
Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review


Abstract
1. Introduction
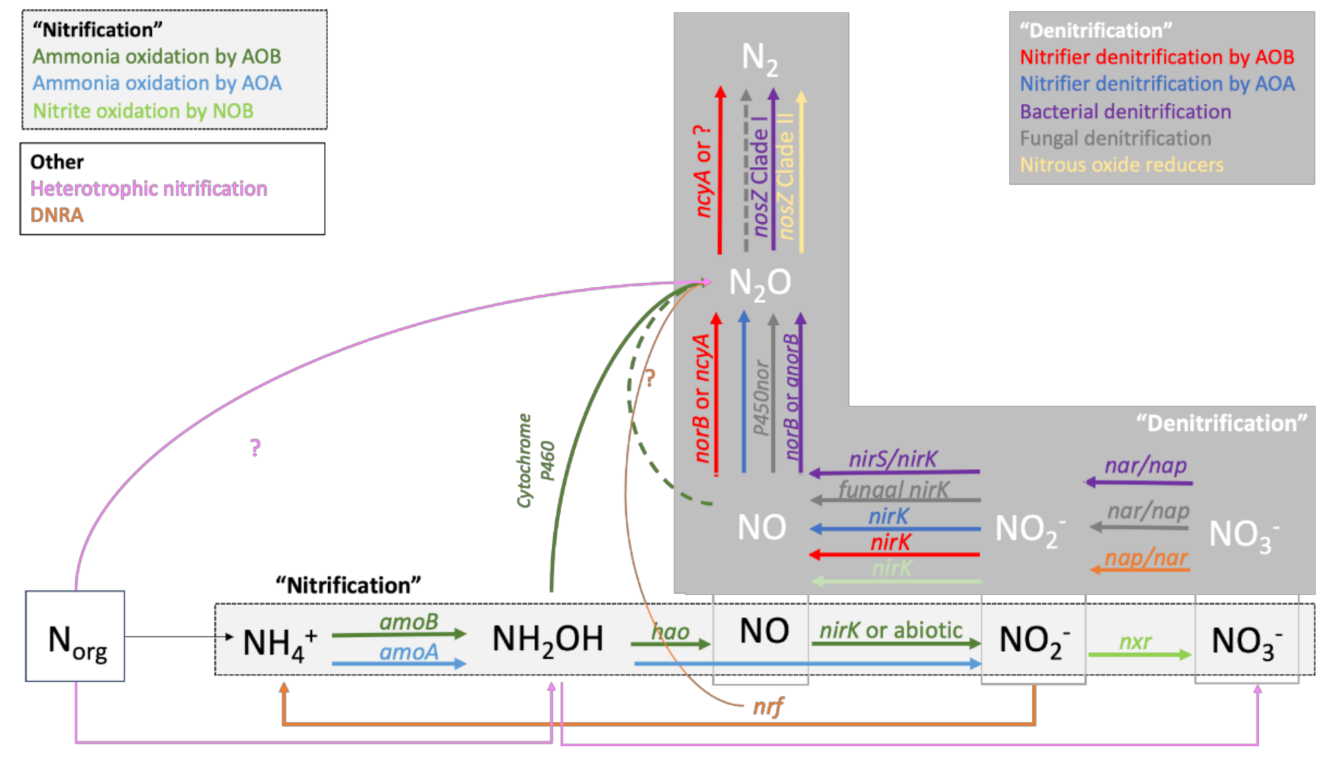
2. The N Cycle and Mechanisms Responsible for the Production and Consumption of N2O in Soils
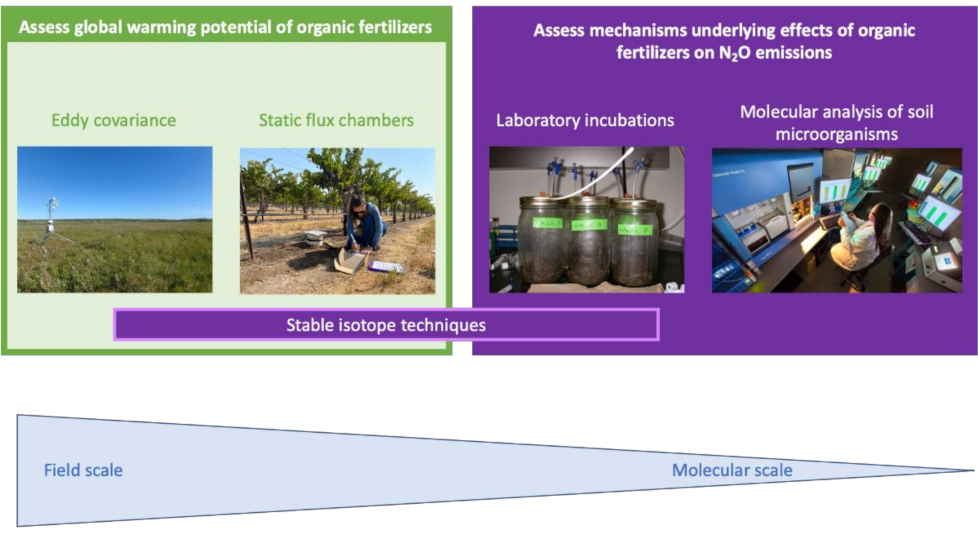
3. Methods to Study N2O Emissions and Underlying Processes in Soils
4. Sources of Organic Fertilizers and N2O Emissions
5. Effects of Organic Fertilizers on Nitrifying and Denitrifying Soil Microorganisms
5.1. Effects of Raw Manure on the Structure and Activity of the Nitrifying and Denitrifying Community
5.2. Effects of Processed Manure on the Structure and Activity of the Nitrifying and Denitrifying Communities
5.3. Management Strategies to Reduce N2O Emissions from Soils Amended with Manure-Based Organic Fertilizers
6. Conclusions, Future Directions and Research Needs
Author Contributions
Funding
Conflicts of Interest
References
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization Mechanisms of Soil Organic Matter: Implications for C-Saturation of Soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Yan, X.; Gong, W. The Role of Chemical and Organic Fertilizers on Yield, Yield Variability and Carbon Sequestration—Results of a 19-Year Experiment. Plant Soil 2010, 331, 471–480. [Google Scholar] [CrossRef]
- Tautges, N.E.; Chiartas, J.L.; Gaudin, A.C.M.; O’Geen, A.T.; Herrera, I.; Scow, K.M. Deep Soil Inventories Reveal That Impacts of Cover Crops and Compost on Soil Carbon Sequestration Differ in Surface and Subsurface Soils. Glob. Chang. Biol. 2019, 25, 3753–3766. [Google Scholar] [CrossRef]
- Brar, B.S.; Singh, K.; Dheri, G.S.; Balwinder-Kumar. Carbon Sequestration and Soil Carbon Pools in a Rice? Wheat Cropping System: Effect of Long-Term Use of Inorganic Fertilizers and Organic Manure. Soil Tillage Res. 2013, 128, 30–36. [Google Scholar] [CrossRef]
- Martínez-Blanco, J.; Lazcano, C.; Christensen, T.H.; Muñoz, P.; Rieradevall, J.; Møller, J.; Antón, A.; Boldrin, A. Compost Benefits for Agriculture Evaluated by Life Cycle Assessment. A Review. Agron. Sustain. Dev. 2013, 33, 721–732. [Google Scholar] [CrossRef]
- Lazcano, C.; Gómez-Brandón, M.; Revilla, P.; Domínguez, J. Short-Term Effects of Organic and Inorganic Fertilizers on Soil Microbial Community Structure and Function: A Field Study with Sweet Corn. Biol. Fertil. Soils 2013, 49, 723–733. [Google Scholar] [CrossRef]
- Ling, N.; Zhu, C.; Xue, C.; Chen, H.; Duan, Y.; Peng, C.; Guo, S.; Shen, Q. Insight into How Organic Amendments Can Shape the Soil Microbiome in Long-Term Field Experiments as Revealed by Network Analysis. Soil Biol. Biochem. 2016, 99, 137–149. [Google Scholar] [CrossRef]
