
Improvement of CO2 and Acetate Coupling into Lactic Acid by Genetic Manipulation of the Hyperthermophilic Bacterium Thermotoga neapolitana

 , ,
, ,  ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Gas and Chemical Analysis
2.3. Construction of Vectors
2.4. Spheroplast Formation and Transformation
3. Results and Discussion
3.1. Assessment of Genetic Tools and Transformation Method
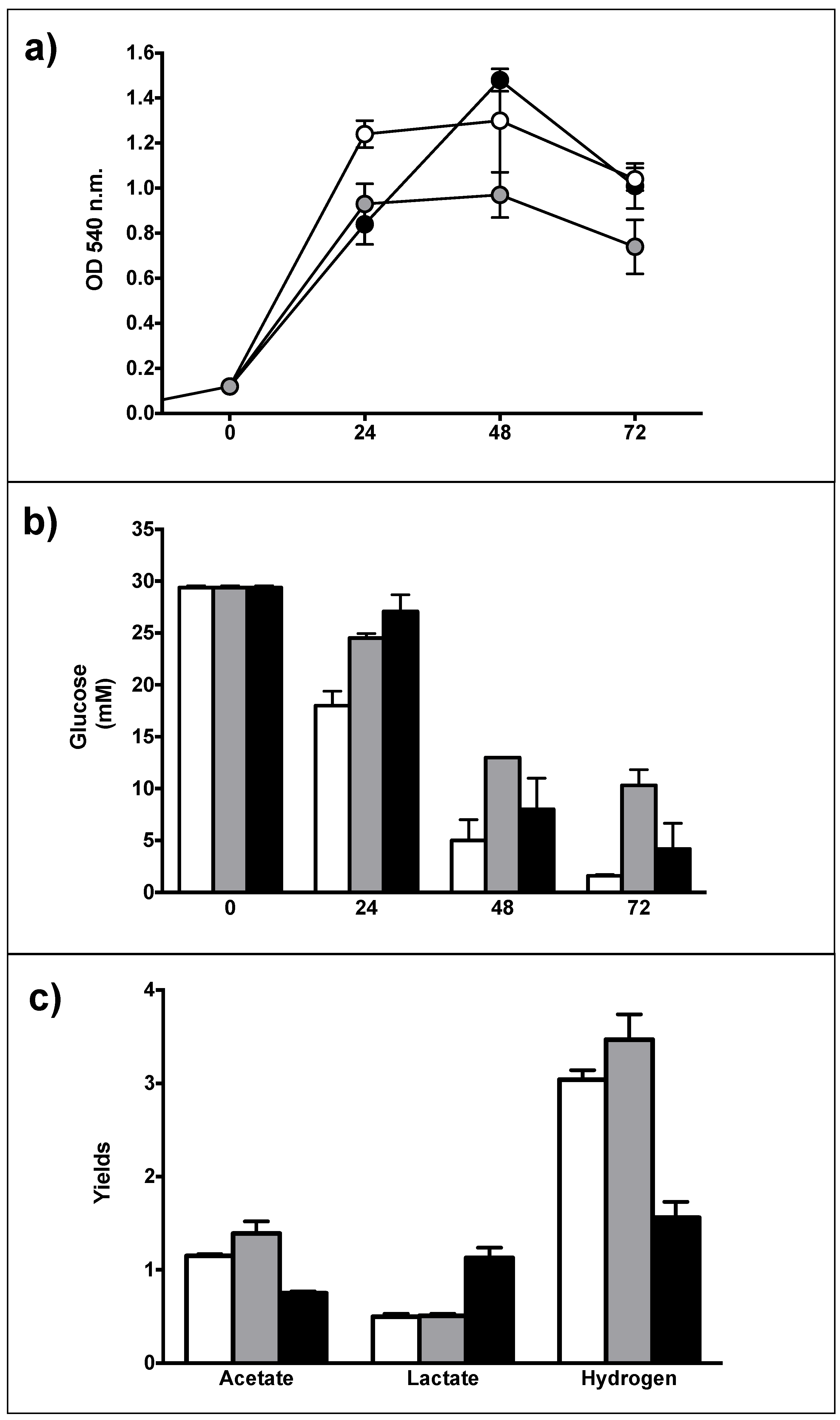
3.2. Generation of Recombinant Strains
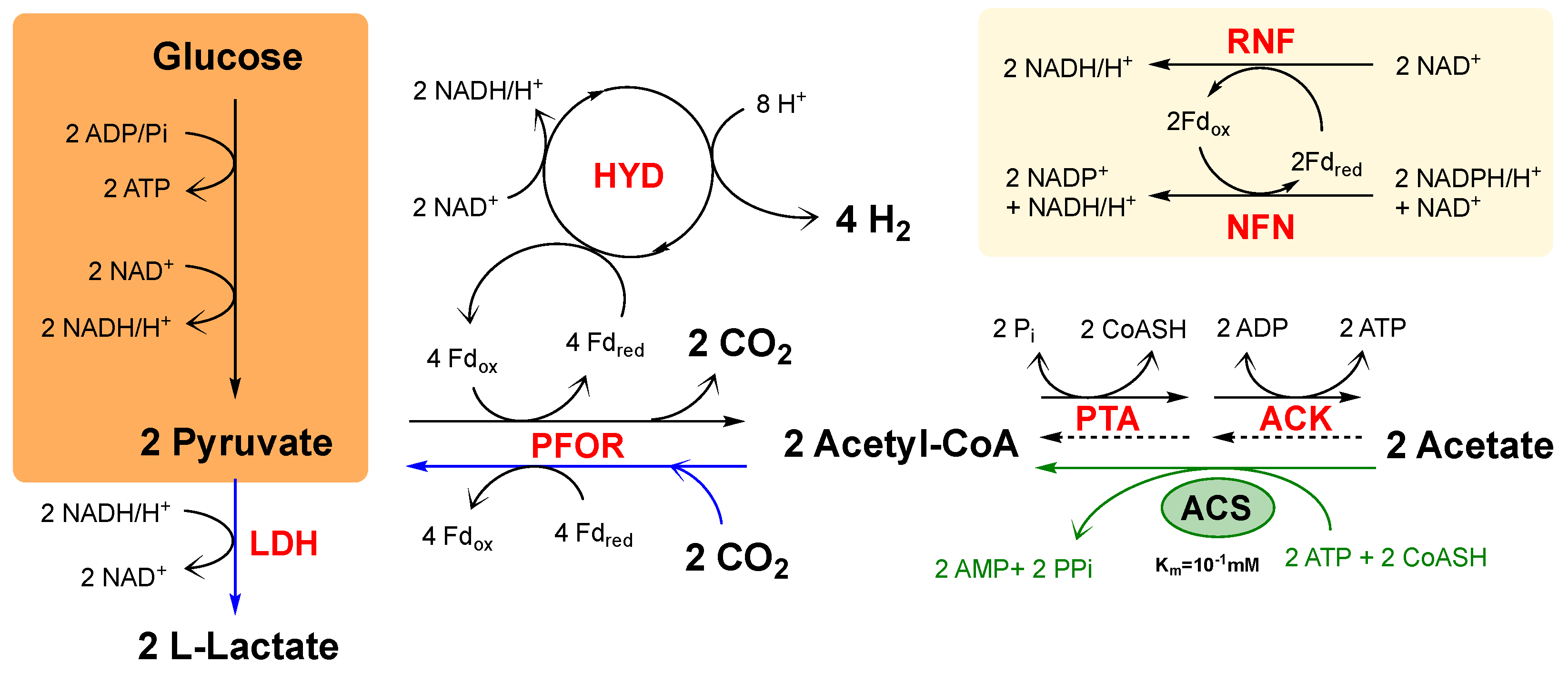
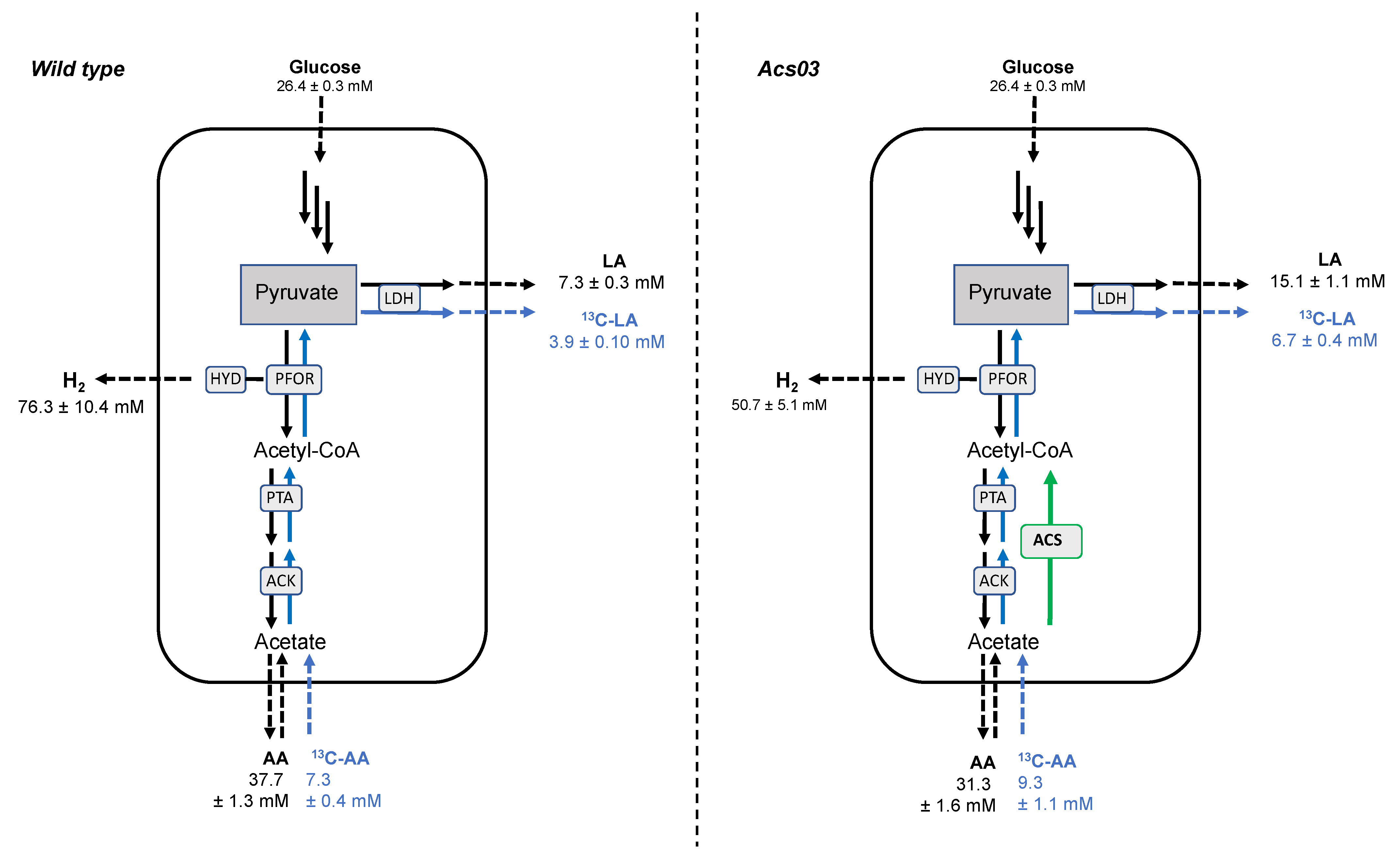
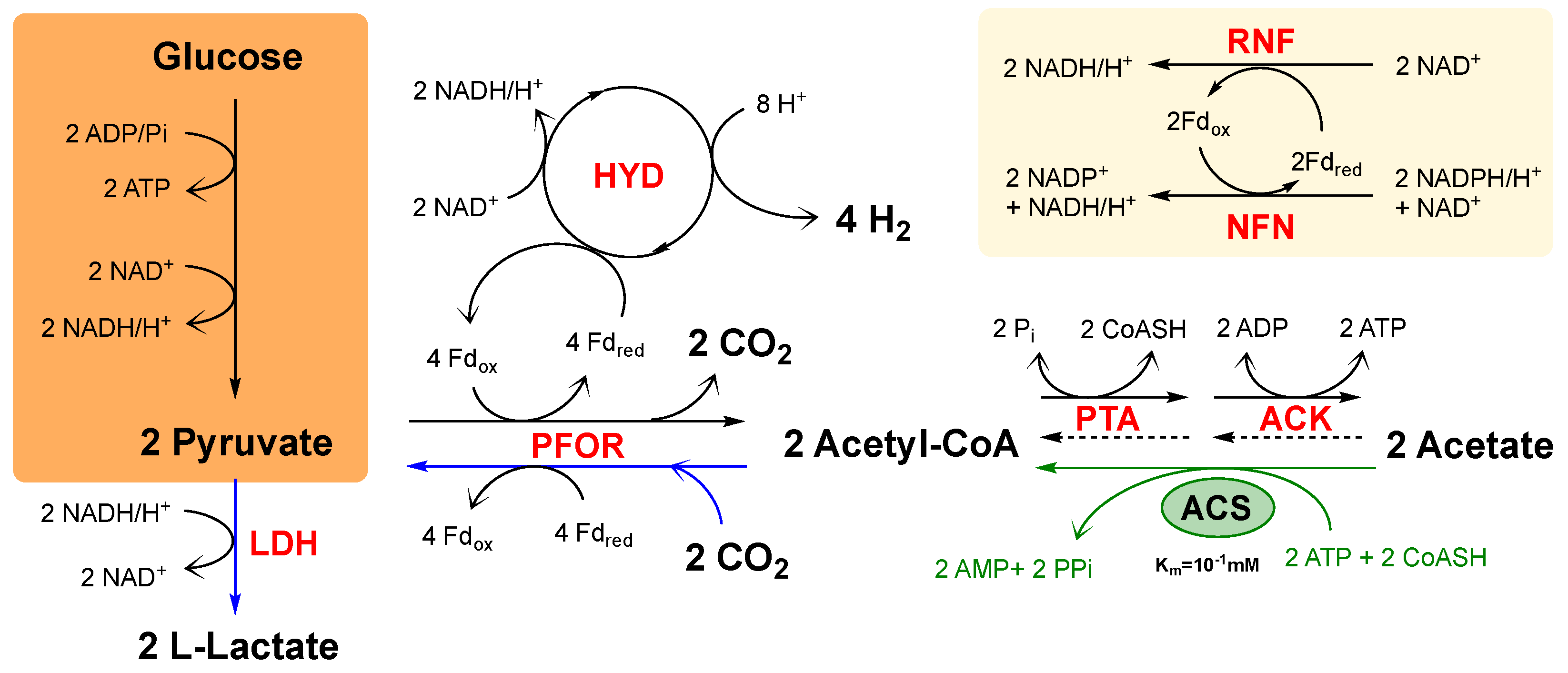
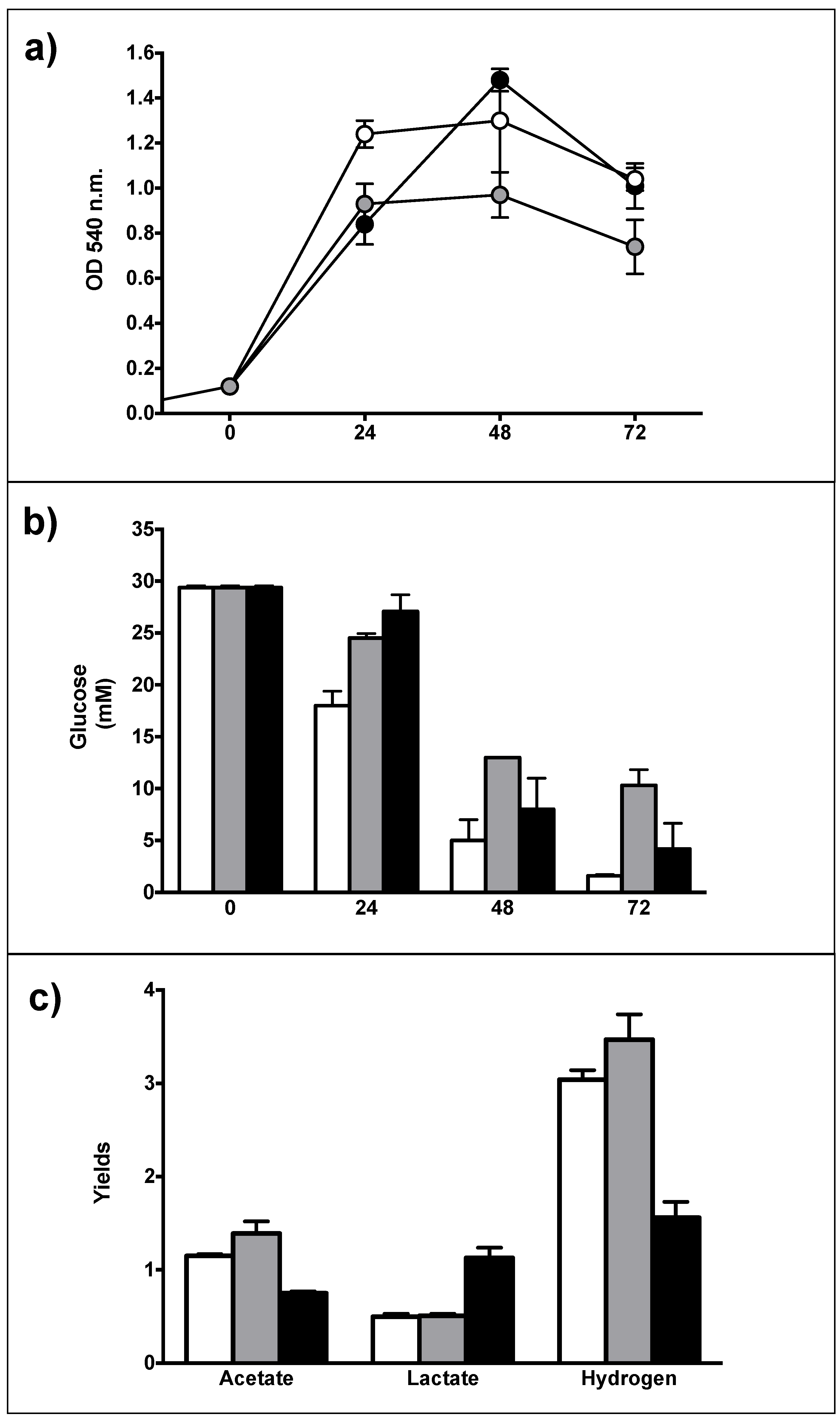
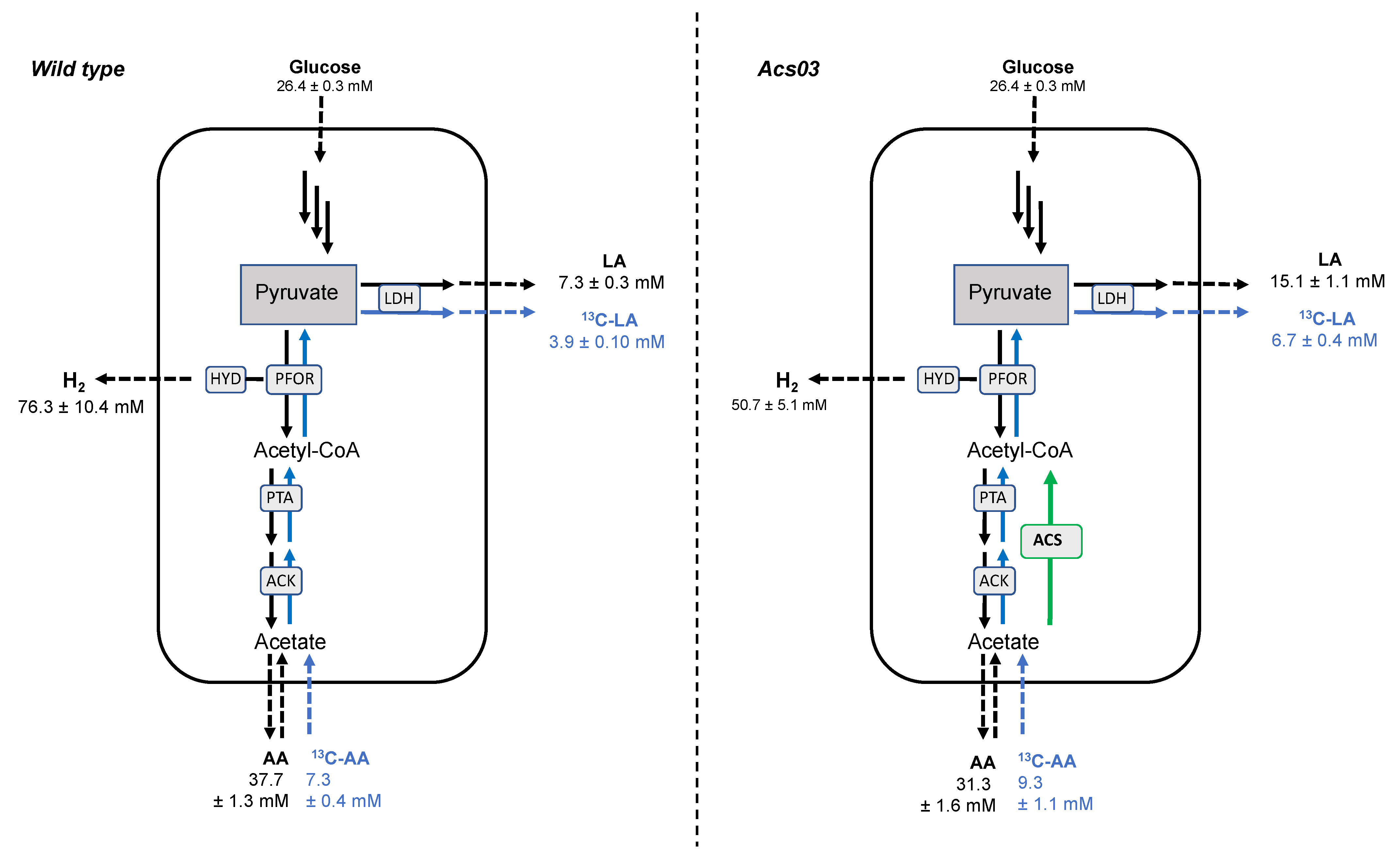
3.3. Catabolic and Anabolic Origins of Lactate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zeldes, B.M.; Keller, M.W.; Loder, A.J.; Straub, C.T.; Adams, M.W.W.; Kelly, R.M. Extremely thermophilic microorganisms as metabolic engineering platforms for production of fuels and industrial chemicals. Front. Microbiol. 2015, 6, 1209. [Google Scholar] [CrossRef] [Green Version]
- Frock, A.D.; Kelly, R.M. Extreme thermophiles: Moving beyond single-enzyme biocatalysis. Curr. Opin. Chem. Eng. 2012, 1, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Verhaart, M.R.A.; Bielen, A.A.M.; Van Der Oost, J.; Stams, A.J.M.; Kengen, S.W.M. Hydrogen production by hyperthermophilic and extremely thermophilic bacteria and archaea: Mechanisms for reductant disposal. Environ. Technol. 2010, 31, 993–1003. [Google Scholar] [CrossRef]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy Conservation in Chemotrophic Anaerobic Bacteria. Am. Soc. Microbiol. Vol. 1977, 41, 100–180. [Google Scholar] [CrossRef]
