
From Strain Characterization to Field Authorization: Highlights on Bacillus velezensis Strain B25 Beneficial Properties for Plants and Its Activities on Phytopathogenic Fungi

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacillus velezensis Strain B25 Genomic Features Assessment
2.1.1. Genome Characterization
2.1.2. Gene Analysis
2.2. Assessment of the Inhibition of Fungal Pathogens by Bacillus velezensis Strain B25
2.2.1. B25 Culture Conditions
2.2.2. In Vitro Confrontations for Antagonism Potential Evaluation
2.2.3. Bioassay in Growth Chamber for Biocontrol Evaluation against F. graminearum Strain B377
2.3. Bacillus velezensis Strain B25 Metabolic Properties Assessment
2.3.1. Phosphate Solubilization Assay
2.3.2. Siderophores Detection Assay
2.3.3. Indolacetic Acid (IAA) Production Assay
2.3.4. Metabolic Profiling on Biolog GenIII Microplates
2.3.5. Antibiograms and Minimal Inhibitory Concentrations Evaluation
2.3.6. Growth Response under UV-C Stress
2.4. Bacillus velezensis Strain B25 Ecotoxicological Impact on Representative Organisms
2.4.1. Effects on Mortality and Reproduction of Eisenia fetida
2.4.2. Chronic Toxicity Effect on Daphnia magna
2.5. Statistical Analysis
3. Results
3.1. Characterization of the General Genomic Features of Strain B25
3.1.1. Genome Specificities
3.1.2. Phylogenetic Relationship of Bacillus velezensis Strain B25
3.1.3. Genes Involved in Biocontrol
3.1.4. Detection of Resistance Genes to Antibiotics
3.1.5. Polysaccharide Degrading Enzymes
3.2. Characterization of the Biocontrol Properties of Bacillus velezensis Strain B25
3.2.1. In Vitro Microbial Interactions Tests
- (i)
- A fast increase of the II was observed for the B25 challenge co-cultures against B278, B375, B376, B377, B381, B383, and B385 (Figure 3); with a 2-day latency phase followed by an exponential-like increase in 7 days, and finally a plateau with II reaching 0.7532, 0.8970, 0.8107, 0.6850, 0.6010, 0.6898, and 0.7243, respectively. For B381 strain, the plateau was not the maximum II reached (0.9626 at 7 days of culture followed by a decrease to the stationary phase). It could be explained by a strong antibiosis of B25 until 7 days before another phase of pathogen growth, however insufficient to resist B25.
- (ii)
- A longer latency phase (4 days) followed by a gradual increase of the II until the end of the experiment (15 days) was observed for the B25 challenge co-cultures against B378, B379, B380, B382, B384, and B386, with maximum II reaching 0.4052, 0.4516, 0.4663, 0.6010, 0.6854, and 0.8157, respectively (Figure 4).
3.2.2. In Planta Biocontrol Activities
3.3. Characterization of the Metabolic Properties of Strain B25
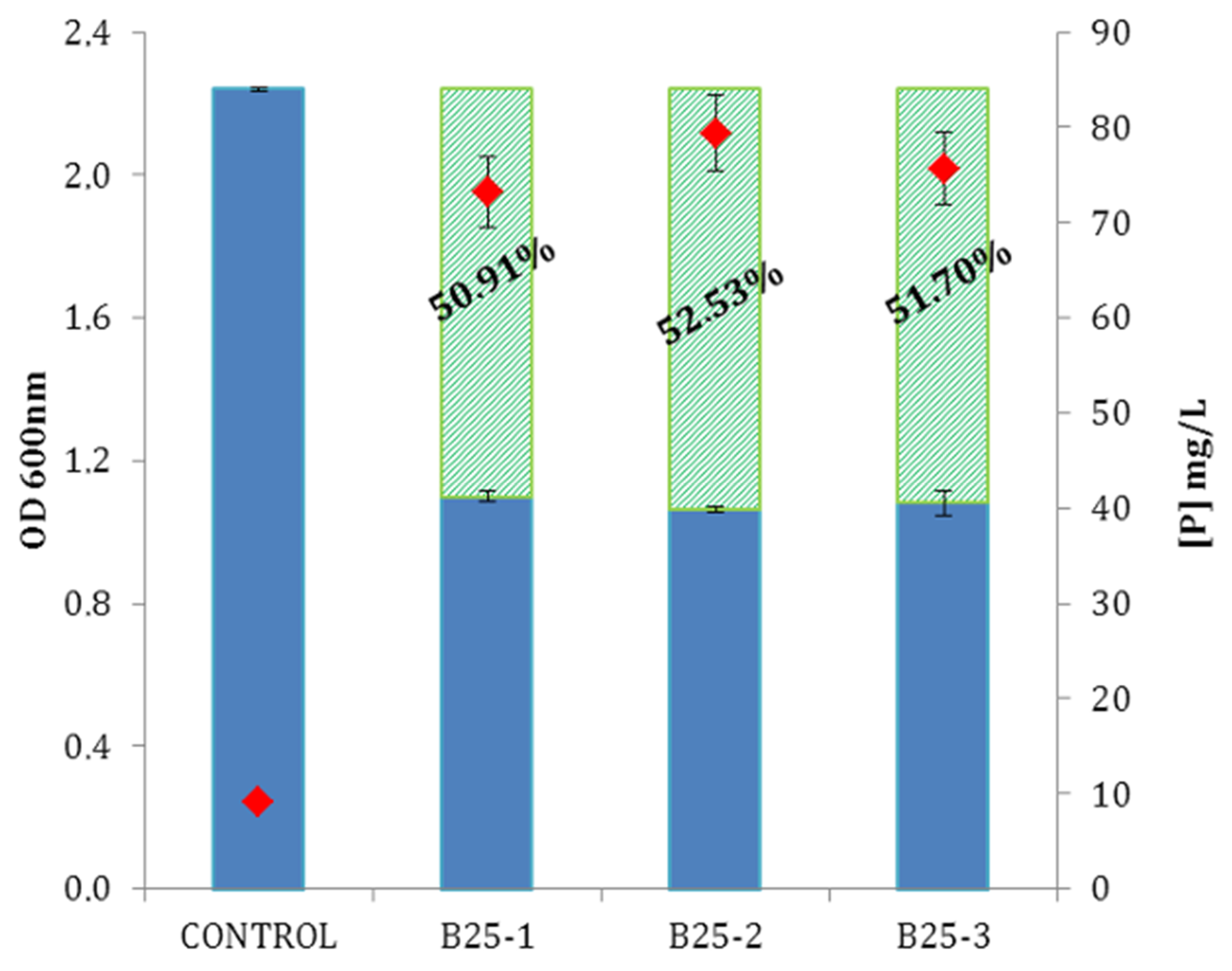
3.3.1. Phosphate Solubilization
3.3.2. Siderophores Detection
3.3.3. IAA Production
3.3.4. Metabolic Profiling on Biolog GenIII Microplates
3.3.5. Minimal Inhibitory Concentrations (MICs) of Antibiotics
3.3.6. UV Sensitivity
3.4. Characterization of the Ecotoxicological Impact of Strain B25
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Approval
