
Analysis of Human Gut Microbiota Composition Associated to the Presence of Commensal and Pathogen Microorganisms in Côte d’Ivoire

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Sample Management and Fecal Nucleic Acids Extraction
2.3. xTAG GPP Assay
2.4. Detection of Adeno-, Entero-, and Parechoviruses by Real-Time PCR
2.5. Blastocystis and Commensal Entamoeba spp. Detection and Identification
2.6. 16S rRNA Gene Amplicon Sequencing
2.7. 16S rRNA Amplicon Data Processing
3. Results
3.1. Study Area and Host Population
3.2. Viral, Bacterial, and Protozoa Detection
3.3. Gut Microbiota Composition
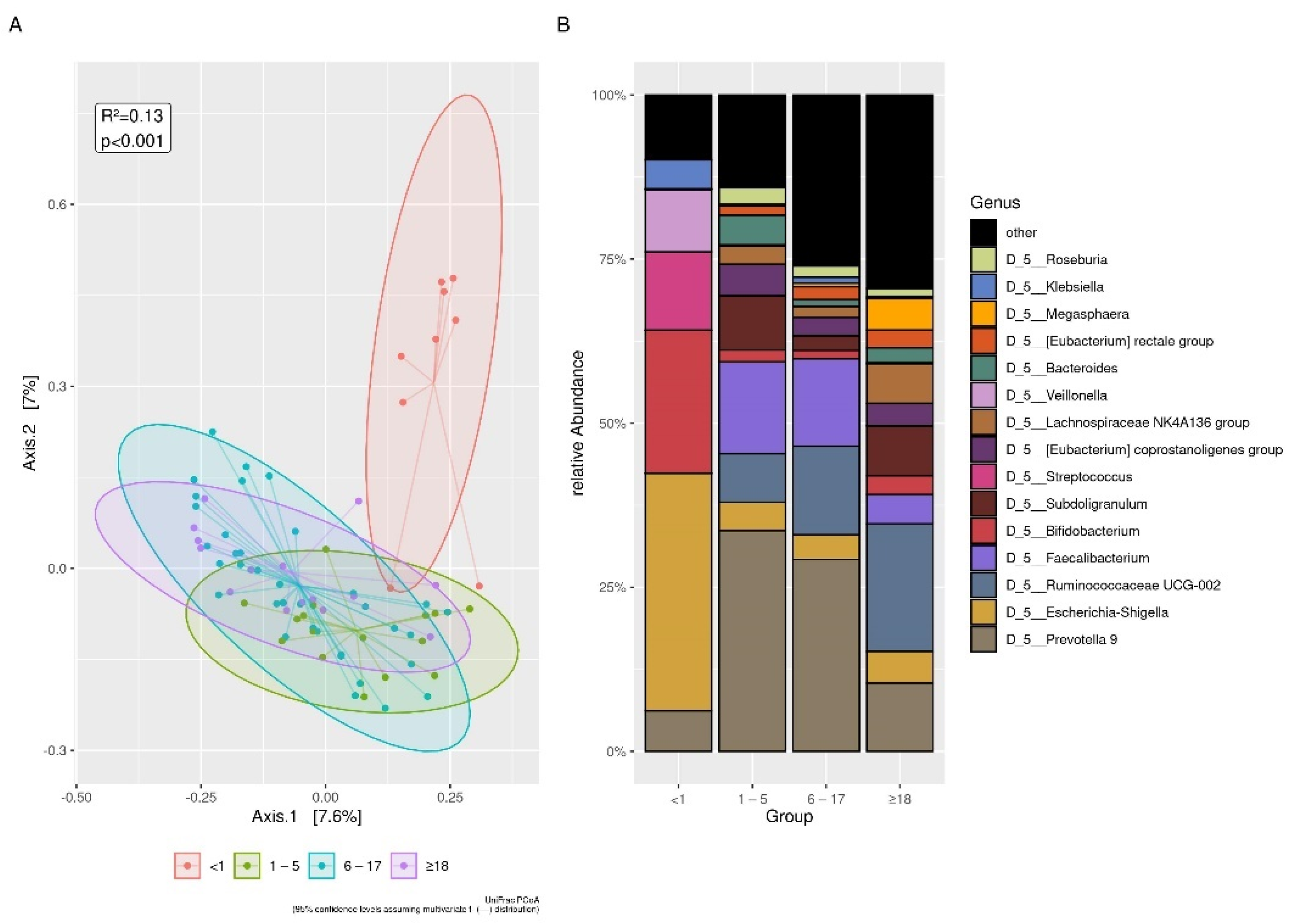
3.3.1. Age
3.3.2. Denovo Clustering
3.3.3. Sex
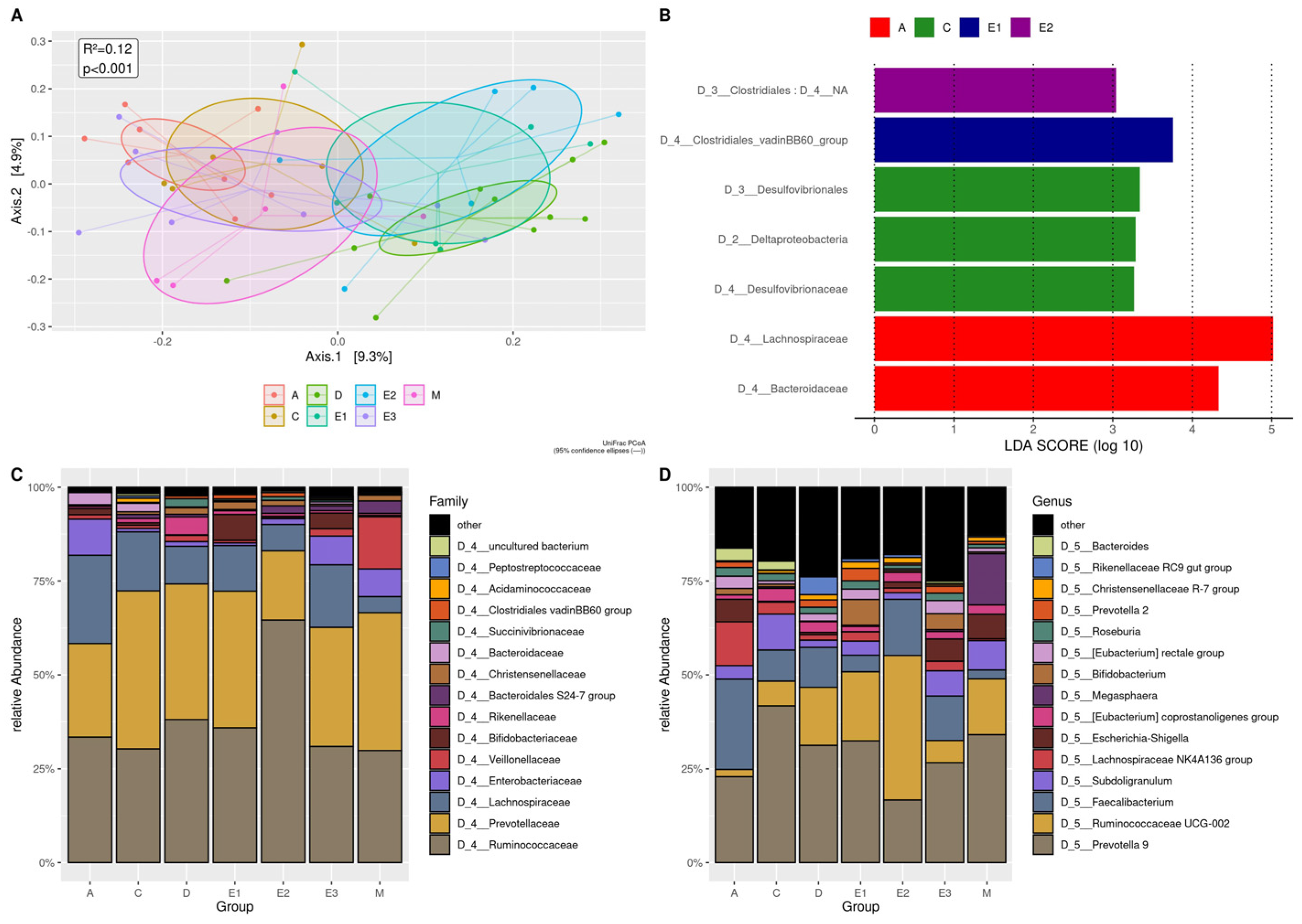
3.3.4. Localities and Seasonality
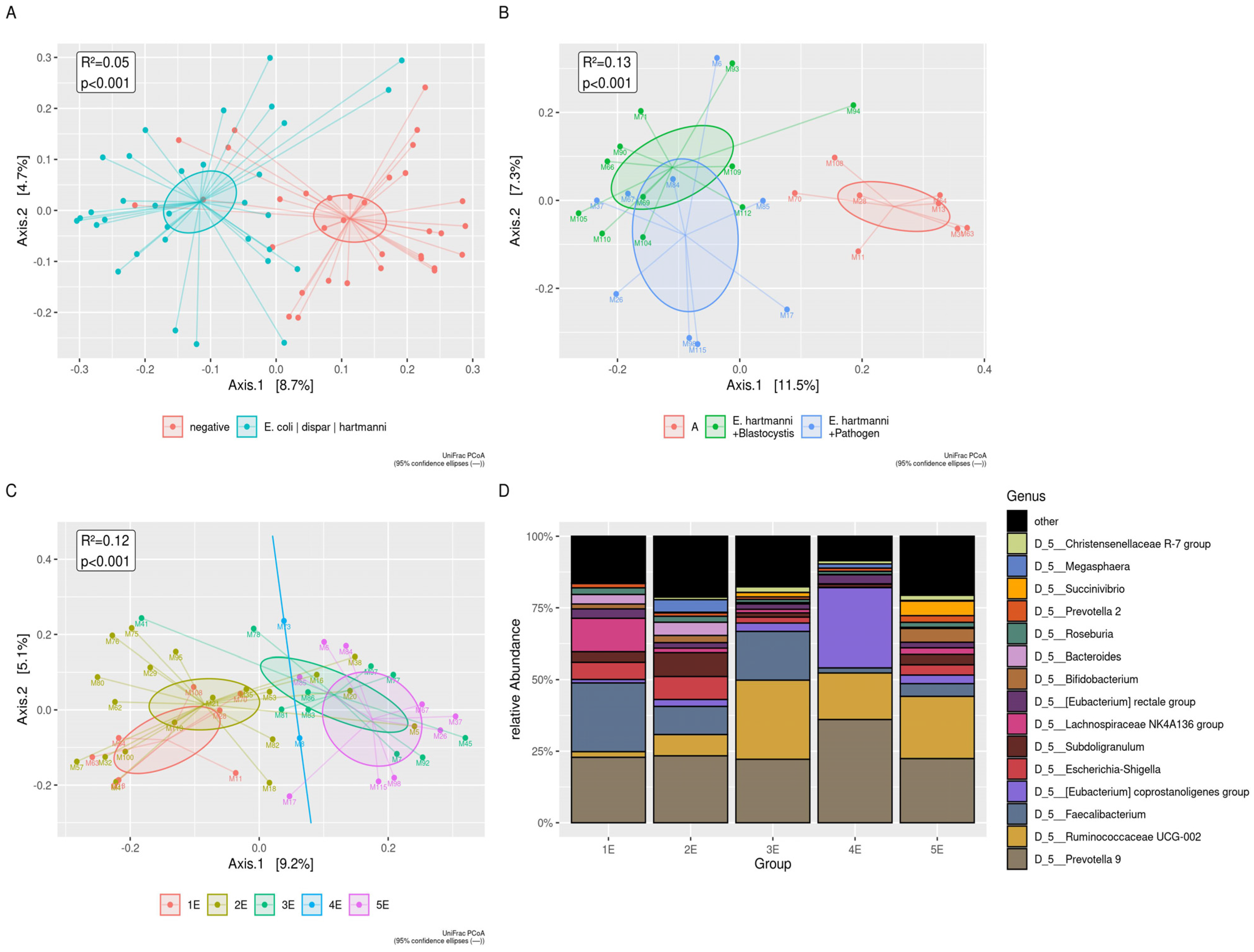
3.4. Blastocystis, Commensal Entamoeba spp., and Intestinal Pathogens
3.5. Blastocystis
3.6. Commensal Entamoeba spp.
3.7. G. duodenalis and Other Intestinal Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vonaesch, P.; Anderson, M.; Sansonetti, P.J. Pathogens, microbiome and the host: Emergence of the ecological Koch’s postulates. FEMS Microbiol. Rev. 2018, 42, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. The gut as communicator between environment and host: Immunological consequences. Eur. J. Pharmacol. 2011, 668 (Suppl. 1), S16–S32. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Gordon, J.I. The core gut microbiome, energy balance and obesity. J. Physiol. 2009, 587 Pt 17, 4153–4158. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Holmes, E.; Li, J.V.; Marchesi, J.R.; Nicholson, J.K. Gut microbiota composition and activity in relation to host metabolic phenotype and disease risk. Cell Metab. 