- Huber, R.; Hannig, M. Thermotogales. Prokaryotes 2006, 899–922. [Google Scholar] [CrossRef]
- Bhandari, V.; Gupta, R.S. The Phylum Thermotogae; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Pradhan, N.; Dipasquale, L.; D’Ippolito, G.; Panico, A.; Lens, P.N.L.; Esposito, G.; Fontana, A. Hydrogen production by the thermophilic bacterium Thermotoga neapolitana. Int. J. Mol. Sci. 2015, 16, 12578–12600. [Google Scholar] [CrossRef] [Green Version]
- Belahbib, H.; Summers, Z.M.; Fardeau, M.L.; Joseph, M.; Tamburini, C.; Dolla, A.; Ollivier, B.; Armougom, F. Towards a congruent reclassification and nomenclature of the thermophilic species of the genus Pseudothermotoga within the order Thermotogales. Syst. Appl. Microbiol. 2018, 41, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Wang, Q.; Rupani, P.F.; Krishnan, S.; Ahmad, F.; Rezania, S.; Rashid, M.A.; Sha, C.; Md Din, M.F. Biohydrogen production via thermophilic fermentation: A prospective application of Thermotoga species. Energy 2020, 197, 117199. [Google Scholar] [CrossRef]
- Lanzilli, M.; Esercizio, N.; Vastano, M.; Xu, Z.; Nuzzo, G.; Gallo, C.; Manzo, E.; Fontana, A.; D’Ippolito, G. Effect of cultivation parameters on fermentation and hydrogen production in the phylum Thermotogae. Int. J. Mol. Sci. 2021, 22, 341. [Google Scholar] [CrossRef]
- Jannasch, H.W.; Huber, R.; Belkin, S.; Stetter, K.O. Thermotoga neapolitana sp. nov. of the extremely thermophilic, eubacterial genus Thermotoga. Arch. Microbiol. 1988, 150, 103–104. [Google Scholar] [CrossRef]