- García-Gil, J.C.; Plaza, C.; Soler-Rovira, P.; Polo, A. Long-Term Effects of Municipal Solid Waste Compost Application on Soil Enzyme Activities and Microbial Biomass. Soil Biol. Biochem. 2000, 32, 1907–1913. [Google Scholar] [CrossRef]
- Sokol, N.W.; Bradford, M.A. Microbial Formation of Stable Soil Carbon Is More Efficient from Belowground than Aboveground Input. Nat. Geosci. 2019, 12, 46–53. [Google Scholar] [CrossRef]
- Ryals, R.; Silver, W.L. Effects of Organic Matter Amendments on Net Primary Productivity and Greenhouse Gas Emissions in Annual Grasslands. Ecol. Appl. 2013, 23, 46–59. [Google Scholar] [CrossRef] [PubMed]
- DeLonge, M.S.; Ryals, R.; Silver, W.L. A Lifecycle Model to Evaluate Carbon Sequestration Potential and Greenhouse Gas Dynamics of Managed Grasslands. Ecosystems 2013, 16, 962–979. [Google Scholar] [CrossRef]
- Lazcano, C.; Tsang, A.; Doane, T.A.; Pettygrove, G.S.; Horwath, W.R.; Burger, M. Soil Nitrous Oxide Emissions in Forage Systems Fertilized with Liquid Dairy Manure and Inorganic Fertilizers. Agric. Ecosyst. Environ. 2016, 225, 160–172. [Google Scholar] [CrossRef]
- Velthof, G.L.; Kuikman, P.J.; Oenema, O. Nitrous Oxide Emission from Animal Manures Applied to Soil under Controlled Conditions. Biol. Fertil. Soils 2003, 37, 221–230. [Google Scholar] [CrossRef]
- Lugato, E.; Leip, A.; Jones, A. Mitigation Potential of Soil Carbon Management Overestimated by Neglecting N2O Emissions. Nat. Clim. Chang. 2018, 8, 219–223. [Google Scholar] [CrossRef]
- Guenet, B.; Gabrielle, B.; Chenu, C.; Arrouays, D.; Balesdent, J.; Bernoux, M.; Bruni, E.; Caliman, J.; Cardinael, R.; Chen, S.; et al. Can N2O Emissions Offset the Benefits from Soil Organic Carbon Storage? Glob. Chang. Biol. 2020, 27, 237–256. [Google Scholar] [CrossRef]
- IPCC Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013.
- van Groenigen, J.W.; Huygens, D.; Boeckx, P.; Kuyper, T.W.; Lubbers, I.M.; Rütting, T.; Groffman, P.M. The Soil N Cycle: New Insights and Key Challenges. SOIL 2015, 1, 235–256. [Google Scholar] [CrossRef]
- Syakila, A.; Kroeze, C. The Global Nitrous Oxide Budget Revisited. Greenh. Gas Meas. Manag. 2011, 1, 17–26. [Google Scholar] [CrossRef]
- Lowrance, R.; Johnson, J.C.; Newton, G.L.; Williams, R.G. Denitrification from Soils of a Year-Round Forage Production System Fertilized with Liquid Dairy Manure. J. Environ. Qual. 1998, 27, 1504–1511. [Google Scholar] [CrossRef]
- Zhu, X.; Silva, L.C.R.; Doane, T.A.; Wu, N.; Horwath, W.R. Quantifying the Effects of Green Waste Compost Application, Water Content and Nitrogen Fertilization on Nitrous Oxide Emissions in 10 Agricultural Soils. J. Environ. Qual. 2013, 42, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, V.; de los Valdez-Perez, M.A.; Luna-Guido, M.; Ceballos-Ramirez, J.M.; Franco-Hernández, O.; van Cleemput, O.; Marsch, R.; Thalasso, F.; Dendooven, L. Emission of Nitrous Oxide and Carbon Dioxide and Dynamics of Mineral N in Wastewater Sludge, Vermicompost or Inorganic Fertilizer Amended Soil at Different Water Contents: A Laboratory Study. Appl. Soil Ecol. 2011, 49, 263–267. [Google Scholar] [CrossRef]