2012, 16, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Collado, M.C.; Mangani, C.; Maleta, K.; Laitinen, K.; Ashorn, P.; Isolauri, E.; Salminen, S. Distinct gut microbiota in southeastern African and northern European infants. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 812–816. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; DeLany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 2013, 98, 111–120. [Google Scholar] [CrossRef]
- Brooks, A.W.; Priya, S.; Blekhman, R.; Bordenstein, S.R. Gut microbiota diversity across ethnicities in the United States. PLoS Biol. 2018, 16, e2006842. [Google Scholar] [CrossRef]
- Ayeni, F.A.; Biagi, E.; Rampelli, S.; Fiori, J.; Soverini, M.; Audu, H.J.; Cristino, S.; Caporali, L.; Schnorr, S.L.; Carelli, V.; et al. Infant and Adult Gut Microbiome and Metabolome in Rural Bassa and Urban Settlers from Nigeria. Cell Rep. 2018, 23, 3056–3067. [Google Scholar] [CrossRef] [PubMed]
- Samb-Ba, B.; Mazenot, C.; Gassama-Sow, A.; Dubourg, G.; Richet, H.; Hugon, P.; Lagier, J.C.; Raoult, D.; Fenollar, F. MALDI-TOF identification of the human Gut microbiome in people with and without diarrhea in Senegal. PLoS ONE 2014, 9, e87419. [Google Scholar] [CrossRef]
- De Filippo, C.; Di Paola, M.; Ramazzotti, M.; Albanese, D.; Pieraccini, G.; Banci, E.; Miglietta, F.; Cavalieri, D.; Lionetti, P. Diet, Environments, and Gut Microbiota. A Preliminary Investigation in Children Living in Rural and Urban Burkina Faso and Italy. Front. Microbiol. 2017, 8, 1979. [Google Scholar] [CrossRef] [PubMed]
- Afolayan, A.O.; Ayeni, F.A.; Moissl-Eichinger, C.; Gorkiewicz, G.; Halwachs, B.; Högenauer, C. Impact of a Nomadic Pastoral Lifestyle on the Gut Microbiome in the Fulani Living in Nigeria. Front. Microbiol. 2019, 10, 2138. [Google Scholar] [CrossRef]