References
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borriss, R. Use of plant-associated Bacillus strains as biofertilizers and biocontrol agents in agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–76. ISBN 978-3-642-20331-2. [Google Scholar]
- Fan, B.; Blom, J.; Klenk, H.P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis form an “operational Group B. amyloliquefaciens” within the B. subtilis species complex. Front. Microbiol. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.H. Bacillus velezensis: A valuable member of bioactive molecules within plant microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [Green Version]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.-A.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A review on the Biotechnological Applications of the Operational Group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42-a review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Li, F.; Wang, Z. Characteristics and application of a novel species of Bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef]
- Borriss, R.; Wu, H.; Gao, X. Secondary metabolites of the plant growth-promoting model rhizobacterium Bacillus velezensis FZB42 are involved in direct suppression of plant pathogens and in stimulation of plant-induced systemic resistance. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms. Discovery and Applications; Singh, H.B., Keswani, C., Eds.; Springer: Singapore, 2019; pp. 147–168. [Google Scholar]
- Wang, C.; Zhao, D.; Qi, G.; Mao, Z.; Hu, X.; Du, B.; Liu, K.; Ding, Y. Effects of Bacillus velezensis FKM10 for promoting the growth of Malus hupehensis Rehd and inhibiting Fusarium Verticillioides. Front. Microbiol. 2020, 10, 2889. [Google Scholar] [CrossRef] [PubMed]
- Reva, O.N.; Swanevelder, D.Z.H.; Mwita, L.A.; Mwakilili, A.D.; Muzondiwa, D.; Joubert, M.; Chan, W.Y.; Lutz, S.; Ahrens, C.H.; Avdeeva, L.V.; et al. Genetic, epigenetic and phenotypic diversity of four Bacillus velezensis strains used for plant protection or as probiotics. Front. Microbiol. 2019, 10, 2610. [Google Scholar] [CrossRef] [Green Version]
- Ben Gharsa, H.; Bouri, M.; Mougou Hamdane, A.; Schuster, C.; Leclerque, A.; Rhouma, A. Bacillus velezensis strain MBY2, a potential agent for the management of crown gall disease. PLoS ONE 2021, 16, e0252823. [Google Scholar] [CrossRef]
- Balderas-Ruíz, K.A.; Bustos, P.; Santamaria, R.I.; González, V.; Cristiano-Fajardo, S.A.; Barrera-Ortíz, S.; Mezo-Villalobos, M.; Aranda-Ocampo, S.; Guevara-García, Á.A.; Galindo, E.; et al. Bacillus velezensis 83 a bacterial strain from mango phyllosphere, useful for biological control and plant growth promotion. AMB Express 2020, 10, 163. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, L.; Chen, W.; Dong, Y.; Yang, C.; Li, C.; Xu, H.; Gao, X.; Chen, R.; Li, L.; et al. Isolation and evaluation of Bacillus velezensis ZW-10 as a potential biological control agent against Magnaporthe oryzae. Biotechnol. Biotechnol. Equip. 2020, 34, 714–724. [Google Scholar] [CrossRef]
