
Isolation and Characterization of Lactic Acid Bacteria and Yeasts from Typical Bulgarian Sourdoughs



Abstract
1. Introduction
2. Materials and Methods
2.1. Sourdoughs
2.2. Physico-Chemical Characterization of Sourdoughs
2.3. Lactic Acid Bacteria and Yeast Enumeration and Isolation
2.4. Molecular Identification of Lactic Acid Bacteria by 16S rDNA Sequence Analysis
2.5. Molecular Identification of Yeast by ITS1-5.8S-ITS2 rRNA Gene Sequence Analysis
2.6. Screening of the LAB Isolates for Amylolytic and Proteolytic Activity and Acid-Producing Capacity
2.7. Antimicrobial Activity of the LAB Strains
2.8. Nucleotide Sequence Accession Number
2.9. Statistical Analysis
3. Results and Discussion
3.1. Physico-Chemical Characterization of Sourdoughs
3.2. Enumeration of LAB and Yeasts
3.3. Molecular Identification of LAB by 16S rDNA Sequence Analysis
3.4. Molecular Identification of Yeasts by ITS1-5.8S-ITS2 Region Sequence Analysis
3.5. LAB Selection
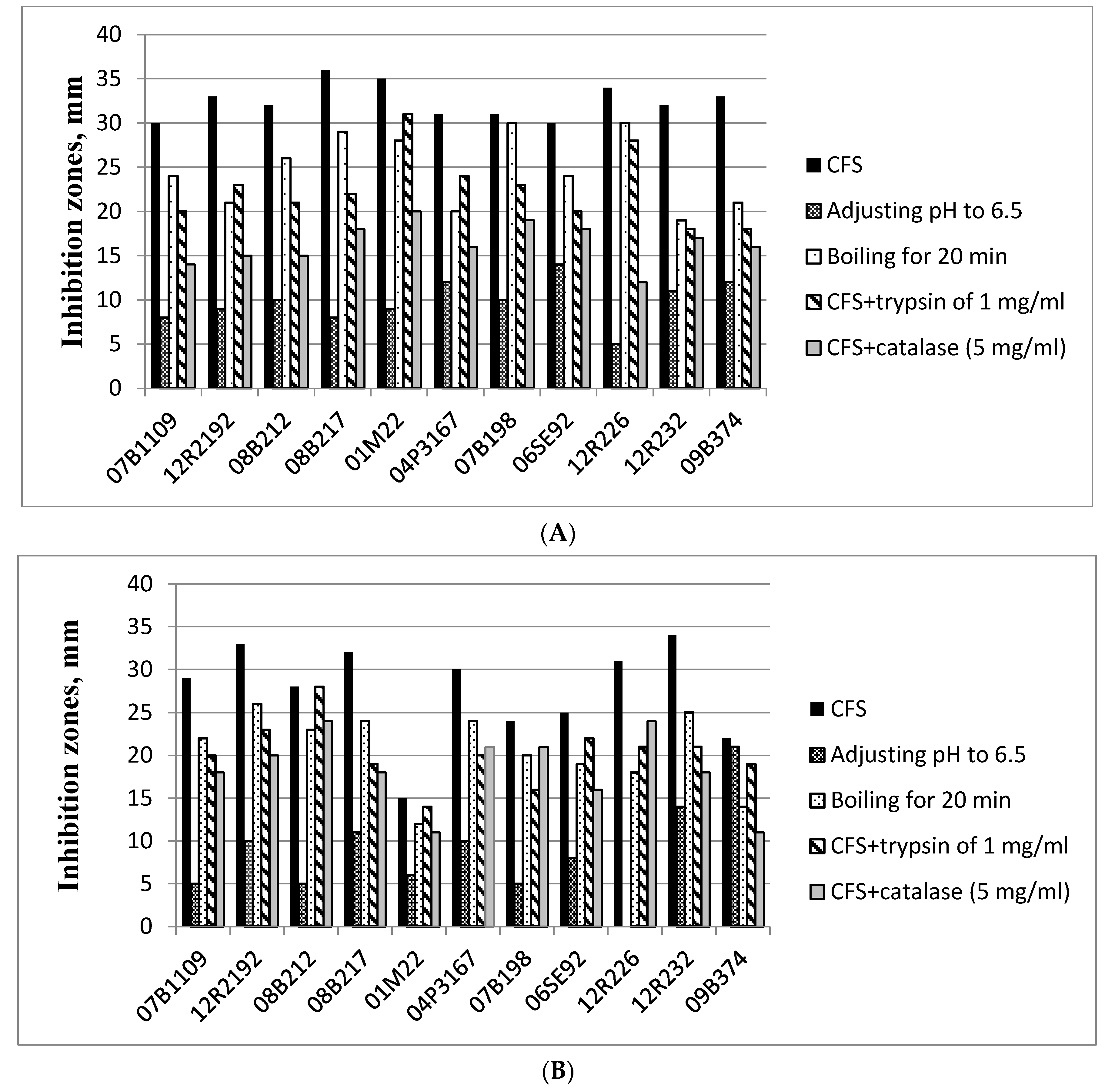
3.6. Antibacterial Activity of the LAB Strains
3.7. Antifungal Activity of the LAB Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Regulation (EU) No 1151/2012 of the European Parliament and of the Council of 21 November 2012 on quality schemes for agricultural products and foodstuffs. Off. J. Eur. Union 2012, 343, 1–29.