- Kuss, S.K.; Best, G.T.; Etheredge, C.A.; Pruijssers, A.J.; Frierson, J.M.; Hooper, L.V.; Dermody, T.S.; Pfeiffer, J.K. Intestinal microbiota promote enteric virus replication and systemic pathogenesis. Science 2011, 334, 249–252. [Google Scholar] [CrossRef]
- Iebba, V.; Santangelo, F.; Totino, V.; Pantanella, F.; Monsia, A.; Di Cristanziano, V.; Di Cave, D.; Schippa, S.; Berrilli, F.; D’Alfonso, R. Gut microbiota related to Giardia duodenalis, Entamoeba spp. and Blastocystis hominis infections in humans from Côte d’Ivoire. J. Infect. Dev. Ctries 2016, 10, 1035–1041. [Google Scholar] [CrossRef]
- Lukeš, J.; Stensvold, C.R.; Jirků-Pomajbíková, K.; Wegener Parfrey, L. Are Human Intestinal Eukaryotes Beneficial or Commensals? PLoS Pathog. 2015, 11, e1005039. [Google Scholar] [CrossRef] [PubMed]
- Popovic, A.; Bourdon, C.; Wang, P.W.; Guttman, D.S.; Voskuijl, W.; Grigg, M.E.; Bandsma, R.H.J.; Parkinson, J. Design and application of a novel two-amplicon approach for defining eukaryotic microbiota. Microbiome 2018, 6, 228. [Google Scholar] [CrossRef]
- Morton, E.R.; Lynch, J.; Froment, A.; Lafosse, S.; Heyer, E.; Przeworski, M.; Blekhman, R.; Ségurel, L. Variation in Rural African Gut Microbiota Is Strongly Correlated with Colonization by Entamoeba and Subsistence. PLoS Genet. 2015, 11, e1005658. [Google Scholar] [CrossRef] [PubMed]
- Audebert, C.; Even, G.; Cian, A.; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef]
- Gilchrist, C.A.; Petri, S.E.; Schneider, B.N.; Reichman, D.J.; Jiang, N.; Begum, S.; Watanabe, K.; Jansen, C.S.; Elliott, K.P.; Burgess, S.L.; et al. Role of the Gut Microbiota of Children in Diarrhea Due to the Protozoan Parasite Entamoeba histolytica. J. Infect. Dis. 2016, 213, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Ngure, F.M.; Reid, B.M.; Humphrey, J.H.; Mbuya, M.N.; Pelto, G.; Stoltzfus, R.J. Water, sanitation, and hygiene (WASH), environmental enteropathy, nutrition, and early child development: Making the links. Ann. N. Y. Acad. Sci. 2014, 1308, 118–128. [Google Scholar] [CrossRef]
