
MPLA and AddaVax® Adjuvants Fail to Promote Intramuscular LaAg Vaccine Protectiveness against Experimental Cutaneous Leishmaniasis

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Parasites
2.3. Immunization by Intramuscular Route
2.4. Infection, Measurement of Lesions and Hypersensitivity
2.5. Parasite Load by Limiting Dilution Assay (LDA)
2.6. Lymphoproliferation
2.7. Flow Cytometry
2.8. Statistics
3. Results
3.1. Intramuscular LaAg Vaccine Induces Partial Protection in the Lesion Size of C57BL/6 Mice Infected with 2 × 105 Promastigotes, But Not with 2 × 106 Promastigotes
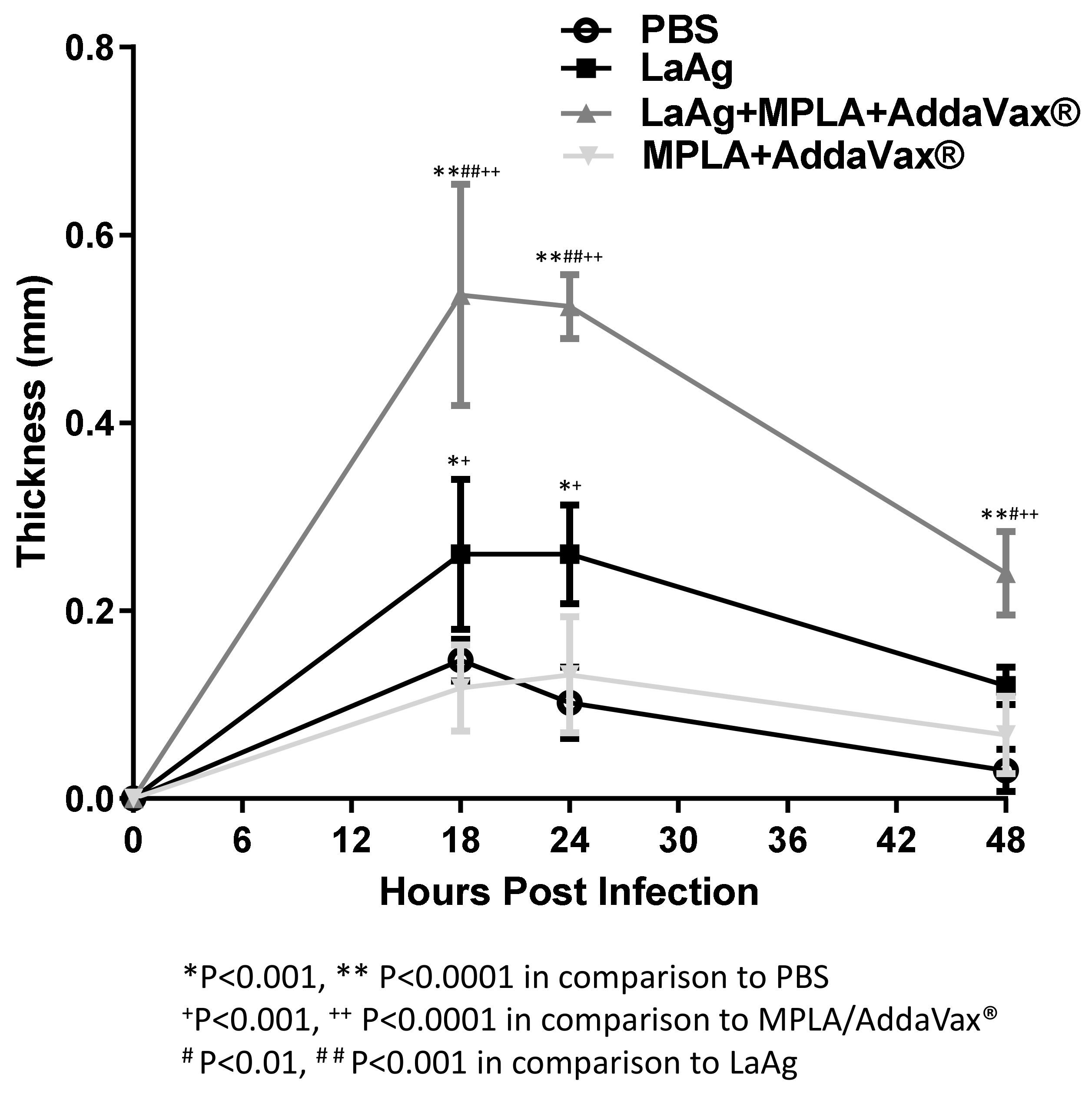
3.2. Intramuscular LaAg Vaccine Induces a Strong Delayed Hypersensitivity Response When Associated with the Adjuvants MPLA/AddaVax®
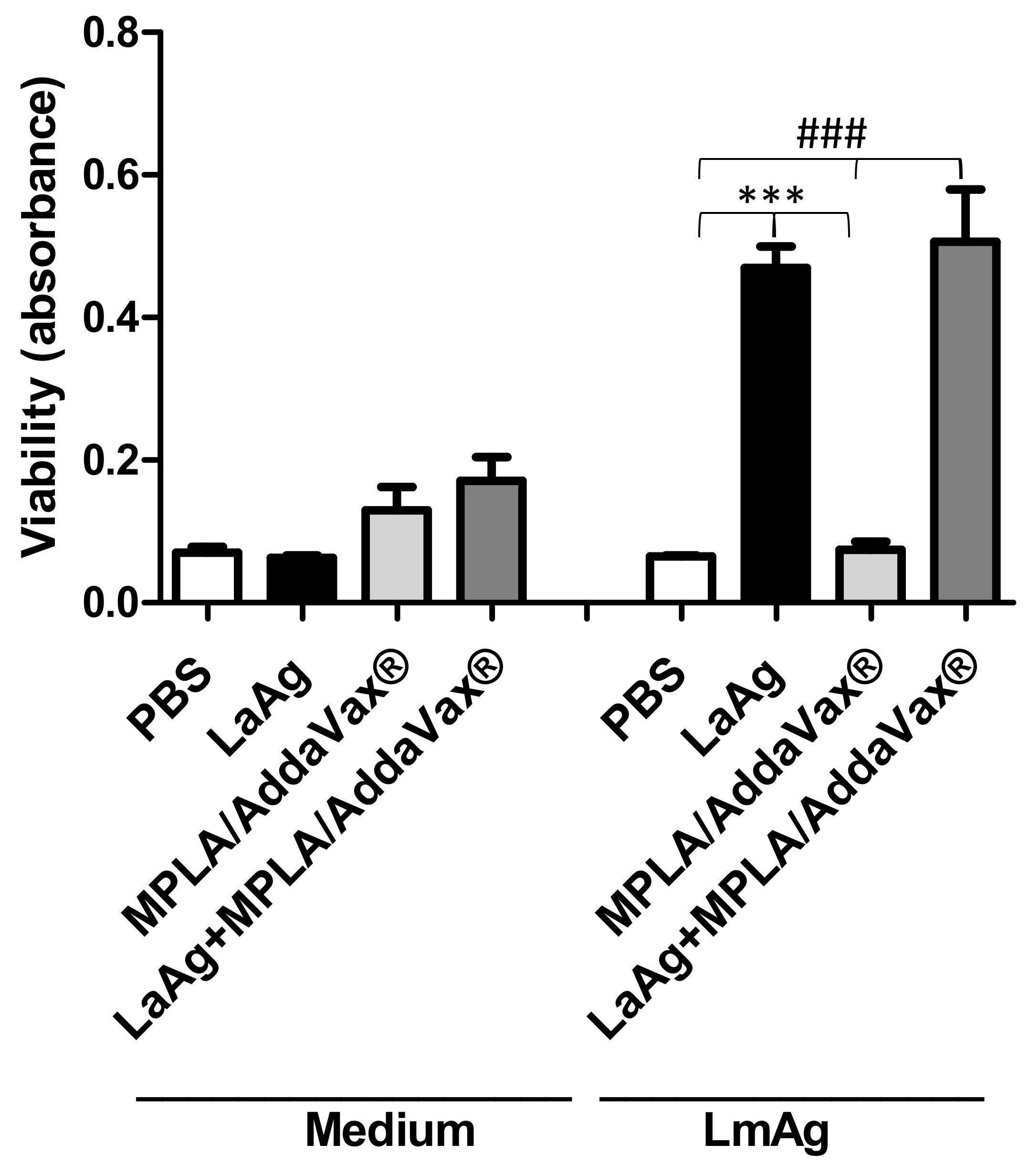
3.3. Immunization with LaAg Alone or LaAg and MPLA/AddaVax® Induces Proliferation of Immune Cells
3.4. LaAg Vaccine Associated with MPLA/AddaVax® Displays the Same Profile As the LaAg Vaccine in C57BL/6 Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Azeredo-Coutinho, R.; Conceicao-Silva, F.; Schubach, A.; Cupolillo, E.; Quintella, L.; Madeira, M.; Pacheco, R.; Valete-Rosalino, C.; Mendonça, S. First report of diffuse cutaneous leishmaniasis and Leishmania amazonensis infection in Rio de Janeiro State, Brazil. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 735–737. [Google Scholar] [CrossRef] [PubMed]
- Kholoud, K.; Bounoua, L.; Sereno, D.; el Hidan, M.; Messouli, M. Emerging and Re-Emerging Leishmaniases in the Mediterranean Area: What Can Be Learned from a Retrospective Review Analysis of the Situation in Morocco during 1990 to 2010? Microorganisms 2020, 8, 1511. [Google Scholar] [CrossRef] [PubMed]
- Sereno, D. Leishmania (Mundinia) spp.: From description to emergence as new human and animal Leishmania pathogens. New Microbes New Infect. 2019, 30, 100540. [Google Scholar] [CrossRef] [PubMed]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.-C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.W.; Mwenechanya, R.; Papadopoulou, B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Negl. Trop. Dis. 2017, 11, e0006052. [Google Scholar] [CrossRef] [PubMed]
- Eddaikra, N.; Ait-Oudhia, K.; Kherrachi, I.; Oury, B.; Moulti-Mati, F.; Benikhlef, R.; Harrat, Z.; Sereno, D. Antimony susceptibility of Leishmania isolates collected over a 30-year period in Algeria. PLoS Negl. Trop. Dis. 2018, 12, e0006310. [Google Scholar] [CrossRef]