- Qiao, J.-Q.; Wu, H.-J.; Huo, R.; Gao, X.-W.; Borriss, R. Stimulation of plant growth and biocontrol by Bacillus amyloliquefaciens subsp. plantarum FZB42 engineered for improved action. Chem. Biol. Technol. Agric. 2014, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wu, H.-J.; Qiao, J.; Gao, X.; Borriss, R. Novel routes for improving biocontrol activity of Bacillus based bioinoculants. Front. Microbiol. 2015, 6, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbore, J.; Brutel, A.; Lemainque, A.; Mairey, B.; Médigue, C.; Vallenet, D.; Lefort, F.; Grizard, D. Complete genome sequence of Bacillus velezensis strain B25, a potential Plant Growth-Promoting Rhizobacterium. Genome Announc. 2016, 4, e00058-16. [Google Scholar] [CrossRef] [Green Version]
- Krebs, B.; Höding, B.; Kübart, S.; Workie, M.A.; Junge, H.; Schmiedeknecht, G.; Grosch, R.; Bochow, H.; Hevesi, M. Use of Bacillus subtilis as biocontrol agent. I. Activities and characterization of Bacillus subtilis strains/Anwendung von Bacillus subtilis als Mittel für den biologischen Pflanzenschutz. I. Aktivitäten und Charakterisierung von Bacillus subtilis-Stämmen. Z. Für Pflanzenkrankh. Pflanzenschutz J. Plant Dis. Prot. 1998, 105, 181–197. [Google Scholar]
- Vallenet, D.; Calteau, A.; Cruveiller, S.; Gachet, M.; Lajus, A.; Josso, A.; Mercier, J.; Renaux, A.; Rollin, J.; Rouy, Z.; et al. MicroScope in 2017: An expanding and evolving integrated resource for community expertise of microbial genomes. Nucleic Acids Res. 2017, 45, D517–D528. [Google Scholar] [CrossRef] [PubMed]
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: An integrated platform for the annotation and exploration of microbial gene functions through genomic, pangenomic and metabolic comparative analysis. Nucleic Acids Res. 2020, 48, D579–D589. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A New software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the NCBI AMRFinder tool and resistance gene database using antimicrobial resistance genotype-phenotype correlations in a collection of NARMS isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Yin, Y.; Mao, X.; Yang, J.C.; Chen, X.; Mao, F.; Xu, Y. dbCAN: A web resource for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2012, 40, W445–W451. [Google Scholar] [CrossRef]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma. Trans. Br. Mycol. Soc. 1971, 57, 363-IN2. [Google Scholar] [CrossRef]
- Mehta, S.; Nautiyal, C.S. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr. Microbiol. 2001, 43, 51–56. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.S.; Fernández, F.J. O-CAS, a fast and universal method for siderophore detection. J. Microbiol. Methods 2007, 70, 127–131. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tompkins, T.A.; Barreau, G.; Mathieu, O. Completed Genome Sequence of Bacillus amyloliquefaciens IT45, a Commercial Strain Used in Horticulture. NCBI Genome Database 2013. Available online: https://www.ncbi.nlm.nih.gov/assembly/GCF_000242855.2/ (accessed on 15 July 2021).