- Zhang, J.; Müller, C.; Cai, Z. Heterotrophic Nitrification of Organic N and Its Contribution to Nitrous Oxide Emissions in Soils. Soil Biol. Biochem. 2015, 84, 199–209. [Google Scholar] [CrossRef]
- Bouwman, A.F.; Boumans, L.J.M.; Batjes, N.H. Modeling Global Annual N2O and NO Emissions from Fertilized Fields: N2O and no Emissions from Fertilizers. Glob. Biogeochem. Cycles 2002, 16, 28-1–28-9. [Google Scholar] [CrossRef]
- Vallejo, A.; Skiba, U.; Garciatorres, L.; Arce, A.; Lopezfernandez, S.; Sanchezmartin, L. Nitrogen Oxides Emission from Soils Bearing a Potato Crop as Influenced by Fertilization with Treated Pig Slurries and Composts. Soil Biol. Biochem. 2006, 38, 2782–2793. [Google Scholar] [CrossRef]
- Chen, H.; Li, X.; Hu, F.; Shi, W. Soil Nitrous Oxide Emissions Following Crop Residue Addition: A Meta-Analysis. Glob. Chang. Biol. 2013, 19, 2956–2964. [Google Scholar] [CrossRef]
- Zhu-Barker, X.; Doane, T.A.; Horwath, W.R. Role of Green Waste Compost in the Production of N2O from Agricultural Soils. Soil Biol. Biochem. 2015, 83, 57–65. [Google Scholar] [CrossRef]
- Davidson, E.A.; Keller, M.; Erickson, H.E.; Verchot, L.V.; Veldkamp, E. Testing a Conceptual Model of Soil Emissions of Nitrous and Nitric Oxides. BioScience 2000, 50, 667. [Google Scholar] [CrossRef]
- Davidson, E.A.; Verchot, L.V. Testing the Hole-in-the-Pipe Model of Nitric and Nitrous Oxide Emissions from Soils Using the TRAGNET Database. Glob. Biogeochem. Cycles 2000, 14, 1035–1043. [Google Scholar] [CrossRef]
- Charles, A.; Rochette, P.; Whalen, J.K.; Angers, D.A.; Chantigny, M.H.; Bertrand, N. Global Nitrous Oxide Emission Factors from Agricultural Soils after Addition of Organic Amendments: A Meta-Analysis. Agric. Ecosyst. Environ. 2017, 236, 88–98. [Google Scholar] [CrossRef]
- Braker, G.; Conrad, R. Diversity, Structure, and Size of N2O-Producing Microbial Communities in Soils—What Matters for Their Functioning? In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 75, ISBN 978-0-12-387046-9. [Google Scholar]
- Ouyang, Y.; Norton, J.M. Short-Term Nitrogen Fertilization Affects Microbial Community Composition and Nitrogen Mineralization Functions in an Agricultural Soil. Appl. Environ. Microbiol. 2019, 86, e02278-19. [Google Scholar] [CrossRef] [PubMed]