- Grimaldi, G.; Tesh, R.B. Leishmaniases of the New World: Current concepts and implications for future research. Clin. Microbiol. Rev. 1993, 6, 230–250. [Google Scholar] [CrossRef]
- Duthie, M.S.; Reed, S.G. Not All Antigens Are Created Equally: Progress, Challenges, and Lessons Associated with Developing a Vaccine for Leishmaniasis. Clin. Vaccine Immunol. 2017, 24, e00108-17. [Google Scholar] [CrossRef]
- Srivastava, S.; Shankar, P.; Mishra, J.; Singh, S. Possibilities and challenges for developing a successful vaccine for leishmaniasis. Parasites Vectors 2016, 9, 277. [Google Scholar] [CrossRef]
- Khamesipour, A.; Dowlati, Y.; Asilian, A.; Hashemi-Fesharki, R.; Javadi, A.; Noazin, S.; Modabber, F. Leishmanization: Use of an old method for evaluation of candidate vaccines against leishmaniasis. Vaccine 2005, 23, 3642–3648. [Google Scholar] [CrossRef]
- Mayrink, W.; da Costa, C.; Magalhães, P.; Melo, M.; Dias, M.; Lima, A.; Michalick, M.; Williams, P. A field trial of a vaccine against American dermal leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 385–387. [Google Scholar] [CrossRef]
- Mayrink, W.; dos Santos, G.C.; de Toledo, V.d.P.C.P.; Guimaraes, T.M.P.D.; Machado-Coelho, G.L.L.; Genaro, O.; da Costa, C.A. Vaccination of C57BL/10 mice against cutaneous leishmaniasis using killed promastigotes of different strains and species of Leishmania. Rev. Soc. Bras. Med. Trop. 2002, 35, 125–132. [Google Scholar] [CrossRef]
- Mayrink, W.; Guimarães, T.; Toledo, V.; da Costa, C.; Vilela, L.; Pinto, J.; Genaro, O. Short report: Evaluation of the potency and stability of a candidate vaccine against American cutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 1999, 61, 294–295. [Google Scholar] [CrossRef] [PubMed]
- Vélez, I.D.; Gilchrist, K.; Arbelaez, M.P.; Rojas, C.A.; Puerta, J.A.; Antunes, C.M.; Zicker, F.; Modabber, F. Failure of a killed Leishmania amazonensis vaccine against American cutaneous leishmaniasis in Colombia. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 593–598. [Google Scholar] [CrossRef]
- Pinheiro, R.O.; Pinto, E.F.; Lopes, J.R.C.; Guedes, H.L.M.; Fentanes, R.F.; Rossi-Bergmann, B. TGF-β-associated enhanced susceptibility to leishmaniasis following intramuscular vaccination of mice with Leishmania amazonensis antigens. Microbes Infect. 2005, 7, 1317–1323. [Google Scholar] [CrossRef]