- Kunst, F.; Ogasawara, N.; Moszer, I.; Albertini, A.M.; Alloni, G.; Azevedo, V.; Bertero, M.G.; Bessières, P.; Bolotin, A.; Borchert, S.; et al. The complete genome sequence of the gram-positive bacterium Bacillus subtilis. Nature 1997, 390, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, S.D.; Zaretsky, E.J.; Tang, M.; Lopez de Leon, A.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete genome sequence of the industrial bacterium Bacillus licheniformis and comparisons with closely related Bacillus species. Genome Biol. 2004, 5, R77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens ga1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microb. Cell Factories 2009, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, J.; Arora, N.K. Phosphate-solubilizing Bacillus sp. enhances growth, phosphorus uptake and oil yield of Mentha arvensis L. 3 Biotech 2019, 9, 126. [Google Scholar] [CrossRef]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Crop protection against Botrytis cinerea by rhizhosphere biological control agent Bacillus velezensis XT1. Microorganisms 2020, 8, 992. [Google Scholar] [CrossRef]
- Jiang, C.-H.; Liao, M.-J.; Wang, H.-K.; Zheng, M.-Z.; Xu, J.-J.; Guo, J.-H. Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis Cinerea. Biol. Control 2018, 126, 147–157. [Google Scholar] [CrossRef]
- Setlow, P. Resistance of spores of Bacillus species to ultraviolet light. Environ. Mol. Mutagen. 2001, 38, 97–104. [Google Scholar] [CrossRef]
- Albayrak, Ç.B. Bacillus species as biocontrol agents for fungal plant pathogens. In Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Series Bacilli in Climate Resilient Agriculture and Bioprospecting; Islam, M.T., Rahman, M.M., Eds.; Springer International Publishing: New York, NY, USA, 2019; Volume 2, pp. 239–265. ISBN 978-3-030-15175-1. [Google Scholar]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Besset-Manzoni, Y.; Joly, P.; Brutel, A.; Gerin, F.; Soudière, O.; Langin, T.; Prigent-Combaret, C. Does in vitro selection of biocontrol agents guarantee success in planta? A Study case of wheat protection against Fusarium seedling blight by soil bacteria. PLoS ONE 2019, 14, e0225655. [Google Scholar] [CrossRef] [Green Version]
- György, É.; Laslo, É.; Antal, M.; András, C.D. Antibiotic resistance pattern of the allochthonous bacteria isolated from commercially available spices. Food Sci. Nutr. 2021, 9, 4550–4560. [Google Scholar] [CrossRef] [PubMed]
- Irorita Fugaban, J.I.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Bacteriocinogenic Bacillus spp. isolated from Korean fermented cabbage (Kimchi)—Beneficial or hazardous? Fermentation 2021, 7, 56. [Google Scholar] [CrossRef]
- Amichot, M.; Joly, P.; Martin-Laurent, F.; Siaussat, D.; Lavoir, A.-V. Biocontrol, new questions for ecotoxicology? Environ. Sci. Pollut. Res. 2018, 25, 33895–33900. [Google Scholar] [CrossRef] [PubMed]