- Dipasquale, L.; D’Ippolito, G.; Fontana, A. Capnophilic lactic fermentation and hydrogen synthesis by Thermotoga neapolitana: An unexpected deviation from the dark fermentation model. Int. J. Hydrog. Energy 2014, 39, 4857–4862. [Google Scholar] [CrossRef]
- Dipasquale, L.; Pradhan, N.; Ippolito, G. Potential of Hydrogen Fermentative Pathways in Marine Thermophilic Bacteria: Dark Fermentation and Capnophilic Lactic Fermentation in Thermotoga and Pseudothermotoga Species. In Grand Challenges in Marine Biotechnology, Grand Challenges in Biology and Biotechnology; Springer: Cham, Switzerland, 2018; pp. 217–235. ISBN 9783319690759. [Google Scholar]
- d’Ippolito, G.; Dipasquale, L.; Fontana, A. Recycling of Carbon Dioxide and Acetate as Lactic Acid by the Hydrogen-Producing Bacterium Thermotoga neapolitana. ChemSusChem 2014, 7, 2678–2683. [Google Scholar] [CrossRef]
- Nuzzo, G.; Landi, S.; Esercizio, N.; Manzo, E.; Fontana, A.; D’Ippolito, G. Capnophilic lactic fermentation from Thermotoga neapolitana: A resourceful pathway to obtain almost enantiopure L-lactic acid. Fermentation 2019, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, N.; Dipasquale, L.; d’Ippolito, G.; Fontana, A.; Panico, A.; Pirozzi, F.; Lens, P.N.L.; Esposito, G. Model development and experimental validation of capnophilic lactic fermentation and hydrogen synthesis by Thermotoga neapolitana. Water Res. 2016, 99, 225–234. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Landi, S.; Esercizio, N.; Lanzilli, M.; Vastano, M.; Dipasquale, L.; Pradhan, N.; Fontana, A. CO2-Induced Transcriptional Reorganization: Molecular Basis of Capnophillic Lactic Fermentation in Thermotoga neapolitana. Front. Microbiol. 2020, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, A.J. The Acetate Switch. Microbiol. Mol. Biol. Rev. 2005, 69, 12–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, P. Acyl adenylates; an enzymatic mechanism of acetate activation. J. Biol. Chem. 1956, 222, 991–1013. [Google Scholar] [CrossRef]