- Leclerc, C. New approaches in vaccine development. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 329–341. [Google Scholar] [CrossRef]
- Carneiro, M.B.H.; Sousa, L.M.D.A.E.; Vaz, L.G.; dos Santos, L.M.; Vilela, L.; de Souza, C.C.; Gonçalves, R.; Tafuri, W.L.; Afonso, L.C.C.; Côrtes, D.F.; et al. Short-term protection conferred by Leishvacin® against experimental Leishmania amazonensis infection in C57BL/6 mice. Parasitol. Int. 2014, 63, 826–834. [Google Scholar] [CrossRef]
- Kenney, R.T.; Sacks, D.L.; Sypek, J.P.; Vilela, L.; Gam, A.; Evans-Davis, K. Protective immunity using recombinant human IL-12 and alum as adjuvants in a primate model of cutaneous leishmaniasis. J. Immunol. 1999, 163, 4481–4488. [Google Scholar]
- Botelho, A.; Tafuri, W.; Genaro, O.; Mayrink, W. Histopathology of human American cutaneous leishmaniasis before and after treatment. Rev. Soc. Bras. Med. Trop. 1998, 31, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hernandezs, M.; Barcante, T.; Vilela, L.; Tafuri, W.; Afonso, L.C.C.; Vieira, L. Vaccine-induced protection against Leishmania amazonensis is obtained in the absence of IL-12/23p40. Immunol. Lett. 2006, 105, 38–47. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aebischer, T.; Wolfram, M.; Patzer, S.I.; Ilg, T.; Wiese, M.; Overath, P. Subunit Vaccination of Mice against New World Cutaneous Leishmaniasis: Comparison of Three Proteins Expressed in Amastigotes and Six Adjuvants. Infect. Immun. 2000, 68, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Ott, G.; Radhakrishnan, R.; Fang, J.-H.; Hora, M.; O’Hagan, D.T. The Adjuvant MF59: A 10-Year Perspective. Vaccine Adjuv. 2003, 42, 211–228. [Google Scholar] [CrossRef]
- Calabro, S.; Tritto, E.; Pezzotti, A.; Taccone, M.; Muzzi, A.; Bertholet, S.; de Gregorio, E.; O’Hagan, D.T.; Baudner, B.; Seubert, A. The adjuvant effect of MF59 is due to the oil-in-water emulsion formulation, none of the individual components induce a comparable adjuvant effect. Vaccine 2013, 31, 3363–3369. [Google Scholar] [CrossRef]
- Ott, G.; Barchfeld, G.L.; Chernoff, D.; Radhakrishnan, R.; van Hoogevest, P.; Van Nest, G. MF59 Design and evaluation of a safe and potent adjuvant for human vaccines. Pharm Biotechnol. 1995, 6, 277–296. [Google Scholar] [CrossRef] [PubMed]