- Boroski, M.; Martins de Freitas, A.; Sirtori, C. Chapter 10 Daphnia magna in environmental research: A useful tool for ecotoxicological evaluation of conventional and emerging contaminants. In Daphnia: Biology and Mathematics Perspectives; Series: Animal Science, Issues and Research; El-Doma, M.O., Ed.; Nova Science: New York, NY, USA, 2014; pp. 345–434. [Google Scholar]
- Pelosi, C.; Barot, S.; Capowiez, Y.; Hedde, M.; Vandenbulcke, F. Pesticides and earthworms. A review. Agron. Sustain. Dev. 2014, 34, 199–228. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacillus velezensis B25 | Bacillus methylotrophicus FZB42 | Bacillus amyloliquefaciens IT-45 | Bacillus subtilis 168 | Bacillus licheniformis ATCC14580 | |
|---|---|---|---|---|---|
| Genome size (bp) | 3,854,619 | 3,918,589 | 3,928,857 | 4,214,630 | 4,222,336 |
| G+C content (%) | 46.71 | 46.4 | 46.62 | 43.5 | 46.2 |
| Protein-coding sequences | 3683 | 3693 | 3949 | 4106 | 4208 |
| Average CDS size (bp) | 934 | 895 | 888 | 896 | 873 |
| Percentage of coding region | 89 | 88 | 89 | 87 | 86 |
| Ribosomal RNA operons | 7 | 10 | 10 | 10 | 7 |
| Number of tRNAs | 68 | 89 | 95 | 86 | 72 |
| Reference study and GenBank accession number | Gerbore et al. [16] LN999829.1 | Chen et al. [33] CP000560.2 | Tompkins et al. [34] CP004065.1 | Kunst et al. [35] AL009126.3 | Rey et al. [36] CP000002.3 |
| Compound | Enzyme | Gene | High Similarity with | Reference Study |
|---|---|---|---|---|
| Surfactin | NRPS | srfABCD | FZB42 | Chen et al. [33] |
| Fengycin | NRPS | fenABCDE | FZB42 | |
| Bacillibactin | NRPS | dhbABCDEF | FZB42 | |
| Bacilysin | NRPS | bacABCDEFG | FZB42 | |
| Macrolactin | PKS | minABCDEFGHI | FZB42 | |
| Bacillaene | PKS/NRPS | baeBCDE, acpK, baeGHIJLMNRS | FZB42 | |
| Difficidin | PKS | dfnAYXBCDEFGHIJKLM | FZB42 | |
| Mycosubtilin | NRPS | mycABC | IT-45 | Tompkins et al. [34] |
| Amylolysin | RP | amyIEKRAMT | GA1 | Arguelles-Arias [37] |
| B25 Gene Label | Gene Symbol | Product | Antibiotic Class | Similarity with (Amino Acid Identity %) |
|---|---|---|---|---|
| BAMMD1_0254 | lmrB | Lincomycin resistance protein | lincosamide | IT45 (99.8%), FZB42 (99.4%), B168 (88.4%) |
| BAMMD1_0286 | Mdr | multidrug-efflux transporter | aminonucleoside, fluoroquinolone | IT45 (100%), FZB42 (99.6%), B168 (90.2%) |
| BAMMD1_0294 | tmrB | Tunicamycin resistance protein | nucleoside | IT45 (98.5%), FZB42 (98.5%), B168 (77%) |
| BAMMD1_0514 | Cfr | Ribosomal RNA large subunit methyltransferase | oxazolidinone, lincosamide, streptogramin, phenicol | IT45 (99.7%), FZB42 (97.4%) |
| BAMMD1_0534 | vmlR | Ribosome protection protein | streptogramin, oxazolidinone, lincosamide, pleuromutilin, phenicol, tetracycline, macrolide | IT45 (100%), FZB42 (98.3%), B168 (71%) |
| BAMMD1_0665 | aadK | Aminoglycoside 6-adenylyltransferase | aminoglycoside | IT45 (99.3%), B168 (63%) |
| BAMMD1_1112 | Bla | Metallo-beta-lactamase type 2 | Beta-lactam | IT45 (99.6%), FZB42 (97.2%) |
| BAMMD1_1128 | penN | Beta-lactamase | Beta-lactam | IT45 (99.7%), FZB42 (95.7%), B168 (88.5%) |
| BAMMD1_2341 | tetB | Tetracycline resistance protein | tetracycline | IT45 (99.3%), B168 (86.7%) |
| Treatment | FSB Disease Severity | |
|---|---|---|