- Chou, T.C.; Lipmann, F. Separation of acetyl transfer enzymes in pigeon liver extract. J. Biol. Chem. 1952, 196, 89–103. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Zhang, Y.; Siewers, V.; Chen, Y.; Nielsen, J. Microbial acetyl-CoA metabolism and metabolic engineering. Metab. Eng. 2015, 28, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Park, J.S.; Kang, C.D.; Cho, H.Y.; Yang, D.; Lee, S.; Cho, K.M. Introduction of a bacterial acetyl-CoA synthesis pathway improves lactic acid production in Saccharomyces cerevisiae. Metab. Eng. 2016, 35, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, N.; Dipasquale, L.; d’Ippolito, G.; Panico, A.; Lens, P.N.L.; Esposito, G.; Fontana, A. Hydrogen and lactic acid synthesis by the wild-type and a laboratory strain of the hyperthermophilic bacterium Thermotoga neapolitana DSMZ 4359T under capnophilic lactic fermentation conditions. Int. J. Hydrog. Energy 2017, 42, 16023–16030. [Google Scholar] [CrossRef]
- Pradhan, N.; Dipasquale, L.; D’Ippolito, G.; Fontana, A.; Panico, A.; Lens, P.N.L.; Pirozzi, F.; Esposito, G. Kinetic modeling of fermentative hydrogen production by Thermotoga neapolitana. Int. J. Hydrog. Energy 2016, 41, 4931–4940. [Google Scholar] [CrossRef]
- Munro, S.A.; Zinder, S.H.; Walker, L.P. The fermentation stoichiometry of Thermotoga neapolitana and influence of temperature, oxygen, and pH on hydrogen production. Biotechnol. Prog. 2009, 25, 1035–1042. [Google Scholar] [CrossRef]
