarczyński, K.; Koćwin-Podsiadła, M. Przydatność kryteriów diagnozujących klasy jakości mięsa wieprzowego do szacowania cech przydatności kulinarnej mięsa. Rocz. Nauk. Pol. Tow. Zootech. 2017, 13, 53–62. [Google Scholar]
- Ropka-Molik, K.; Knapik, J.; Pieszka, M. Frekwencja trzech mutacji typu SNP w genie karboksylazy acetylo-CoA (ACACA) u owiec rasy merynos polski. In Proceedings of the Bioróżnorodność Zwierząt Gospodarskich—Praktyczne Wykorzystanie—Teraźniejszość i Przyszłość, Balice, Poland, 15–17 October 2013. [Google Scholar]
- Steri, M.; Idda, M.L.; Whalen, M.B.; Orrù, V. Genetic variants in mRNA untranslated regions. Wiley Interdiscip. Rev. RNA 2018, 9, e1474. [Google Scholar] [CrossRef]
- Kucharski, M.; Kaczor, U. Desaturaza stearylo-CoA—regulator metabolizmu lipidów Stearoyl-CoA desaturase—the lipid metabolism regulator. Adv. Hyg. Exp. Med. 2014, 68, 334–342. [Google Scholar]
- Barrett, L.W.; Fletcher, S.; Wilton, S.D. Regulation of eukaryotic gene expression by the untranslated gene regions and other non-coding elements. Cell. Mol. Life Sci. 2012, 69, 3613–3634. [Google Scholar] [CrossRef] [PubMed]
- Pichon, X.; Wilson, L.A.; Stoneley, M.; Bastide, A.; King, H.A.; Somers, J.; Willis, A.E. RNA Binding Protein/RNA Element Interactions and the Control of Translation. Curr. Protein Pept. Sci. 2012, 13, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Matsuhashi, T.; Maruyama, S.; Uemoto, Y.; Kobayashi, N.; Mannen, H.; Abe, T.; Sakaguchi, S.; Kobayashi, E. Effects of bovine fatty acid synthase, stearoyl-coenzyme A desaturase, sterol regulatory element-binding protein 1, and growth hormone gene polymorphisms on fatty acid composition and carcass traits in Japanese Black cattle. J. Anim. Sci. 2011, 89, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Walkowiak, E.; Zakrzewska, I. Oznaczanie poziomu bilirubiny w tkance mięśniowej wieprzowiny metodą Jendrosika i Grafa. Med. Weter. 1976, 32, 502–504. [Google Scholar]
- Chwastowska-Siwiecka, I.; Skiepko, N.; Gugołek, A.; Pomianowski, J.F. Zmiany ubytków masy i właściwości fizykochemicznych mięsa króliczego przechowywanego chłodniczo w różnych warunkach modyfikowanej atmosfery. Wiad. Zootech. 2015, 3, 9–19. [Google Scholar]



{kind=link}
{kind=link}
| Gene | SNP | Gene Region | Endonuclease | Digested PCR Product | refSNP | Primers |
|---|---|---|---|---|---|---|
| SCD | A/G | 3′UTR | HpyCH4III | 72,46,39/111,46 | rs80954581 c.*164A > G | TGCTACCAGGATGCTAAAGATG CAAGAGAAGGACAATGGGACT |
| ACACA | T/C | 5′UTR | AluI | 126,51,14/101,51, 25,14 | rs325238919 c.-456T > C | CTCCCCTCTCTCAGCTCCAG GGCAGGAATGTTACAAAACG |