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.M.; Weckxa, S. Microbial ecology of sourdough fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef]
- Reese, A.T.; Madden, A.A.; Joossens, M.; Lacaze, G.; Dunn, R.R. Influences of ingredients and bakers on the bacteria and fungi in sourdough starters and bread. mSphere 2020, 5, e00950-19. [Google Scholar] [CrossRef]
- Comasio, A.; Verce, M.; Van Kerrebroeck, S.; De Vuyst, L. Diverse Microbial Composition of Sourdoughs from Different Origins. Front. Microbiol. 2020. [CrossRef]
- Landis, E.A.; Oliverio, A.M.; McKenney, E.A.; Nichols, L.M.; Kfoury, N.; Biango-Daniels, M.; Shell, L.K.; Madden, A.A.; Shapiro, L.; Sakunala, S.; et al. The diversity and function of sourdough starter microbiomes. eLife 2021. [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Casado, A.; Álvarez, A.; González, L.; Fernández, D.; Marcos, J.L.; Tornadijo, M.E. Effect of fermentation on microbiological, physicochemical and physical characteristics of sourdough and impact of its use on bread quality. Czech J. Food Sci. 2017, 35, 496–506. [Google Scholar]
- Koistinen, V.M.; Mattila, O.; Katina, K.; Poutanen, K.; Aura, A.M.; Hanhineva, K. Metabolic profiling of sourdough fermented wheat and rye bread. Sci. Rep. 2018, 8, 5684. [Google Scholar] [CrossRef] [PubMed]
- Gobbett, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 57–69. [Google Scholar] [CrossRef]
- Arendt, E.K.; Ryan, L.A.M.; Dal Bello, F. Impact of sourdough on the texture of bread. Food Microbiol. 2007, 24, 165–174. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Celano, G.; De Angelis, M.; Minervini, F.; Gobbetti, M. Different flour microbial communities drive to sourdoughs characterized by diverse bacterial strains and free amino acid profiles. Front. Microbiol. 2016, 7, 1770. [Google Scholar] [CrossRef]
- Minervini, F.; Lattanzi, A.; De Angelis, M.; Celano, G.; Gobbetti, M. House microbiotas as sources of lactic acid bacteria and yeasts in traditional Italian sourdoughs. Food Microbiol. 2015, 52, 66–76. [Google Scholar] [CrossRef]
- Minervini, F.; Di Cagno, R.; Lattanzi, A.A.; De Angelis, M.; Antonielli, L.; Cardinali, G.; Cappelle, S.; Gobbetti, M. Lactic acid bacterium and yeast microbiotas of 19 sourdoughs used for traditional/typical Italian breads: Interactions between ingredients and microbial species diversity. Appl. Environ. Microbiol. 2012, 78, 251–1264. [Google Scholar] [CrossRef]
- De Vuyst, L.; Schrijvers, V.; Paramithiotis, S.; Hoste, B.; Vancanneyt, M.; Swings, J.; Kalantzopoulos, G.; Tsakalidou, E.; Messens, W. 2002. The biodiversity of lactic acid bacteria in Greek traditional wheat sourdoughs is reflected in both composition and metabolite formation. Appl. Environ. Microbiol. 2002, 68, 6059–6069. [Google Scholar] [CrossRef]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Lhomme, E.; Lattanzi, A.; Xavier, D.; Minervini, F.; De Angelis, M.; Lacaze, G.; Onno, B.; Gobbetti, M. Lactic acid bacterium and yeast microbiotas of sixteen French traditional sourdoughs. Int. J. Food Microbiol. 2015, 215, 161–170. [Google Scholar] [CrossRef]
- Vogelmann, S.; Seitter, M.; Singer, U.; Brandt, M.J.; Hertel, C. Adaptability of lactic acid bacteria and yeasts to sourdoughs prepared from cereals, pseudocereals and cassava and use of competitive strains as starters. Int. J. Food Microbiol. 2009, 130, 205–212. [Google Scholar] [CrossRef]