- Di Cristanziano, V.; Timmen-Wego, M.; Lübke, N.; Kaiser, R.; Pfister, H.; Di Cave, D.; Berrilli, F.; Kaboré, Y.; D’Alfonso, R. Application of Luminex Gastrointestinal Pathogen Panel to human stool samples from Côte d’Ivoire. J. Infect. Dev. Ctries 2015, 9, 884–889. [Google Scholar] [CrossRef]
- Cristanziano, V.D.; Bottcher, S.; Diedrich, S.; Timmen-Wego, M.; Knops, E.; Lübke, N.; Kaiser, R.; Pfister, H.; Kaboré, Y.; D’Alfonso, R. Detection and characterization of enteroviruses and parechoviruses in healthy people living in the South of Cote d’Ivoire. J. Clin. Virol. 2015, 71, 40–43. [Google Scholar] [CrossRef]
- Souppart, L.; Sanciu, G.; Cian, A.; Wawrzyniak, I.; Delbac, F.; Capron, M.; Dei-Cas, E.; Boorom, K.; Delhaes, L.; Viscogliosi, E. Molecular epidemiology of human Blastocystis isolates in France. Parasitol. Res. 2009, 105, 413–421. [Google Scholar] [CrossRef] [PubMed]
- D’Alfonso, R.; Santoro, M.; Essi, D.; Monsia, A.; Kaboré, Y.; Glé, C.; Di Cave, D.; Sorge, R.P.; Di Cristanziano, V.; Berrilli, F. Blastocystis in Côte d’Ivoire: Molecular identification and epidemiological data. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2243–2250. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.L.; Bandea, R.; Martins, L.A.; de Macedo, H.W.; Peralta, R.H.; Peralta, J.M.; Ndubuisi, M.I.; da Silva, A.J. Differential identification of Entamoeba spp. based on the analysis of 18S rRNA. Parasitol. Res. 2010, 106, 883–888. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Bertrand, T.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- RC Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 10 February 2015).