| FASN | G/A | Missense Arg433Gln | HpaII | 208/160 | rs326206604 c.1328G > A | CTGACCCCAAGCTCTTTGAC CTCACCTTCGAGGTCGCTCT |
| Index | Genotype | PLW | PUŁ | PL | Total | GLM | ||
|---|---|---|---|---|---|---|---|---|
| FASN | Breed | di∙bj | ||||||
| Water exudation (%) | GG | 39.8 ± 2.36 | 28.8 ± 2.50 | - | 31.6 ± 1.11 ab | * | *** | ns |
| AG | 34.8 ± 1.03 | 30.6 ± 0.93 | 35.6 ± 0.89 | 30.8 ± 0.87 a | ||||
| AA | 34.5 ± 2.00 | 31.2 ± 0.62 | 36.1 ± 1.04 | 33.2 ± 1.10 b | ||||
| Meat colour L* (lightness) | GG | 52.8 ± 0.72 a | 54.7 ± 1.51 | - | 54.9 ± 0.51 | ns | *** | ns |
| AG | 53.8 ± 0.31 ab | 54.8 ± 0.57 | 55.5 ± 0.32 | 54.5 ±0.40 | ||||
| AA | 54.5 ± 0.31 b | 55.2 ± 0.38 | 55.0 ± 0.38 | 55.0 ±0.51 | ||||
| Meat colour b* (yellowness) | GG | 2.27 ± 0.27 | 2.78 ± 0.48 | - | 3.00 ± 0.16 | ns | *** | ns |
| AG | 2.51 ± 0.11 | 2.85 ± 0.18 | 2.47 ± 0.09 | 2.99 ± 0.13 | ||||
| AA | 2.01 ± 0.23 | 2.99 ± 0.12 | 2.45 ± 0.11 | 2.91 ± 0.16 | ||||
| pH 45 min (longissimus dorsi) | GG | 6.35 ± 0.07 | 6.25 ± 0.08 b | - | 6.13 ± 0.03 a | ** | *** | ns |
| AG | 6.28 ± 0.05 | 6.23 ± 0.03 ab | 6.34 ± 0.02 A | 6.21 ± 0.03 ab | ||||
| AA | 6.35 ± 0.06 | 6.13 ± 0.02 a | 6.24 ± 0.02 B | 6.17 ± 0.03 b | ||||
| pH 45 min (semimembranosus) | GG | 6.33 ± 0.05 | 6.21 ± 0.07 | - | 6.32 ± 0.03 | ns | *** | ns |
| AG | 6.34 ± 0.02 | 6.13 ± 0.03 | 6.33 ± 0.02A | 6.34 ± 0.02 | ||||
| AA | 6.29 ± 0.05 | 6.12 ± 0.02 | 6.25 ± 0.02B | 6.29 ± 0.03 | ||||
| Carcass yield (%) | GG | 77.7 ± 0.36 a | 75.1 ± 0.07 B | - | 74.8 ± 0.14 A | ns | * | ns |
| AG | 77.2 ± 0.16 ab | 74.3 ± 0.08 A | 76.6 ± 0.05 A | 75.5 ± 0.11 B | ||||
| AA | 76.8 ± 0.31 b | 74.5 ± 0.20 A | 75.7 ± 0.04 B | 75.9 ± 0.14 B | ||||
| Average backfat thickness (cm) | GG | 1.28 ± 0.09 | 1.56 ± 0.04 | - | 1.53 ±0.14 A | ns | * | ns |
| AG | 1.41 ± 0.04 | 1.56 ± 0.06 | 1.20 ± 0.05 | 1.36 ± 0.11 B | ||||
| AA | 1.34 ± 0.08 | 1.46 ± 0.16 | 1.24 ± 0.04 | 1.27 ± 0.14 B | ||||
| Loin mass (kg) | GG | 6.56 ± 0.14 A | 5.06 ± 0.04 | - | 5.21 ±0.07 A | ns | *** | ns |
| AG | 6.19 ± 0.06 AB | 5.08 ± 0.05 | 5.89 ± 0.07 | 5.75 ± 0.06 B | ||||
| AA | 5.84 ± 0.12 B | 5.06 ± 0.11 | 6.00 ± 0.08 | 5.91 ± 0.07 B | ||||
| Average daily gain (g/day) | GG | 919 ± 36 | 739 ± 58 | - | 799 ± 21 A | ns | *** | ns |
| AG | 920 ± 16 | 774 ± 22 | 958 ± 17 | 886 ± 16 A | ||||
| AA | 955 ± 30 | 786 ± 14 | 938 ± 15 | 925 ± 21 B | ||||
| Feed conversion (kg/kg) | GG | 2.71 ± 0.07 | 3.35 ± 0.13 B | - | 2.99 ±0.05 A | ns | *** | ns |
| AG | 2.68 ± 0.03 | 3.06 ± 0.05 AB | 2.72 ± 0.03 | 2.80 ± 0.04 B | ||||
| AA | 2.73 ± 0.06 | 3.01 ± 0.03 A | 2.78 ± 0.03 | 2.81 ± 0.05 B | ||||