- Kobayashi, J.; Furukawa, M.; Ohshiro, T.; Suzuki, H. Thermoadaptation-directed evolution of chloramphenicol acetyltransferase in an error-prone thermophile using improved procedures. Appl. Genet. Mol. Biotechnol. 2015, 99, 5563–5572. [Google Scholar] [CrossRef]
- Han, D.; Norris, S.M.; Xu, Z. Construction and transformation of a Thermotoga-E. coli shuttle vector. BMC Biotechnol. 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Käslin, S.A.; Childers, S.E.; Noll, K.M. Membrane-associated redox activities in Thermotoga neapolitana. Arch. Microbiol. 1998, 170, 297–303. [Google Scholar] [CrossRef]
- Han, D.; Xu, H.; Puranik, R.; Xu, Z. Natural transformation of Thermotoga sp. strain RQ7. BMC Biotechnol. 2014, 14. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.S.; Vargas, M.; Mityas, C.; Noll, K.M. Liposome-mediated DNA uptake and transient expression in Thermotoga. Extremophiles 2001, 5. [Google Scholar] [CrossRef]
- Xu, H.; Han, D.; Xu, Z. Expression of Heterologous Cellulases in Thermotoga sp. Strain RQ2. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Xu, Z. Development of a pyrE-based selective system for Thermotoga sp. strain RQ7. Extremophiles 2017, 21, 297–306. [Google Scholar] [CrossRef]
- Singh, R.; White, D.; Demirel, Y.; Kelly, R.; Noll, K.; Blum, P. Uncoupling fermentative synthesis of molecular hydrogen from biomass formation in Thermotoga maritima. Appl. Environ. Microbiol. 2018, 84, 998–1016. [Google Scholar] [CrossRef] [Green Version]