- Siragusa, S.; Di Cagno, R.; Ercolini, D.; Minervini, F.; Gobbetti, M.; De Angelis, M. Taxonomic structure and monitoring of the dominant population of lactic acid bacteria during wheat flour sourdough type I propagation using Lactobacillus sanfranciscensis starters. Appl. Environ. Microbiol. 2009, 75, 1099–1109. [Google Scholar] [CrossRef]
- Capriles, V.D.; Areas, J.A.G. Novel approaches in gluten-free bread making: Interface between food science, nutrition, and health. Compr. Rev. Food Sci. Food Saf. 2014, 13, 871–890. [Google Scholar] [CrossRef]
- Matos, M.E.; Rosell, C.M. A review: Understanding gluten free dough for reaching breads with physical quality and nutritional balance. J. Sci. Food Agric. 2015, 95, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Santarelli, S.; Claus, S.; Sanderson, J.; Hudspith, B.N.; Brostoff, J.; Ward, J.L.; Lovegrove, A.; Shewry, P.R.; Jones, H.E.; et al. Effect of breadmaking process on in vitro gut microbiota parameters in irritable bowel syndrome. PLoS ONE 2014, 9, e111225. [Google Scholar] [CrossRef]
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and cereal fermentation in a nutritional perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Rosén, L.; Östman, E.M.; Shewry, P.R.; Ward, J.L.; Andersson, A.A.M.; Piironen, V.; Lampi, A.M.; Rakszegi, M.; Bedő, Z.; Björc, I. Postprandial glycemia, insulinemia, and satiety responses in healthy subjects after whole grain rye bread made from different rye varieties. J. Agric. Food Chem. 2011, 59, 12139–12148. [Google Scholar] [CrossRef] [PubMed]
- Lacaze, G.; Wick, M.; Cappelle, S. Emerging fermentation technologies: Development of novel sourdoughs. Food Microbiol. 2007, 24, 155–160. [Google Scholar] [CrossRef]
- Gänzle, M.; Ripari, V. Composition and function of sourdough microbiota: From ecological theory to bread quality. Int. J. Food Microbiol. 2016, 239, 19–25. [Google Scholar] [CrossRef]
- Demirbaş, F.; İspirli, H.; Kurnaz, A.A.; Yilmaz, M.T.; Dertli, E. Antimicrobial and functional properties of lactic acid bacteria isolated from sourdoughs. LWT Food Sci. Technol. 2017, 79, 361–366. [Google Scholar] [CrossRef]
- de Vuyst, L.; Van Kerrebroeck, S.; Leroy, F. Microbial ecology and process technology of sourdough fermentation. Adv. Appl. Microbiol. 2017, 100, 49–160. [Google Scholar]
- Pétel, C.; Onno, B.; Prost, C. Sourdough volatile compounds and their contribution to bread: A review. Trends Food Sci. Technol. 2017, 59, 105–123. [Google Scholar] [CrossRef]
- de Vuyst, L.; Leroy, F. Bacteriocins from lactic acid bacteria: Production, purification, and food applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- Settanni, L.; Corsetti, A. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol. 2008, 121, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Negash, A.W.; Tsehai, B.A. Current applications of bacteriocin. Int. J. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef]
- Cizeikiene, D.; Juodeikiene, G.; Paskevicus, A.; Bartkiene, E. Antimicrobial activity of lactic acid bacteria against pathogenic and spoilage microorganism isolated from food and their control in wheat bread. Food Control 2013, 3, 539–545. [Google Scholar] [CrossRef]