| Daily feed intake (kg) | GG | 2.49 ± 0.10 | 2.41 ± 0.13 | - | 2.35 ±0.05 A | ns | *** | ns |
| AG | 2.45 ± 0.04 | 2.34 ± 0.05 | 2.60 ± 0.03 | 2.46 ±0.04 A | ||||
| AA | 2.58 ± 0.08 | 2.34 ± 0.03 | 2.59 ± 0.04 | 2.57 ± 0.05 B | ||||
| Age at slaughter (day) | GG | 165 ± 5.35 | 203 ± 8.56 B | - | 181 ± 3.39 A | ns | *** | ns |
| AG | 169 ± 2.20 | 182 ± 3.20 A | 162 ± 3.22 | 171 ± 2.71 B | ||||
| AA | 162 ± 4.52 | 183 ± 2.13 AB | 167 ± 2.76 | 169 ± 3.41 B | ||||
| Index | Genotype | PLW | PUŁ | PL | Total | GLM | ||
|---|---|---|---|---|---|---|---|---|
| ACACA | Breed | di∙bj | ||||||
| IMF (%) | CC | - | 1.37 ± 0.15 a | - | 1.31 ± 0.10 a | * | *** | ns |
| CT | 1.34 ± 0.09 | 1.67 ± 0.07 b | 1.12 ± 0.02 | 1.58 ± 0.08 b | ||||
| TT | 1.34 ± 0.05 | 1.72 ± 0.03 b | 1.12 ± 0.02 | 1.59 ± 0.08 b | ||||
| Meat colour b* (yellowness) | CC | - | 2.50 ± 0.37 | - | 2.47 ± 0.30 | ns | *** | ns |
| CT | 2.40 ± 0.20 | 2.89 ± 0.16 | 2.64 ± 0.13 | 2.73 ± 0.15 | ||||
| TT | 2.42 ± 0.12 | 3.05 ± 0.13 | 2.42 ± 0.09 | 2.69 ± 0.12 | ||||
| pH 45 min (longissimus dorsi) | CC | - | 6.13 ± 0.07 | - | 6.15 ± 0.06 | ns | *** | ns |
| CT | 6.30 ± 0.05 | 6.14 ± 0.03 | 6.33 ± 0.03 a | 6.22 ± 0.03 | ||||
| TT | 6.30 ± 0.03 | 6.17 ± 0.02 | 6.26 ± 0.02 b | 6.23 ± 0.03 | ||||
| pH 45 min (semimembranosus) | CC | - | 6.16 ± 0.06 | - | 6.18 ± 0.05 | ns | *** | ns |
| CT | 6.35 ± 0.03 | 6.11 ± 0.03 | 6.34 ± 0.02 a | 6.21 ± 0.03 | ||||
| TT | 6.29 ± 0.02 | 6.13 ± 0.02 | 6.27 ± 0.02 b | 6.23 ± 0.02 | ||||
| Carcass yield (%) | CC | - | 74.1 ± 0.20 a | - | 74.8 ± 0.26 A | ns | * | ns |
| CT | 77.1 ± 0.26 | 74.5 ± 0.08 b | 76.2 ± 0.12 | 75.5 ± 0.13 B | ||||
| TT | 77.4 ± 0.16 | 74.5 ± 0.07 b | 76.3 ± 0.09 | 75.9 ± 0.11 B | ||||
| Average backfat thickness (cm) | CC | - | 1.53 ± 0.12 | - | 1.43 ± 0.10 | ns | *** | ns |
| CT | 1.33 ± 0.07 | 1.58 ± 0.05 | 1.17 ± 0.05 | 1.42 ± 0.05 | ||||
| TT | 1.34 ± 0.04 | 1.54 ± 0.04 | 1.25 ± 0.04 | 1.40 ± 0.04 | ||||
| Average daily gain (g/day) | CC | - | 739 ± 44 | - | 767 ± 38 A | ns | *** | ns |
| CT | 938 ± 26 | 776 ± 20 | 956 ± 20 | 857 ± 19 B | ||||
| TT | 921 ± 15 | 788 ± 15 | 961 ± 15 | 874 ± 16 B | ||||
| Daily feed intake (kg) | CC | - | 2.23 ± 0.10 | - | 2.27 ± 0.09 a | ns | *** | ns |
| CT | 2.51 ± 0.07 | 2.31 ± 0.04 | 2.58 ± 0.05 | 2.43 ± 0.04 b | ||||
| TT | 2.46 ± 0.04 | 2.38 ± 0.03 | 2.63 ± 0.03 | 2.47 ± 0.04 b | ||||
| Index | Genotype | PLW | PUŁ | PL | Total | GLM | ||
|---|---|---|---|---|---|---|---|---|
| SCD | Breed | di∙bj | ||||||
| IMF (%) | AA | - | - | - | - | ns | *** | ns |
| AG | 1.28 ± 0.11 | 1.72 ± 0.08 | 1.23 ± 0.07 | 1.58 ± 0.07 a | ||||
| GG | 1.36 ± 0.06 | 1.66 ± 0.06 | 1.13 ± 0.02 | 1.39 ± 0.05 b | ||||