- Coler, R.N.; Reed, S.G. Second-generation vaccines against leishmaniasis. Trends Parasitol. 2005, 21, 244–249. [Google Scholar] [CrossRef]
- Coler, R.N.; Skeiky, Y.A.W.; Bernards, K.; Greeson, K.; Carter, D.; Cornellison, C.D.; Modabber, F.; Campos-Neto, A.; Reed, S.G. Immunization with a Polyprotein Vaccine Consisting of the T-Cell Antigens Thiol-Specific Antioxidant, Leishmania major Stress-Inducible Protein 1, and Leishmania elongation Initiation Factor Protects against Leishmaniasis. Infect. Immun. 2002, 70, 4215–4225. [Google Scholar] [CrossRef] [PubMed]
- Bertholet, S.; Goto, Y.; Carter, L.; Bhatia, A.; Howard, R.F.; Carter, D.; Coler, R.N.; Vedvick, T.S.; Reed, S.G. Optimized subunit vaccine protects against experimental leishmaniasis. Vaccine 2009, 27, 7036–7045. [Google Scholar] [CrossRef]
- Pratti, J.E.S.; Ramos, T.D.; Pereira, J.C.; da Fonseca-Martins, A.M.; Maciel-Oliveira, D.; Oliveira-Silva, G.; Mello, M.F.d.; Chaves, S.P.; Gomes, D.C.O.; Diaz, B.L.; et al. Efficacy of intranasal LaAg vaccine against Leishmania amazonensis infection in partially resistant C57Bl/6 mice. Parasites Vectors 2016, 9, 1–11. [Google Scholar] [CrossRef]
- Pinto, E.F.; Pinheiro, R.O.; Rayol, A.; Larraga, V.; Rossi-Bergmann, B. Intranasal Vaccination against Cutaneous Leishmaniasis with a Particulated Leishmanial Antigen or DNA Encoding LACK. Infect. Immun. 2004, 72, 4521–4527. [Google Scholar] [CrossRef] [PubMed]
- Global Vaccine Safety Initiative. Report of a Meeting, Santiago, Chile, 8–9 October 2018; World Health Organization: Geneva, Switzerland, 2019; p. 89. [Google Scholar]
- J.A.X. USA. Why Mice for Biomedical Research? Available online: https://www.jax.org/about-us/why-mice (accessed on 6 January 2020).
- Côrtes, D.F.; Carneiro, M.B.H.; Santos, L.M.; Souza, T.C.D.O.; Maioli, T.U.; Duz, A.L.C.; Ramos-Jorge, M.L.; Afonso, L.C.C.; Carneiro, C.; Vieira, L.Q. Low and high-dose intradermal infection with Leishmania major and Leishmania amazonensis in C57BL/6 mice. Memórias Inst. Oswaldo Cruz 2010, 105, 736–745. [Google Scholar] [CrossRef]
- Dos-Santos, J.S.; Firmino-Cruz, L.; Ramos, T.D.; da Fonseca-Martins, A.M.; Oliveira-Maciel, D.; De-Medeiros, J.V.R.; Chaves, S.P.; Gomes, D.C.O.; Guedes, H.L.D.M. Characterization of Sv129 Mice as a Susceptible Model to Leishmania amazonensis. Front. Med. 2019, 6, 1–12. [Google Scholar] [CrossRef]