- Firestone, M.K.; Davidson, E.A. Microbiological basis of NO and N2O production and consumption in soil. In Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous Oxide Emissions from Soils: How Well Do We Understand the Processes and Their Controls? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Prosser, J.I.; Hink, L.; Gubry-Rangin, C.; Nicol, G.W. Nitrous Oxide Production by Ammonia Oxidizers: Physiological Diversity, Niche Differentiation and Potential Mitigation Strategies. Glob. Chang. Biol. 2020, 26, 103–118. [Google Scholar] [CrossRef]
- Caranto, J.D.; Vilbert, A.C.; Lancaster, K.M. Nitrosomonas europaea Cytochrome P460 Is a Direct Link between Nitrification and Nitrous Oxide Emission. Proc. Natl. Acad. Sci. USA 2016, 113, 14704–14709. [Google Scholar] [CrossRef] [PubMed]
- Knowles, R. Denitrification. Microbiol. Rev. 1982, 46, 43–70. [Google Scholar] [CrossRef]
- Miller, M.N.; Zebarth, B.J.; Dandie, C.E.; Burton, D.L.; Goyer, C.; Trevors, J.T. Influence of Liquid Manure on Soil Denitrifier Abundance, Denitrification, and Nitrous Oxide Emissions. Soil Sci. Soc. Am. J. 2009, 73, 760–768. [Google Scholar] [CrossRef]
- Hu, H.-W.; Chen, D.; He, J.-Z. Microbial Regulation of Terrestrial Nitrous Oxide Formation: Understanding the Biological Pathways for Prediction of Emission Rates. FEMS Microbiol. Rev. 2015, 39, 729–749. [Google Scholar] [CrossRef]
- Thomson, A.J.; Giannopoulos, G.; Pretty, J.; Baggs, E.M.; Richardson, D.J. Biological Sources and Sinks of Nitrous Oxide and Strategies to Mitigate Emissions. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1157–1168. [Google Scholar] [CrossRef]
- ŠImek, M.; Cooper, J.E. The Influence of Soil PH on Denitrification: Progress towards the Understanding of This Interaction over the Last 50 Years: Soil PH and Denitrification. Eur. J. Soil Sci. 2002, 53, 345–354. [Google Scholar] [CrossRef]
- Liu, B.; Mørkved, P.T.; Frostegård, Å.; Bakken, L.R. Denitrification Gene Pools, Transcription and Kinetics of NO, N2O and N2 Production as Affected by Soil PH: Gene Transcription, Gas Kinetics as Affected by Soil PH. FEMS Microbiol. Ecol. 2010, 72, 407–417. [Google Scholar] [CrossRef]
- Hallin, S.; Philippot, L.; Löffler, F.E.; Sanford, R.A.; Jones, C.M. Genomics and Ecology of Novel N2O-Reducing Microorganisms. Trends Microbiol. 2018, 26, 43–55. [Google Scholar] [CrossRef]
- Maeda, K.; Spor, A.; Edel-Hermann, V.; Heraud, C.; Breuil, M.-C.; Bizouard, F.; Toyoda, S.; Yoshida, N.; Steinberg, C.; Philippot, L. N2O Production, a Widespread Trait in Fungi. Sci. Rep. 2015, 5, 9697. [Google Scholar] [CrossRef] [PubMed]
- Wrage-Mönnig, N.; Horn, M.A.; Well, R.; Müller, C.; Velthof, G.; Oenema, O. The Role of Nitrifier Denitrification in the Production of Nitrous Oxide Revisited. Soil Biol. Biochem. 2018, 123, A3–A16. [Google Scholar] [CrossRef]