- White, D.; Singh, R.; Rudrappa, D.; Mateo, J.; Kramer, L.; Freese, L.; Blum, P. Contribution of pentose catabolism to molecular hydrogen formation by targeted disruption of arabinose isomerase (araA) in the hyperthermophilic bacterium Thermotoga maritima. Appl. Environ. Microbiol. 2017, 83, 2631–2647. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; White, D.; Blum, P. Identification of the ATPase Subunit of the Primary Maltose Transporter in the Hyperthermophilic Anaerobe Thermotoga maritima. Appl. Environ. Microbiol. 2017, 83, e00930-17. [Google Scholar] [CrossRef] [Green Version]
- Greening, C.; Trchounian, A.; Costa, K.; Müller, V.; Schuchmann, K.; Chowdhury, N.P. Complex Multimeric [FeFe] Hydrogenases: Biochemistry, Physiology and New Opportunities for the Hydrogen Economy. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Xiao, Y.; Ruan, Z.; Liu, Z.; Wu, S.G.; Varman, A.M.; Liu, Y.; Tang, Y.J. Engineering Escherichia coli to convert acetic acid to free fatty acids. Biochem. Eng. J. 2013, 76, 60–69. [Google Scholar] [CrossRef]
- Zhu, N.; Xia, H.; Wang, Z.; Zhao, X.; Chen, T. Engineering of Acetate Recycling and Citrate Synthase to Improve Aerobic Succinate Production in Corynebacterium glutamicum. PLoS ONE 2012, 8. [Google Scholar] [CrossRef] [Green Version]
- Noh, M.H.; Lim, H.G.; Woo, S.H.; Song, J.; Jung, G.Y. Production of itaconic acid from acetate by engineering acid-tolerant Escherichia coli W. Biotechnol. Bioeng. 2018, 115, 729–738. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, W.; Chen, H.; Liu, B.; Lin, B.; Tao, Y. Metabolic engineering for efficient supply of acetyl-CoA from different carbon sources in Escherichia coli. Microb. Cell Fact. 2019, 18, 130. [Google Scholar] [CrossRef]