| Test 1 | Test 2 | |
| Control | 0.9 (± 0.31) ab | 0 (± 0) a |
| Bacillus velezensis B25 | 0 (± 0) a | 0.12 (± 0.12) a |
| Fusarium graminearum B377 | 6.5 (± 1.33) c | 8.38 (± 1.51) b |
| B25 + B377 | 4.41(± 1.1) bc | 4.59 (± 0.96) c |
| Substrates of Biolog Gen III | |||||
|---|---|---|---|---|---|
| Negative Control | − | D-Lactic Acid Methyl Ester | + | l-Aspartic Acid | + |
| Dextrin | + | l-Lactic Acid | + | l-Glutamic Acid | + |
| d-Maltose | + | Citric Acid | + | l-Histidine | + |
| d-trehalose | + | α-Keto-Glutaric Acid | + | l-Pyroglutamic Acid | + |
| d-Cellobiose | + | d-Malic Acid | + | l-Serine | + |
| Gentiobiose | + | l-Malic Acid | + | Pectin | + |
| Sucrose | + | Bromo-Succinic Acid | + | d-Galacturonic Acid | + |
| d-Turanose | + | Tween 40 | + | l-Galactonic Acid Lactone | + |
| Stachyose | + | ϒ-Amino-Butyric Acid | + | Positive control | + |
| d-Raffinose | + | α-Hydroxy-Butyric Acid | − | pH 6 | + |
| α-d-Lactose | + | β-Hydroxy-d, l-Butyric Acid | + | pH 5 | + |
| d-Melibiose | + | α-Keto-Butyric Acid | − | 1% NaCl | + |
| β-Methyl-d-Glucoside | + | Acetoacetic Acid | + | 4% NaCl | + |
| d-Salicin | + | Propionic Acid | + | 8% NaCl | + |
| N-Acetyl-d-Glucosamine | + | Acetic Acid | + | 1% Sodium Lactate | + |
| N-Acetyl-d-Mannosamine | + | Formic Acid | + | Fusidic acid | − |
| N-Acetyl-d-Galactosamine | + | l-Fucose | + | d-Serine-2 | − |
| N-Acetyl Neuraminic Acid | + | l-Rhamnose | + | Troleandomycin | − |
| α-d-Glucose | + | Inosine | + | Rifamycin SV | − |
| d-Mannose | + | d-Sorbitol | + | Minocycline | − |
| d-Fructose | + | d-Mannitol | + | Lincomycin | + |
| d-Galactose | + | d-Arabitol | + | Guanidine HCl | + |
| 3-Methyl Glucose | + | Myo-inositol | + | Niaproof 4 | − |
| d-Fucose | + | Glycerol | + | Vancomycin | − |
| d-Gluconic Acid | + | d-Glucose-6-PO4 | + | Tetrazolium violet | + |
| d-Glucuronic Acid | + | d-Fructose-6-PO4 | + | Tetrazolium Blue | − |
| Glucuronamide | + | d-Aspartic Acid | + | Nalidixic Acid | − |
| Music Acid | + | d-Serine | − | Lithium Chloride | + |
| Quinic Acid | + | Gelatin | + | Potassium Tellurite | + |
| d-Saccharic Acid | + | Glycyl-l-Proline | + | Aztreonam | − |
| p-Hydroxy-Phenylacetic Acid | − | l-Alanine | + | Sodium Butyrate | + |
| Methyl pyruvate | + | l-Arginine | + | Sodium Bromate | − |
| Antibiotics | Imipenem | Amikacin | Ceftazidime | Ciprofloxacin | Vancomycin | Erythromycin | Tetracycline |
|---|---|---|---|---|---|---|---|
| standard range [µg·mL−1] | [32–0.002] | [256–0.015] | [256–0.015] | [32–0.002] | [256–0.015] | [256–0.015] | [256–0.015] |
| MIC for Bacillus B25 [µg·mL−1] | 0.015 | 1 | 32 | 0.03 | 0.5 | 0.12 | 4 |
| Exposure Time under UV-C (Minutes) | |||||||
|---|---|---|---|---|---|---|---|
| Bacterial strains | 0 | 5 | 10 | 15 | 20 | 25 | 30 |
| Bacillus B25 | + | + | + | − | − | − | − |
| Bacillus IT45 | + | − | − | − | − | − | − |
| Product Tested | Bacillus velezensis Strain B25 (1 × 108 cfu·g−1) | |||
|---|---|---|---|---|
| Assay Element/Substrate | Eisenia fetida/Artificial Soil | |||