- Magnusson, J.; Schnürer, J. Lactobacillus coryniformis subsp. coryniformis strain Si3 produces a broad-spectrum proteinaceous antifungal compound. Appl. Environ. Microbiol. 2001, 67, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Marco Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [PubMed]
- Gerbaldo, G.A.; Barberis, C.; Pascual, L.; Dalcero, A.; Barberis, L. Antifungal activity of two Lactobacillus strains with potential probiotic properties. FEMS Microbiol. Lett. 2012, 332, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ogunbanwo, S.T.; Sanni, A.I.; Onilude, A.A. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol. 2003, 2, 219–227. [Google Scholar]
- Muthusamy, K.; Soundharrajan, I.; Srisesharam, S.; Kim, D.; Kuppusamy, P.; Lee, K.D.; Choi, K.C. Probiotic characteristics and antifungal activity of Lactobacillus plantarum and its impact on fermentation of Italian ryegrass at low moisture. Appl. Sci. 2020, 10, 417. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Next-generation approaches to the microbial ecology of food fermentations. BMB Rep. 2012, 45, 377–389. [Google Scholar] [CrossRef]
- Bessmeltseva, M.; Viiard, E.; Simm, J.; Paalme, T.; Sarand, I. Evolution of bacterial consortia in spontaneously started rye sourdoughs during two months of daily propagation. PLoS ONE 2014, 9, e95449. [Google Scholar] [CrossRef]
- Viiard, E.; Mihhalevski, A.; Rühka, T.; Paalme, T.; Sarand, I. Evaluation of the microbial community in industrial rye sourdough upon continuous back-slopping propagation revealed Lactobacillus helveticus as the dominant species. J. Appl. Microbiol. 2013, 114, 404–412. [Google Scholar] [CrossRef]
- Meroth, C.B.; Hammes, W.P.; Hertel, C. Identification and population dynamics of yeasts in sourdough fermentation processes by PCR-denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2003, 69, 7453–7461. [Google Scholar] [CrossRef]
- Rossetti, L.; Giraffa, G. Rapid identification of dairy lactic acid bacteria by M13-generated, RAPD-PCR fingerprint databases. J. Microbiol. Methods 2005, 63, 135–144. [Google Scholar] [CrossRef]
- Jeyaram, K.; Singh, W.M.; Capece, A.; Romano, P. Molecular identification of yeast species associated with ‘Hamei’-a traditional starter used for rice wine production in Manipur, India. Int. J. Food Microbiol. 2008, 124, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Lopandica, K.; Zelgerb, S.; Banszky, L.K.; Eliskases-Lechnerd, F.; Prillinger, H. Identification of yeasts associated with milk products using traditional and molecular techniques. Food Microbiol. 2006, 23, 341–350. [Google Scholar] [CrossRef]
- Petkova, M.; Stefanova, P.; Gotcheva, V.; Kuzmanova, I.; Angelov, A. Microbiological and physicochemical characterization of traditional Bulgarian sourdoughs and screening of lactic acid bacteria for amylolytic activity. J. Chem. Technol. Metall. 2020, 55, 921–934. [Google Scholar]
- Fleet, G.H. Yeasts in foods and beverages: Impact on product quality and safety. Curr. Opin. Biotechnol. 2007, 18, 170–175. [Google Scholar] [CrossRef]
- Martin, K.J.; Rygiewicz, P.T. Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiol. 2005, 5. [Google Scholar] [CrossRef]
- De Filippis, F.; Parente, E.; Ercolini, D. Metagenomics insights into food fermentations. Microb. Biotechnol. 2017, 10, 91–102. [Google Scholar] [CrossRef]