- Shi, L.L.; Da, Y.Y.; Zheng, W.T.; Chen, G.Q.; Li, Z.J. Production of polyhydroxyalkanoate from acetate by metabolically engineered Aeromonas hydrophilia. J. Biosci. Bioeng. 2020, 130, 290–294. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Strain | Selection | Strategy | Transformation Technique | Target | Ref. |
|---|---|---|---|---|---|
| T. maritima | ant (kanamycin) | Replicating plasmid | sph-DOTAP/ electroporation | kan heterologous expression | [27] |
| ant (kanamycin) | Replicating plasmid | sph-DOTAP | kan heterologous expression | [30] | |
| aux (uracil) | Chromosomal recombination | sph-DOTAP/ natural transformation | knock-out & knock-in araA | [34] | |
| aux (uracil) | Chromosomal recombination | natural transformation | knock-out malkX genes | [33] | |
| ant (kanamycin) | Chromosomal recombination | electroporation | transient inactivation of ldh | [33] | |
| aux (uracil) | Chromosomal recombination | electroporation | knock-in malk3 | [33] | |
| T. sp. RQ7 | ant (kanamycin) | Replicating plasmid | sph-DOTAP/ electroporation | kan heterologous expression | [27] |
| ant (kanamycin) | Replicating plasmid | natural transformation | kan heterologous expression | [29] | |
| aux (uracil) | Replicating plasmid | natural transformation | pyrE heterologous expression | [32] | |
| T. sp. RQ2 | ant (kanamycin) | Replicating plasmid | natural transformation | kan + amya-celB heterologous expression | [31] |
| ant (kanamycin) | Replicating plasmid | natural transformation | kan + xynb-celB heterologous expression | [31] | |
| ant (kanamycin) | Replicating plasmid | natural transformation | kan + xynb-celA heterologous expression | [31] | |