| Application Rates (kg·ha−1) | Average Mortality at 28 Days (%) * | Biomass Increase (Average %) ** | Fecundity | |
| Average Number per Adult | Effects (%) *** | |||
| Control | 0 | +80.69 | 17.84 | NA |
| 1 | 1.30 (NS) | +87.20 (NS) | 16.93 (NS) | −5.63 (NS) |
| 2 | −3.90 (NS) | +86.84 (NS) | 14.33 (NS) | −20.14 (NS) |
| 10 | −3.90 (NS) | +85.51 (NS) | 17.15 (NS) | −4.39 (NS) |
| NOEC (kg·ha−1) | 10 | |||
| EC50 (kg·ha−1) | >10 | |||
| Endpoint | Concentration (mg·L−1) | |
|---|---|---|
| Immobilization | EC10 (95% confidence limits) | 2853 (Not determined) |
| Immobilization | EC20 (95% confidence limits) | 4484 (Not determined) |
| Immobilization | EC50 (95% confidence limits) | 9712 (Not determined) |
| No Observed Effect Concentration | 4500 | |
| Lowest Observed Effect Concentration | 10,000 | |
| Body length | EC10 (95% confidence limits) | 5963 (4413 to 8058) |
| Body length | EC20 (95% confidence limits) | >10,000 (Not determined) |
| Body length | EC50 (95% confidence limits) | >10,000 (Not determined) |
| No Observed Effect Concentration | 2200 | |
| Lowest Observed Effect Concentration | 4500 | |
| Reproduction | EC10 (95% confidence limits) | 4240 (Not determined) |
| Reproduction | EC20 (95% confidence limits) | 5062 (Not determined) |
| Reproduction | EC50 (95% confidence limits) | 6855 (Not determined) |
| No Observed Effect Concentration | 4500 | |
| Lowest Observed Effect Concentration | 10,000 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joly, P.; Calteau, A.; Wauquier, A.; Dumas, R.; Beuvin, M.; Vallenet, D.; Crovadore, J.; Cochard, B.; Lefort, F.; Berthon, J.-Y. From Strain Characterization to Field Authorization: Highlights on Bacillus velezensis Strain B25 Beneficial Properties for Plants and Its Activities on Phytopathogenic Fungi. Microorganisms 2021, 9, 1924. https://doi.org/10.3390/microorganisms9091924
Joly P, Calteau A, Wauquier A, Dumas R, Beuvin M, Vallenet D, Crovadore J, Cochard B, Lefort F, Berthon J-Y. From Strain Characterization to Field Authorization: Highlights on Bacillus velezensis Strain B25 Beneficial Properties for Plants and Its Activities on Phytopathogenic Fungi. Microorganisms. 2021; 9(9):1924. https://doi.org/10.3390/microorganisms9091924
Chicago/Turabian StyleJoly, Pierre, Alexandra Calteau, Aurélie Wauquier, Rémi Dumas, Mylène Beuvin, David Vallenet, Julien Crovadore, Bastien Cochard, François Lefort, and Jean-Yves Berthon. 2021. "From Strain Characterization to Field Authorization: Highlights on Bacillus velezensis Strain B25 Beneficial Properties for Plants and Its Activities on Phytopathogenic Fungi" Microorganisms 9, no. 9: 1924. https://doi.org/10.3390/microorganisms9091924
APA StyleJoly, P., Calteau, A., Wauquier, A., Dumas, R., Beuvin, M., Vallenet, D., Crovadore, J., Cochard, B., Lefort, F., & Berthon, J.-Y. (2021). From Strain Characterization to Field Authorization: Highlights on Bacillus velezensis Strain B25 Beneficial Properties for Plants and Its Activities on Phytopathogenic Fungi. Microorganisms, 9(9), 1924. https://doi.org/10.3390/microorganisms9091924