| Water exudation (%) | AA | - | - | - | - | ns | *** | ns |
| AG | 32.4 ± 2.17 | 29.9 ± 0.91 | 35.2 ± 2.44 | 30.9 ± 1.18 | ||||
| GG | 35.6 ± 1.16 | 30.6 ± 0.64 | 35.8 ± 0.76 | 33.5 ± 0.93 | ||||
| Meat colour L* (lightness) | AA | - | - | - | - | * | ns | ns |
| AG | 53.1 ± 0.60 | 54.5 ± 0.54 | 54.5 ± 0.89 | 54.3 ± 0.55 a | ||||
| GG | 54.1 ± 0.32 | 55.6 ± 0.38 | 55.1 ± 0.28 | 55.1 ± 0.43 b | ||||
| Meat colour b* (yellowness) | AA | - | - | - | - | * | *** | ns |
| AG | 2.36 ± 0.24 | 2.69 ± 0.17 a | 2.11 ± 0.25 | 2.57 ± 0.18 a | ||||
| GG | 2.37 ± 0.13 | 3.11 ± 0.12 b | 2.43 ± 0.08 | 2.71 ± 0.14 b | ||||
| pH 45 min (longissimus dorsi) | AA | - | - | - | - | ns | *** | ns |
| AG | 6.31 ± 0.06 | 6.17 ± 0.03 | 6.25± 0.06 | 6.21± 0.04 | ||||
| GG | 6.29 ± 0.03 | 6.14 ± 0.02 | 6.30± 0.02 | 6.19± 0.03 | ||||
| Loin mass (kg) | AA | - | - | - | - | ns | *** | ns |
| AG | 6.39 ± 0.16a | 5.00 ± 0.05 | 5.64 ± 0.17 a | 5.32 ± 0.08 a | ||||
| GG | 6.09 ± 0.08b | 5.06 ± 0.04 | 6.03 ± 0.05 b | 5.64 ± 0.06 b | ||||
| Ham mass (kg) | AA | - | - | - | - | ns | *** | ns |
| AG | 9.05 ± 0.16a | 8.72 ± 0.08 | 9.28 ± 0.13 | 8.84 ± 0.09 a | ||||
| GG | 9.36 ± 0.09b | 8.80 ± 0.05 | 9.26 ± 0.04 | 9.09 ± 0.07 b | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piórkowska, K.; Małopolska, M.; Ropka-Molik, K.; Szyndler-Nędza, M.; Wiechniak, A.; Żukowski, K.; Lambert, B.; Tyra, M. Evaluation of SCD, ACACA and FASN Mutations: Effects on Pork Quality and Other Production Traits in Pigs Selected Based on RNA-Seq Results. Animals 2020, 10, 123. https://doi.org/10.3390/ani10010123
Piórkowska K, Małopolska M, Ropka-Molik K, Szyndler-Nędza M, Wiechniak A, Żukowski K, Lambert B, Tyra M. Evaluation of SCD, ACACA and FASN Mutations: Effects on Pork Quality and Other Production Traits in Pigs Selected Based on RNA-Seq Results. Animals. 2020; 10(1):123. https://doi.org/10.3390/ani10010123
Chicago/Turabian StylePiórkowska, Katarzyna, Martyna Małopolska, Katarzyna Ropka-Molik, Magdalena Szyndler-Nędza, Angelika Wiechniak, Kacper Żukowski, Barry Lambert, and Mirosław Tyra. 2020. "Evaluation of SCD, ACACA and FASN Mutations: Effects on Pork Quality and Other Production Traits in Pigs Selected Based on RNA-Seq Results" Animals 10, no. 1: 123. https://doi.org/10.3390/ani10010123
APA StylePiórkowska, K., Małopolska, M., Ropka-Molik, K., Szyndler-Nędza, M., Wiechniak, A., Żukowski, K., Lambert, B., & Tyra, M. (2020). Evaluation of SCD, ACACA and FASN Mutations: Effects on Pork Quality and Other Production Traits in Pigs Selected Based on RNA-Seq Results. Animals, 10(1), 123. https://doi.org/10.3390/ani10010123