- Guedes, H.L.D.M.; Pinheiro, R.O.; Chaves, S.P.; De-Simone, S.G.; Rossi-Bergmann, B. Serine proteases of Leishmania amazonensis as immunomodulatory and disease-aggravating components of the crude LaAg vaccine. Vaccine 2010, 28, 5491–5496. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Petrovsky, N.; Aguilar, J.C. Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol. 2004, 82, 488–496. [Google Scholar] [CrossRef]
- De Luca, P.M.; Mayrink, W.; Alves, C.R.; Coutinho, S.G.; Oliveira, M.P.; Bertho, A.L.; Toledo, V.P.; Costa, C.A.; Genaro, O.; Mendonça, S.C. Evaluation of the stability and immunogenicity of autoclaved and nonautoclaved preparations of a vaccine against American tegumentary leishmaniasis. Vaccine 1999, 17, 1179–1185. [Google Scholar] [CrossRef]
- Coler, R.N.; Goto, Y.; Bogatzki, L.; Raman, V.; Reed, S.G. Leish-111f, a Recombinant Polyprotein Vaccine That Protects against Visceral Leishmaniasis by Elicitation of CD4+ T Cells. Infect. Immun. 2007, 75, 4648–4654. [Google Scholar] [CrossRef]
- Black, C.A. Delayed type hypersensitivity: Current theories with an historic perspective. Dermatol. Online J. 1999, 5, 5–7. [Google Scholar]
- Richerson, H.B.; Dvorak, H.F.; Leskowitz, S. Cutaneous Basophil Hypersensitivity. J. Exp. Med. 1970, 132, 546–557. [Google Scholar] [CrossRef]
- Manzur, A.; Bari, A.U. Sensitivity of leishmanin skin test in patients of acute cutaneous leishmaniasis. Dermatol. Online J. 2006, 12, 12. [Google Scholar]
- Bacellar, O.; Lessa, H.; Schriefer, A.; Machado, P.; de Jesus, A.R.; Dutra, W.O.; Gollob, K.J.; Carvalho, E.M. Up-Regulation of Th1-Type Responses in Mucosal Leishmaniasis Patients. Infect. Immun. 2002, 70, 6734–6740. [Google Scholar] [CrossRef]
- Carvalho, E.M.; Bacellar, O.; Brownell, C.; Regis, T.; Coffman, R.L.; Reed, S.G. Restoration of IFN-gamma production and lymphocyte proliferation in visceral leishmaniasis. J. Immunol. 1994, 152, 5949–5956. [Google Scholar] [PubMed]
- Vélez, I.D.; Gilchrist, K.; Martínez, S.; Ramírez-Pineda, J.R.; Ashman, J.A.; Alves, F.P.; Coler, R.N.; Bogatzki, L.Y.; Kahn, S.J.; Beckmann, A.M.; et al. Safety and immunogenicity of a defined vaccine for the prevention of cutaneous leishmaniasis. Vaccine 2009, 28, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, R.O.; Pinto, E.F.; Benedito, A.B.; Lopes, U.G.; Rossi-Bergmann, B. The T-cell anergy induced by Leishmania amazonensis antigens is related with defective antigen presentation and apoptosis. An. Acad. Bras. Ciências 2004, 76, 519–527. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Germanó, M.J.; Lozano, E.S.; Sanchez, M.V.; Bruna, F.A.; García-Bustos, M.F.; Lochedino, A.L.S.; Salomón, M.C.; Fernandes, A.P.; Mackern-Oberti, J.P.; Cargnelutti, D.E. Evaluation of different total Leishmania amazonensis antigens for the development of a first-generation vaccine formulated with a Toll-like receptor-3 agonist to prevent cutaneous leishmaniasis. Memórias Inst. Oswaldo Cruz 2020, 115, e200067. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira-Maciel, D.; dos-Santos, J.S.; Oliveira-Silva, G.; Mello, M.F.d.; da Fonseca-Martins, A.M.; Carneiro, M.P.D.; Ramos, T.D.; Firmino-Cruz, L.; Gomes, D.C.O.; Rossi-Bergmann, B.; et al. MPLA and AddaVax® Adjuvants Fail to Promote Intramuscular LaAg Vaccine Protectiveness against Experimental Cutaneous Leishmaniasis. Microorganisms 2021, 9, 1272. https://doi.org/10.3390/microorganisms9061272
Oliveira-Maciel D, dos-Santos JS, Oliveira-Silva G, Mello MFd, da Fonseca-Martins AM, Carneiro MPD, Ramos TD, Firmino-Cruz L, Gomes DCO, Rossi-Bergmann B, et al. MPLA and AddaVax® Adjuvants Fail to Promote Intramuscular LaAg Vaccine Protectiveness against Experimental Cutaneous Leishmaniasis. Microorganisms. 2021; 9(6):1272. https://doi.org/10.3390/microorganisms9061272
Chicago/Turabian StyleOliveira-Maciel, Diogo, Júlio Souza dos-Santos, Gabriel Oliveira-Silva, Mirian França de Mello, Alessandra Marcia da Fonseca-Martins, Monique Pacheco Duarte Carneiro, Tadeu Diniz Ramos, Luan Firmino-Cruz, Daniel Claudio Oliveira Gomes, Bartira Rossi-Bergmann, and et al. 2021. "MPLA and AddaVax® Adjuvants Fail to Promote Intramuscular LaAg Vaccine Protectiveness against Experimental Cutaneous Leishmaniasis" Microorganisms 9, no. 6: 1272. https://doi.org/10.3390/microorganisms9061272
APA StyleOliveira-Maciel, D., dos-Santos, J. S., Oliveira-Silva, G., Mello, M. F. d., da Fonseca-Martins, A. M., Carneiro, M. P. D., Ramos, T. D., Firmino-Cruz, L., Gomes, D. C. O., Rossi-Bergmann, B., & de Matos Guedes, H. L. (2021). MPLA and AddaVax® Adjuvants Fail to Promote Intramuscular LaAg Vaccine Protectiveness against Experimental Cutaneous Leishmaniasis. Microorganisms, 9(6), 1272. https://doi.org/10.3390/microorganisms9061272