- Jagadeesana, B.; Gerner-Smidt, P.; Allard, M.W.; Leuillet, S.; Winkler, A.; Xiao, Y.; Chaffron, S.; Van Der Vossen, J.; Tang, S.; Katase, M.; et al. The use of next generation sequencing for improving food safety: Translation into practice. Food Microbiol. 2019, 79, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Stefan Weckx, S.; Van Kerrebroeck, S.; De Vuyst, L. Omics approaches to understand sourdough fermentation processes. Int. J. Food Microbiol. 2019, 302, 90–102. [Google Scholar] [CrossRef]
- Menezes, L.A.A.; SavoSardaro, M.L.; Duarte, R.T.D.; Mazzon, R.R.; Neviani, E.; Gatti, M.; De Dea Lindner, J. Sourdough bacterial dynamics revealed by metagenomic analysis in Brazil. Food Microbiol. 2020, 85, 103302. [Google Scholar] [CrossRef]
- Bondia-Pons, I.; Aura, A.-M.; Vuorela, S.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Rye phenolics in nutrition and health. J. Cereal Sci. 2009, 49, 323–336. [Google Scholar] [CrossRef]
- Sanz-Penella, J.M.; Tamayo-Ramos, J.A.; Haros, M. Application of bifidobacteria as starter culture in whole wheat sourdough breadmaking. Food Bioprocess. Technol. 2012, 5, 2370–2380. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef]
- Agati, V.; Guyot, J.; Morlon-Guyot, P.; Talamond, P.; Hounhouigan, D.J. Isolation and characterization of new amylolytic strains of Lactobacillus fermentum from fermented maize doughs (mawe and ogi) from Benin. J. Appl. Microbiol. 1998, 85, 512–520. [Google Scholar] [CrossRef]
- Chen, P.H.; Chen, R.Y.; Chou, J.Y. Screening and evaluation of yeast antagonists for biological control of Botrytis cinerea on strawberry fruits. Mycobiology 2018, 46, 33–46. [Google Scholar] [CrossRef]
- Hebert, E.M.; Raya, R.R.; de Giori, G.S. Nutritional requirements and nitrogen-dependent regulation of proteinase activity of Lactobacillus helveticus CRL (1062). Appl. Env. Microbiol. 2000, 66, 5316–5321. [Google Scholar] [CrossRef]
- Yang, E.; Fan, L.; Jiang, Y.; Doucette, C.; Fillmore, S. Antimicrobial activity of bacteriocin-producing lactic acid bacteria isolated from cheeses and yogurts. AMB Express 2012, 2, 48. [Google Scholar] [CrossRef]
- Vera, A.; Ly-Chatain, M.H.; Rigobello, V.; Demarigny, Y. Description of a French natural wheat sourdough over 10 consecutive days focussing on the lactobacilli present in the microbiota. Antonie Van Leeuwenhoek 2012, 101, 369–377. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Follador, R. Metabolism of oligosaccharides and starch in lactobacilli: A review. Front. Microbiol. 2012, 3, 1–15. [Google Scholar] [CrossRef]
- Hayek, S.A.; Ibrahim, S.A. Current limitations and challenges with lactic acid bacteria: A review. Food Nutr. Sci. 2013, 4, 73–87. [Google Scholar] [CrossRef]
- Rice, T.; Sahin, A.W.; Lynch, K.M.; Arendt, E.K.; Coffey, A. Isolation, characterisation and exploitation of lactic acid bacteria capable of efficient conversion of sugars to mannitol. Int. J. Food Microbiol. 2020, 321, 108546. [Google Scholar] [CrossRef]
- Michel, E.; Monfort, C.; Deffrasnes, M.; Guezenec, S.; Lhomme, E.; Barret, M.; Sicard, D.; Dousset, X.; Onno, B. Characterization of relative abundance of lactic acid bacteria species in French organic sourdough by cultural, qPCR and MiSeq high-throughput sequencing methods. Int. J. Food Microbiol. 2016, 239, 35–43. [Google Scholar] [CrossRef]