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Brewster, R.; Tamburini, F.B.; Asiimwe, E.; Oduaran, O.; Hazelhurst, S.; Bhatt, A.S. Surveying Gut Microbiome Research in Africans: Toward Improved Diversity and Representation. Trends Microbiol. 2019, 27, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Korpe, P.S.; Petri, W.A. Environmental enteropathy: Critical implications of a poorly understood condition. Trends Mol. Med. 2012, 18, 328–336. [Google Scholar] [CrossRef]
- Stecher, B. The Roles of Inflammation, Nutrient Availability and the Commensal Microbiota in Enteric Pathogen Infection. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Gorvitovskaia, A.; Holmes, S.P.; Huse, S.M. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Huse, S.M.; Ye, Y.; Zhou, Y.; Fodor, A.A. A core human microbiome as viewed through 16S rRNA sequence clusters. PLoS ONE 2012, 7, e34242. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Ning, K. Stereotypes about Enterotype: The Old and New Ideas. Genom. Proteom. Bioinform. 2019, 17, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.V.; de Roode, J.C.; Gerardo, N.M. Diet-microbiome-disease: Investigating diet’s influence on infectious disease resistance through alteration of the gut microbiome. PLoS Pathog. 2019, 15, e1007891. [Google Scholar] [CrossRef]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed]
- Labbé, A.; Ganopolsky, J.G.; Martoni, C.J.; Prakash, S.; Jones, M.L. Bacterial bile metabolising gene abundance in Crohn’s, ulcerative colitis and type 2 diabetes metagenomes. PLoS ONE 2014, 9, e115175. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Ferrer, M. Functional Redundancy-Induced Stability of Gut Microbiota Subjected to Disturbance. Trends Microbiol. 2016, 24, 402–413. [Google Scholar] [CrossRef]
- Livanos, A.E.; Snider, E.J.; Whittier, S.; Chong, D.H.; Wang, T.C.; Abrams, J.A.; Freedberg, D.E. Rapid gastrointestinal loss of Clostridial Clusters IV and XIVa in the ICU associates with an expansion of gut pathogens. PLoS ONE 2018, 13, e0200322. [Google Scholar]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. 2), S1–S63. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef]
- Jalanka, J.; Major, G.; Murray, K.; Singh, G.; Nowak, A.; Kurtz, C.; Silos-Santiago, I.; Johnston, J.M.; de Vos, W.M.; Spiller, R. The Effect of Psyllium Husk on Intestinal Microbiota in Constipated Patients and Healthy Controls. Int. J. Mol. Sci. 2019, 20, 433. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Kelley, S.T.; Chen, Y.; Escobar, J.S.; Mueller, N.T.; Ley, R.E.; McDonald, D.; Huang, S.; Swafford, A.D.; Knight, R.; et al. Age- and Sex-Dependent Patterns of Gut Microbial Diversity in Human Adults. mSystems 2019, 4, e00261-19. [Google Scholar] [CrossRef]
- Takagi, T.; Naito, Y.; Inoue, R.; Kashiwagi, S.; Uchiyama, K.; Mizushima, K.; Tsuchiya, S.; Dohi, O.; Yoshida, N.; Kamada, K.; et al. Differences in gut microbiota associated with age, sex, and stool consistency in healthy Japanese subjects. J. Gastroenterol. 2019, 54, 53–63. [Google Scholar] [CrossRef]
- Ouattara, M.; N’guéssan, N.A.; Yapi, A.; N’goran, E.K. Prevalence and spatial distribution of Entamoeba histolytica/dispar and Giardia lamblia among schoolchildren in Agboville area (Côte d’Ivoire). PLoS Negl. Trop. Dis. 2010, 4, e574. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- El Safadi, D.; Gaayeb, L.; Meloni, D.; Cian, A.; Poirier, P.; Wawrzyniak, I.; Delbac, F.; Dabboussi, F.; Delhaes, L.; Seck, M.; et al. Children of Senegal River Basin show the highest prevalence of Blastocystis sp. ever observed worldwide. BMC Infect. Dis. 2014, 14, 164. [Google Scholar] [CrossRef]
- Lokmer, A.; Cian, A.; Froment, A.; Gantois, N.; Viscogliosi, E.; Chabé, M.; Ségurel, L. Use of shotgun metagenomics for the identification of protozoa in the gut microbiota of healthy individuals from worldwide populations with various industrialization levels. PLoS ONE 2019, 14, e0211139. [Google Scholar] [CrossRef] [PubMed]