- Wunderlin, P.; Mohn, J.; Joss, A.; Emmenegger, L.; Siegrist, H. Mechanisms of N2O Production in Biological Wastewater Treatment under Nitrifying and Denitrifying Conditions. Water Res. 2012, 46, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Domeignoz-Horta, L.A.; Putz, M.; Spor, A.; Bru, D.; Breuil, M.C.; Hallin, S.; Philippot, L. Non-Denitrifying Nitrous Oxide-Reducing Bacteria—An Effective N2O Sink in Soil. Soil Biol. Biochem. 2016, 103, 376–379. [Google Scholar] [CrossRef]
- Hénault, C.; Grossel, A.; Mary, B.; Roussel, M.; Léonard, J. Nitrous Oxide Emission by Agricultural Soils: A Review of Spatial and Temporal Variability for Mitigation. Pedosphere 2012, 22, 426–433. [Google Scholar] [CrossRef]
- Eugster, W.; Merbold, L. Eddy Covariance for Quantifying Trace Gas Fluxes from Soils. SOIL 2015, 1, 187–205. [Google Scholar] [CrossRef]
- Parkin, T.B.; Venterea, R.T. USDA-ARS GRACEnet Project Protocols, Chapter 3. Chamber-Based Trace Gas Flux Measurements. In Sampling Protocols; Follett, R.F., Ed.; USDA-ARS: Beltsville, MD, USA, 2010; pp. 1–39. [Google Scholar]
- Zhu, X.; Burger, M.; Doane, T.A.; Horwath, W.R. Ammonia Oxidation Pathways and Nitrifier Denitrification Are Significant Sources of N2O and NO under Low Oxygen Availability. Proc. Natl. Acad. Sci. USA 2013, 110, 6328–6333. [Google Scholar] [CrossRef]
- Bateman, E.J.; Baggs, E.M. Contributions of Nitrification and Denitrification to N2O Emissions from Soils at Different Water-Filled Pore Space. Biol. Fertil. Soils 2005, 41, 379–388. [Google Scholar] [CrossRef]
- Qin, S.; Yuan, H.; Dong, W.; Hu, C.; Oenema, O.; Zhang, Y. Relationship between Soil Properties and the Bias of N2O Reduction by Acetylene Inhibition Technique for Analyzing Soil Denitrification Potential. Soil Biol. Biochem. 2013, 66, 182–187. [Google Scholar] [CrossRef]
- Decock, C.; Six, J. How Reliable Is the Intramolecular Distribution of 15N in N2O to Source Partition N2O Emitted from Soil? Soil Biol. Biochem. 2013, 65, 114–127. [Google Scholar] [CrossRef]
- Baggs, E.M. A Review of Stable Isotope Techniques for N2O Source Partitioning in Soils: Recent Progress, Remaining Challenges and Future Considerations. Rapid Commun. Mass Spectrom. 2008, 22, 1664–1672. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; Vilgalys, R.J. Quantitative Analyses of Nitrogen Cycling Genes in Soils. Pedobiologia 2005, 49, 665–672. [Google Scholar] [CrossRef]
- Chu, H.; Fujii, T.; Morimoto, S.; Lin, X.; Yagi, K.; Hu, J.; Zhang, J. Community Structure of Ammonia-Oxidizing Bacteria under Long-Term Application of Mineral Fertilizer and Organic Manure in a Sandy Loam Soil. Appl. Environ. Microbiol. 2007, 73, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Mintie, A.T.; Heichen, R.S.; Cromack, K.; Myrold, D.D.; Bottomley, P.J. Ammonia-Oxidizing Bacteria along Meadow-to-Forest Transects in the Oregon Cascade Mountains. Appl. Environ. Microbiol. 2003, 69, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-D.; Wells, G.F.; Bae, H.; Criddle, C.S.; Francis, C.A. Occurrence of Ammonia-Oxidizing Archaea in Wastewater Treatment Plant Bioreactors. Appl. Environ. Microbiol. 2006, 72, 5643–5647. [Google Scholar] [CrossRef]