- Ripari, V.; Cecchi, T.; Berard, E. Microbiological characterisation and volatiles profile of model, ex-novo, and traditional Italian white wheat sourdoughs. Food Chem. 2016, 205, 297–307. [Google Scholar] [CrossRef]
- Dertli, E.; Mercan, E.; Arıcı, M.; Yılmaz, M.T.; Sağdıç, O. Characterisation of lactic acid bacteria from Turkish sourdough and determination of their exopolysaccharide (EPS) production characteristics. LWT Food Sci. Technol. 2016, 71, 116–124. [Google Scholar] [CrossRef]
- Robert, H.; Gabriel, V.; Fontagné-Faucher, C. Biodiversity of lactic acid bacteria in French wheat sourdough as determined by molecular characterization using species-specific PCR. Int. J. Food Microbiol. 2009, 135, 53–59. [Google Scholar] [CrossRef]
- Tan, Q.; Xu, H.; Aguilar, Z.P.; Peng, S.; Dong, S.; Wang, B.; Li, P.; Chen, T.; Xu, F.; Wei, H. Safety assessment and probiotic evaluation of Enterococcus faecium YF5 isolated from sourdough. J. Food Sci. 2013, 78, 4. [Google Scholar] [CrossRef] [PubMed]
- Succi, M.; Reale, A.; Andrighetto, C.; Lombardi, A.; Sorrentino, E.; Coppola, R. Presence of yeasts in southern Italian sourdoughs from Triticum aestivum flour. FEMS Microbiol. Lett. 2003, 225, 143–148. [Google Scholar] [CrossRef]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Lavermicocca, P.; Morea, M.; Baruzzi, F.; Tosti, N.; Gobbetti, M. Phenotypic and molecular identification and clustering of lactic acid bacteria and yeasts from wheat (species Triticum durum and Triticum aestivum) sourdoughs of Southern Italy. Int. J. Food Microbiol. 2001, 64, 95–104. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Müller, M.; Ehrmann, M.; Tsakalidou, A.; Seiler, H.; Vogel, R.; Kalantzopoulos, G. Polyphasic identification of wild yeast strains isolated from Greek sourdoughs. Syst. Appl. Microbiol. 2000, 23, 156–164. [Google Scholar] [CrossRef]
- Marizeth Groenewald, M.; Teun Boekhout, T.; Cécile Neuvéglise, C.; Claude Gaillardin, C.; Piet, W.M.; van Dijck, P.W.M.; Markus Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- Peterson, S.W.; Kurtzman, C.P. Ribosomal RNA sequence divergence among sibling species of yeasts. Syst. Appl. Microbiol. 1991, 14, 124–129. [Google Scholar] [CrossRef]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic acid bacteria isolation from spontaneous sourdough and their characterization including antimicrobial and antifungal properties evaluation. Microorganisms 2020, 8, 64. [Google Scholar] [CrossRef]
- Gotcheva, V.; Petrova, G.; Petkova, M.; Kuzmanova, Y.; Angelov, A. Molecular and in vitro assessment of some probiotic characteristics of amylolytic Lactobacillus plantarum strains from Bulgarian fermented products. Eng. Life Sci. 2018, 18, 820–830. [Google Scholar] [CrossRef]
- Pereira, A.P.M.; Stradiotto, G.C.; Freire, L.; Alvarenga, V.O.; Crucello, A.; Morassi, L.L.P.; Silva, F.P.; .Sant’Ana, A.S. Occurrence and enumeration of rope-producing spore forming bacteria in flour and their spoilage potential in different bread formulations. LWT 2020, 133, 110108. [Google Scholar] [CrossRef]