- Berrilli, F.; D’Alfonso, R.; Giangaspero, A.; Marangi, M.; Brandonisio, O.; Kaboré, Y.; Glé, C.; Cianfanelli, C.; Lauro, R.; Di Cave, D. Giardia duodenalis genotypes and Cryptosporidium species in humans and domestic animals in Cote d’Ivoire: Occurrence and evidence for environmental contamination. Trans. R Soc. Trop. Med. Hyg. 2012, 106, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Annu. Rev. Immunol. 2012, 30, 759–795. [Google Scholar] [CrossRef] [PubMed]
- Muhsen, K.; Levine, M.M. A systematic review and meta-analysis of the association between Giardia lamblia and endemic pediatric diarrhea in developing countries. Clin. Infect. Dis. 2012, 55 (Suppl. 4), S271–S293. [Google Scholar] [CrossRef]
- Berrilli, F.; Di Cave, D.; N’Guessan, R.; Kaboré, Y.; Giangaspero, A.; Sorge, R.P.; D’Alfonso, R. Social determinants associated with Giardia duodenalis infection in southern Côte d’Ivoire. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1799–1802. [Google Scholar] [CrossRef]
- Graziani, C.; Talocco, C.; De Sire, R.; Petito, V.; Lopetuso, L.R.; Gervasoni, J.; Persichilli, S.; Franceschi, F.; Ojetti, V.; Gasbarrini, A.; et al. Intestinal permeability in physiological and pathological conditions: Major determinants and assessment modalities. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 795–810. [Google Scholar] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.L.; Chan, C.C.; Chen, H.P.; Fung, C.P.; Lin, C.P.; Chan, W.L.; Liu, C.Y. Clinical characteristics and endoscopic findings associated with Blastocystis hominis in healthy adults. Am. J. Trop. Med. Hyg. 2003, 69, 213–216. [Google Scholar] [CrossRef][Green Version]
- Scanlan, P.D.; Stensvold, C.R.; Rajilić-Stojanović, M.; Heilig, H.G.; De Vos, W.M.; O’Toole, P.W.; Cotter, P.D. The microbial eukaryote Blastocystis is a prevalent and diverse member of the healthy human gut microbiota. FEMS Microbiol. Ecol. 2014, 90, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.O.; Bonde, I.; Nielsen, H.B.; Stensvold, C.R. A retrospective metagenomics approach to studying Blastocystis. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Ramírez, M.E.; Partida-Rodríguez, O.; Laforest-Lapointe, I.; Reynolds, L.A.; Brown, E.M.; Valdez-Salazar, A.; Morán-Silva, P.; Rojas-Velázquez, L.; Morien, E.; Parfrey, L.W.; et al. Asymptomatic Intestinal Colonization with Protist Blastocystis is Strongly Associated with Distinct Microbiome Ecological Patterns. mSystems 2018, 3, e00007-18. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Gérard, P. Metabolism of cholesterol and bile acids by the gut microbiota. Pathogens 2013, 3, 14–24. [Google Scholar] [CrossRef]
- Basson, A.; Trotter, A.; Rodriguez-Palacios, A.; Cominelli, F. Mucosal Interactions between Genetics, Diet, and Microbiome in Inflammatory Bowel Disease. Front. Immunol. 2016, 7, 290. [Google Scholar] [CrossRef]
- Rampelli, S.; Candela, M.; Turroni, S.; Biagi, E.; Pflueger, M.; Wolters, M.; Ahrens, W.; Brigidi, P. Microbiota and lifestyle interactions through the lifespan. Trends Food Sci. Technol. 2016, 57, 265–272. [Google Scholar] [CrossRef]
- Tito, R.Y.; Chaffron, S.; Caenepeel, C.; Lima-Mendez, G.; Wang, J.; Vieira-Silva, S.; Falony, G.; Hildebrand, F.; Darzi, Y.; Rymenans, L.; et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut 2019, 68, 1180–1189. [Google Scholar] [CrossRef]
- Faith, D.P. Phylogenetic Diversity and Conservation Evaluation: Perspectives on Multiple Values, Indices, and Scales of Application. In Phylogenetic Diversity: Applications and Challenges in Biodiversity Science; Scherson, R.A., Faith, D.P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–26. [Google Scholar]