| T. neapolitana | ant (chloramphenicol) | Replicating plasmid | sph-DOTAP | cat heterologous expression | [30] |
| LA/AA | |
|---|---|
| wt | 0.44 |
| Empty vector | 0.36 |
| Acs03 | 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esercizio, N.; Lanzilli, M.; Vastano, M.; Xu, Z.; Landi, S.; Caso, L.; Gallo, C.; Nuzzo, G.; Manzo, E.; Fontana, A.; et al. Improvement of CO2 and Acetate Coupling into Lactic Acid by Genetic Manipulation of the Hyperthermophilic Bacterium Thermotoga neapolitana. Microorganisms 2021, 9, 1688. https://doi.org/10.3390/microorganisms9081688
Esercizio N, Lanzilli M, Vastano M, Xu Z, Landi S, Caso L, Gallo C, Nuzzo G, Manzo E, Fontana A, et al. Improvement of CO2 and Acetate Coupling into Lactic Acid by Genetic Manipulation of the Hyperthermophilic Bacterium Thermotoga neapolitana. Microorganisms. 2021; 9(8):1688. https://doi.org/10.3390/microorganisms9081688
Chicago/Turabian StyleEsercizio, Nunzia, Mariamichela Lanzilli, Marco Vastano, Zhaohui Xu, Simone Landi, Lucio Caso, Carmela Gallo, Genoveffa Nuzzo, Emiliano Manzo, Angelo Fontana, and et al. 2021. "Improvement of CO2 and Acetate Coupling into Lactic Acid by Genetic Manipulation of the Hyperthermophilic Bacterium Thermotoga neapolitana" Microorganisms 9, no. 8: 1688. https://doi.org/10.3390/microorganisms9081688
APA StyleEsercizio, N., Lanzilli, M., Vastano, M., Xu, Z., Landi, S., Caso, L., Gallo, C., Nuzzo, G., Manzo, E., Fontana, A., & d’Ippolito, G. (2021). Improvement of CO2 and Acetate Coupling into Lactic Acid by Genetic Manipulation of the Hyperthermophilic Bacterium Thermotoga neapolitana. Microorganisms, 9(8), 1688. https://doi.org/10.3390/microorganisms9081688