- Barman, S.; Ghosh, R.; Sengupta, S.; Mandal, N.C. Longterm storage of post-packaged bread by controlling spoilage pathogens using Lactobacillus fermentum C14 isolated from homemade curd. PLoS ONE 2017, 12, e0184020. [Google Scholar] [CrossRef]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic acid bacteria antimicrobial compounds: Characteristics and applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Todorov, S.; Wachsman, M.; Ignatova, T.; Ivanova, I. Lactobacillus plantarum ST16Pa—are we ready to use it as bio-protective culture? Bulg. J. Agric. Sci. 2014, 20, 55–58. [Google Scholar]
- Todorov, S.; Dicks, L. Lactobacillus plantarum isolated from molasses produced bacteriocins active against Gram negative bacteria. Enzym. Microb. Technol. 2005, 36, 318–326. [Google Scholar] [CrossRef]
- Joshi, V.; Sharma, S.; Rana, N. Production, purification, stability and efficacy of bacteriocin from isolates of natural lactic acid fermentation of vegetables. Food Technol. Biotechnol. 2006, 44, 435–439. [Google Scholar]
- Cherif, A.; Ouzari, H.; Daffonchio, D.; Cherif, H.; BenSlama, K.; Hassen, A.; Jaoua, S.; Boudabous, A. Thuricin 7: A novel bacteriocin produced by Bacillus thuringiensis BMG1Æ7, a new strain isolated from soil. Lett. Appl. Microbiol. 2001, 32, 243–247. [Google Scholar] [CrossRef]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Visconti, A. Antifungal activity of phenyllactic acid against molds isolated from bakery products. Appl. Environ. Microbiol. 2003, 69, 634–640. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torino, M.I.; Obregozo, M.D.; de Valdez, G. A ready-to use antifungal starter culture improves the shelf life of packaged bread. J. Food Prot. 2010, 73, 758–762. [Google Scholar] [CrossRef]
- Ryu, E.H.; Yang, E.J.; Woo, E.R.; Chang, H.C. Purification and characterization of antifungal compounds from Lactobacillus plantarum HD1 isolated from kimchi. Food Microbiol. 2014, 41, 19–26. [Google Scholar] [CrossRef]
- Taghi-Zadeh, A.; Nejati, F. Screening of lactic acid bacteria Isolated from Iranian sourdoughs for antifungal activity: Enterococcus faecium showed the most potent antifungal activity in bread. Appl. Food Biotechnol. 2017, 4, 219–227. [Google Scholar]
- Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Baudy-Floc’h, M.; Le Blay, G.; Haertlé, T.; Chobert, J. Antifungal properties of durancins isolated from Enterococcus durans A5-11 and of its synthetic fragments. Lett. Appl. Microbiol. 2013, 56, 237–244. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample Code | Depository | Flour Origin | % NaCl per Backslopping | % Sourdough Used in a Backslopping | No. of Backsloppings | Time and Temperature of Backslopping |
|---|---|---|---|---|---|---|
| 01M | Bakery Art1, Plovdiv | Triticumaestivum, white | 1.5 | 20 | 3 | 24 h/22 °C |
| 02P1 | Bakery The Bread, Plovdiv | Secale cereale, wholegrain | 2.0 | 20 | 3 | 9 h/28 °C |
| 03P2 | Bakery Art 2, Plovdiv | T. aestivum, white | 2.0 | 20 | 3 | 9 h/28 °C |
| 04P3 | Bakery Art 3, Plovdiv | T. aestivum, white | 2.0 | 20 | 3 | 9 h/28 °C |
| 05S | Bakery “8”, Smolyan | T. aestivum, white | 1.5 | 25 | 3 | 18 h/25 °C |
| 06SE | Homemade, Selcha | T. aestivum/Cicer arietinum (90/10%) | 1.5 | 30 | 1 | 18 h/22 °C |
| 07B1 | Bakery Samun 1, Bansko | T. aestivum, white | 1.5 | 25 | 3 | 18 h/25 °C |
| 08B2 | Bakery Samun 2, Bansko | T. aestivum, wholegrain | 1.5 | 25 | 3 | 18 h/22 °C |