- Tsolis, R.M.; Bäumler, A.J. Gastrointestinal host-pathogen interaction in the age of microbiome research. Curr. Opin. Microbiol. 2020, 53, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Ordiz, M.I.; Stephenson, K.; Agapova, S.; Wylie, K.M.; Maleta, K.; Martin, J.; Trehan, I.; Tarr, P.I.; Manary, M.J. Environmental Enteric Dysfunction and the Fecal Microbiota in Malawian Children. Am. J. Trop. Med. Hyg. 2017, 96, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.R.; Davenport, E.R.; Gautam, Y.; Bhandari, D.; Tandukar, S.; Ng, K.M.; Fragiadakis, G.K.; Holmes, S.; Gautam, G.P.; Leach, J.; et al. Gut microbiome transition across a lifestyle gradient in Himalaya. PLoS Biol. 2018, 16, e2005396. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, R.; Janardhan, H.P.; George, S.; Raghava, M.V.; Muliyil, J.; Ramakrishna, B.S. Molecular studies of fecal anaerobic commensal bacteria in acute diarrhea in children. J. Pediatr. Gastroenterol. Nutr. 2008, 46, 514–519. [Google Scholar] [CrossRef]
- Ubeda, C.; Djukovic, A.; Isaac, S. Roles of the intestinal microbiota in pathogen protection. Clin. Transl. Immunol. 2017, 6, e128. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Ramanan, V.; Korolev, K.S. Interactions between species introduce spurious associations in microbiome studies. PLoS Comput. Biol. 2018, 14, e1005939. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Díaz, C.; García-Orozco, A.; Riera-Leal, A.; Padilla-Arellano, J.R.; Fafutis-Morris, M. Microbiota and Its Role on Viral Evasion: Is It With Us or Against Us? Front. Cell Infect. Microbiol. 2019, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Shetty, S.A.; Marathe, N.P.; Lanjekar, V.; Ranade, D.; Shouche, Y.S. Comparative genome analysis of Megasphaera sp. reveals niche specialization and its potential role in the human gut. PLoS ONE 2013, 8, e79353. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Chang, Y.; Kim, H.N.; Ryu, S.; Kwon, M.J.; Cho, Y.K.; Kim, H.L.; Cheong, H.S.; Joo, E.J. Alterations of the Gut Microbiome in Chronic Hepatitis B Virus Infection Associated with Alanine Aminotransferase Level. J. Clin. Med. 2019, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Chabé, M.; Lokmer, A.; Ségurel, L. Gut Protozoa: Friends or Foes of the Human Gut Microbiota? Trends Parasitol. 2017, 33, 925–934. [Google Scholar] [CrossRef]
- Marie, C.; Petri, W.A. Regulation of virulence of Entamoeba histolytica. Annu. Rev. Microbiol. 2014, 68, 493–520. [Google Scholar] [CrossRef]
- Wilson, I.W.; Weedall, G.D.; Hall, N. Host-Parasite interactions in Entamoeba histolytica and Entamoeba dispar: What have we learned from their genomes? Parasite Immunol. 2012, 34, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Luna, J.; Piña-Vázquez, C.; Reyes-López, M.; Ortiz-Estrada, G.; de la Garza, M. Proteases from Entamoeba spp. and Pathogenic Free-Living Amoebae as Virulence Factors. J. Trop. Med. 2013, 2013, 890603. [Google Scholar] [CrossRef] [PubMed]
- Leon-Coria, A.; Kumar, M.; Chadee, K. The delicate balance between. Gut Microbes 2020, 11, 118–125. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Groups | <1 year | >1–5 years | 6–17 years | >18 years | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Localities | n F/M | Median min–max | n F/M | Median min–max | n F/M | Median min–max | n F/M | Median min–max | n F/M | Median min–max | |
| Bonoua | 8 2/6 | 0.1 0.02–0.5 | 10 6/4 | 4.5 2–5 | 20 10/10 | 11.5 8–17 | 15 7/8 | 30 18–59 | 53 25/28 | 11 0.02–59 | |
| Assouindé | 1 1/0 | 0.75 | - | - | 3 2/1 | 11 11–15 | - | - | 4 3/1 | 11 0.75–15 | |
| Kimokro | - | - | 1 0/1 | 2.5 | 5 2/3 | 8 6–10 | - | - | 6 2/4 | 7.5 2.5–10 | |
| Yaou | - | - | 5 3/2 | 5 4–5 | 8 3/5 | 6.5 6–13 | - | - | 13 6/7 | 6 4–13 | |
| Total | 9 3/6 | 0.1 0.1–0.7 | 16 9/7 | 4.5 1.8–5 | 36 17/19 | 10 6–17 | 15 7/8 | 0.1 0.1–05 | 76 36/40 | 9 0.02–59 | |
| Viruses and Bacteria | Positive Samples n (F/M) | Mean Age (min–max) | Detected Pathogens (n) |
|---|---|---|---|
| Adenovirus (AdV) | 17 (9/8) | 13.5 (0.25–59) | EV (2); NoV (2); ETEC (3); Campylobacter (1) |
| Parechovirus (PeV) | 4 (1/3) | 5.4 (0.1–14) | EV (2); NoV (1); Campylobacter (1) |