| 09B3 | Bakery Samun 3, Bansko | Triticum monococcum, wholegrain | 1.5 | 25 | 3 | 18 h/22 °C |
| 10B4 | Bakery Samun 4, Bansko | Secale cereale, wholegrain | 1.5 | 25 | 3 | 18 h/22 °C |
| 11R1 | Bakery Kusi 1, Ruse | T. aestivum, white | 2.0 | 30 | 7 | 24 h/30 °C |
| 12R2 | Bakery Kusi 2, Ruse | T. aestivum, white | 2.0 | 30 | 7 | 24 h/24 °C |
| Sample | pH | TTA, mL 0.1 NaOH | Dry Matter, % | MRS | MR 5 | M 17 | M 17 G | SA | ME | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TVC, cfu/mL | No. of Isolates | TVC, cfu/mL | No. of Isolates | TVC, cfu/mL | No. of Isolates | TVC, cfu/mL | No. of Isolates | TVC, cfu/mL | No. of Isolates | TVC, cfu/mL | No. of Isolates | ||||
| 01M | 3.78 | 9.4 | 53.10 | 3.2 × 105 | 4 | 6.1 × 105 | 5 | 7.0 × 105 | 5 | 2.1 × 105 | 3 | 4.2 × 108 | 4 | 5.4 × 108 | 3 |
| 02P1 | 3.57 | 11.6 | 36.20 | 9.2 × 105 | 7 | 4.4 × 105 | 5 | 4.3 × 105 | 4 | 7.9 × 105 | 6 | 6.5 × 1010 | 5 | 6.8 × 1010 | 5 |
| 03P2 | 3.84 | 9.8 | 40.30 | 2.6 × 105 | 3 | 2.7 × 105 | 3 | 2.7 × 105 | 3 | 4.8 × 105 | 4 | 6.3 × 109 | 5 | 1.1 × 1010 | 5 |
| 04P3 | 3.60 | 11.4 | 37.14 | 2.8 × 104 | 3 | 2.0 × 105 | 3 | 1.1 × 105 | 7 | 1.1 × 105 | 2 | 6.6 × 1010 | 5 | 7.0 × 1010 | 5 |
| 05S | 3.96 | 10.4 | 58.16 | 3.1 × 1012 | 4 | 2.5 × 1012 | 4 | 4.2 × 1011 | 4 | 2.3 × 1012 | 3 | 5.3 × 109 | 5 | 5.1 × 108 | 5 |
| 06SE | 5.11 | 5.2 | 55.21 | 1.7 × 104 | 8 | 1.4 × 104 | 7 | 3.3 × 105 | 5 | 1.6 × 104 | 7 | 1.0 × 104 | 5 | 2.1 × 104 | 4 |
| 07B1 | 3.61 | 12.0 | 42.02 | 2.7 × 1011 | 3 | 3.6 × 1011 | 4 | 9.9 × 108 | 7 | 9.2 × 109 | 6 | 6.6 × 1010 | 5 | 8.0 × 109 | 4 |
| 08B2 | 4.07 | 8.1 | 39.39 | 1.8 × 1011 | 2 | 3.9 × 1011 | 4 | 2.9 × 1010 | 4 | 8.3 × 1010 | 6 | 3.5 × 109 | 4 | 2.1 × 109 | 4 |
| 09B3 | 4.17 | 8.3 | 43.50 | 2.0 × 1011 | 2 | 1.5 × 1011 | 7 | 9.8 × 1011 | 6 | 2.6 × 1011 | 3 | 6.4 × 109 | 5 | 4.2 × 106 | 4 |
| 10B4 | 3.58 | 12.4 | 33.01 | 9.7 × 1011 | 7 | 9.9 × 1011 | 6 | 8.5 × 108 | 6 | 3.1 × 109 | 4 | 7.4 × 108 | 5 | 1.0 × 1010 | 4 |
| 11R1 | 3.62 | 10.2 | 49.86 | 3.2 × 107 | 4 | 1.8 × 1011 | 2 | 3.5 × 108 | 4 | 1.7 × 108 | 2 | 9.8 × 108 | 5 | 9.4 × 109 | 4 |
| 12R2 | 3.64 | 10.0 | 53.18 | 2.1 × 108 | 2 | 1.0 × 1011 | 8 | 2.6 × 108 | 3 | 3.8 × 108 | 4 | 3.4 × 109 | 2 | 1.9 × 109 | 3 |
| Total isolates from LAB media: 215 | Total yeast isolates: 106 | ||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petkova, M.; Stefanova, P.; Gotcheva, V.; Angelov, A. Isolation and Characterization of Lactic Acid Bacteria and Yeasts from Typical Bulgarian Sourdoughs. Microorganisms 2021, 9, 1346. https://doi.org/10.3390/microorganisms9071346
Petkova M, Stefanova P, Gotcheva V, Angelov A. Isolation and Characterization of Lactic Acid Bacteria and Yeasts from Typical Bulgarian Sourdoughs. Microorganisms. 2021; 9(7):1346. https://doi.org/10.3390/microorganisms9071346
Chicago/Turabian StylePetkova, Mariana, Petya Stefanova, Velitchka Gotcheva, and Angel Angelov. 2021. "Isolation and Characterization of Lactic Acid Bacteria and Yeasts from Typical Bulgarian Sourdoughs" Microorganisms 9, no. 7: 1346. https://doi.org/10.3390/microorganisms9071346
APA StylePetkova, M., Stefanova, P., Gotcheva, V., & Angelov, A. (2021). Isolation and Characterization of Lactic Acid Bacteria and Yeasts from Typical Bulgarian Sourdoughs. Microorganisms, 9(7), 1346. https://doi.org/10.3390/microorganisms9071346