| Enterovirus (EV) | 14 (5/9) | 6.5 (0.1–14) | PeV (2); AdV (3); NoV(3); Campylobacter (2); ETEC (4) |
| Norovirus G1/G2 (NoV) | 8 (3/5) | 12.7 (0.1–39) | EV (3); AdV (2); PeV (1); RoV (1) |
| Rotavirus A (RoV) | 1 (1/0) | 39 | RoV (1) |
| Campylobacter | 5 (3/2) | 6.3 (9.75–9) | E.coli O157 (1); ETEC (1); STEC (1); Shigella (1); EV (2); PeV (1); AdV (1) |
| E.coli O157 | 4 (2/2) | 11 (17–8) | ETEC (3); STEC (1); Campylobacter (1); Shigella (1); EV (2); PeV (1); AdV (1) |
| ETEC | 10 (6/4) | 7 (1.8–17) | E.coli O157 (3); STEC (2); Campylobacter (3); Shigella (2); EV (4); AdV (3) |
| Salmonella | 1 (0/1) | 43 | - |
| STEC | 2 (1/1) | 9 (9) | E.coli O157 (1); ETEC (2); Campylobacter (1) |
| Shigella | 4 (3/1) | 11.7 (0.75–24) | E.coli O157 (1); ETEC (2); Campylobacter (1); PeV (1) |
| E. hartmanni | En. coli | E. dispar | Blastocystis | G. duodenalis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Localities | Sex | Age | Sex | Age | Sex | Age | Sex | Age | Sex | Age |
| n (F/M) | Mean (min–max) | n (F/M) | Mean (min–max) | n (F/M) | Mean (min–max) | n (F/M) | Mean (min–max) | n (F/M) | Mean (min–max) | |
| Bonoua | 16 (6/10) | 21 (9–50) | 2 (2F) | 3 (2–4) | 1 (1F) | 5 | 28 (17/11) | 22 (9–59) | 8 * (6/2) | 17 (2–50) |
| Assouindé | 1 (1F) | 11 | - | - | - | - | 3 (2/1) | 12 (11–15) | 3 (2/1) | 12 (11–15) |
| Kimoukro | 1 (1M) | 6 | 3 (1/2) | 7 (2–10) | - | - | 4 (2/2) | 8 (6–10) | 7 (2–10) | 8 (4/4) |
| Yaou | 2 (2F) | 5 (4–6) | 6 (4/2) | 6 (5–7) | 1 (1M) | 5 | 12 (6/6) | 6 (4–13) | 8 (4/4) | 5 (4–7) |
| Total | 20 (9/11) | 18 (4–50) | 11 (7/4) | 12 (2–59) | 2 (1/1) | 5 | 47 (27/20) | 14 (4–59) | 24 (14/10) | 11 (2–50) |
| Age Groups (n) | <1 year (9) Median (min–max) | 1–5 years (16) Median (min–max) | 6–17 years (36) Median (min–max) | >18 years (15) Median (min–max) | |
|---|---|---|---|---|---|
| Alpha Diversity | |||||
| Shannon index | 1.703 (0.6059–3.019) | 3.411 (2.2730–4.131) c | 3.703 (2.1940–4.962) a | 3.631 (1.5140–4.589) c | |
| Observed OTUs | 39.00 (19.00–86.00) | 151.00 (79.00–258.00) b | 189.00 (94.00–459.00) a | 177.00 (82.00–305.00) a | |
| Faith’s phylogenetic diversity | 4.601 (0.9572–12.04) | 12.850 (7.2710–20.82) c | 16.560 (7.3410–34.15) a | 17.540 (8.4570–25.67) a | |
| Chao1 index | 39.50 (19.00–95.07) | 189.50 (98.60–313.30) b | 228.20 (105.50–610.00) a | 231.00 (100.40–406.90) b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Cristanziano, V.; Farowski, F.; Berrilli, F.; Santoro, M.; Di Cave, D.; Glé, C.; Daeumer, M.; Thielen, A.; Wirtz, M.; Kaiser, R.; et al. Analysis of Human Gut Microbiota Composition Associated to the Presence of Commensal and Pathogen Microorganisms in Côte d’Ivoire. Microorganisms 2021, 9, 1763. https://doi.org/10.3390/microorganisms9081763
Di Cristanziano V, Farowski F, Berrilli F, Santoro M, Di Cave D, Glé C, Daeumer M, Thielen A, Wirtz M, Kaiser R, et al. Analysis of Human Gut Microbiota Composition Associated to the Presence of Commensal and Pathogen Microorganisms in Côte d’Ivoire. Microorganisms. 2021; 9(8):1763. https://doi.org/10.3390/microorganisms9081763
Chicago/Turabian StyleDi Cristanziano, Veronica, Fedja Farowski, Federica Berrilli, Maristella Santoro, David Di Cave, Christophe Glé, Martin Daeumer, Alexander Thielen, Maike Wirtz, Rolf Kaiser, and et al. 2021. "Analysis of Human Gut Microbiota Composition Associated to the Presence of Commensal and Pathogen Microorganisms in Côte d’Ivoire" Microorganisms 9, no. 8: 1763. https://doi.org/10.3390/microorganisms9081763
APA StyleDi Cristanziano, V., Farowski, F., Berrilli, F., Santoro, M., Di Cave, D., Glé, C., Daeumer, M., Thielen, A., Wirtz, M., Kaiser, R., Eberhardt, K. A., Vehreschild, M. J. G. T., & D’Alfonso, R. (2021). Analysis of Human Gut Microbiota Composition Associated to the Presence of Commensal and Pathogen Microorganisms in Côte d’Ivoire. Microorganisms, 9(8), 1763. https://doi.org/10.3390/microorganisms9081763