- Ramos, T.M.; Jay-Russell, M.T.; Millner, P.D.; Shade, J.; Misiewicz, T.; Sorge, U.S.; Hutchinson, M.; Lilley, J.; Pires, A.F.A. Assessment of Biological Soil Amendments of Animal Origin Use, Research Needs, and Extension Opportunities in Organic Production. Front. Sustain. Food Syst. 2019, 3, 73. [Google Scholar] [CrossRef]
- Lupton, S. Markets for Waste and Waste–Derived Fertilizers. An Empirical Survey. J. Rural Stud. 2017, 55, 83–99. [Google Scholar] [CrossRef]
- Thangarajan, R.; Bolan, N.S.; Tian, G.; Naidu, R.; Kunhikrishnan, A. Role of Organic Amendment Application on Greenhouse Gas Emission from Soil. Sci. Total Environ. 2013, 465, 72–96. [Google Scholar] [CrossRef]
- Potter, P.; Ramankutty, N.; Bennett, E.M.; Donner, S.D. Characterizing the Spatial Patterns of Global Fertilizer Application and Manure Production. Earth Interact. 2010, 14, 1–22. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; Bai, Z.; Ma, L.; Oenema, O. Global Animal Production and Nitrogen and Phosphorus Flows. Soil Res. 2017, 55, 451. [Google Scholar] [CrossRef]
- Oenema, O.; Wrage, N.; Velthof, G.L.; van Groenigen, J.W.; Dolfing, J.; Kuikman, P.J. Trends in Global Nitrous Oxide Emissions from Animal Production Systems. Nutr. Cycl. Agroecosyst. 2005, 72, 51–65. [Google Scholar] [CrossRef]
- Whalen, J.K.; Thomas, B.W.; Sharifi, M. Novel Practices and Smart Technologies to Maximize the Nitrogen Fertilizer Value of Manure for Crop Production in Cold Humid Temperate Regions. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2019; Volume 153, pp. 1–85. ISBN 978-0-12-817404-3. [Google Scholar]
- Xia, F.; Mei, K.; Xu, Y.; Zhang, C.; Dahlgren, R.A.; Zhang, M. Response of N2O Emission to Manure Application in Field Trials of Agricultural Soils across the Globe. Sci. Total Environ. 2020, 733, 139390. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, Y.; Liu, J.; Gao, J.; Zhang, Y.; Ni, J.; Xie, D.; Wang, Z.; Gao, M. Partial Substitution of Chemical Fertilizer by Organic Materials Changed the Abundance, Diversity, and Activity of NirS-Type Denitrifying Bacterial Communities in a Vegetable Soil. Appl. Soil Ecol. 2020, 152, 103589. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Luo, J.; Di, H.J.; Liu, D.; Fan, J.; Ding, W. Nitrosospira Cluster 8a Plays a Predominant Role in the Nitrification Process of a Subtropical Ultisol under Long-Term Inorganic and Organic Fertilization. Appl. Environ. Microbiol. 2018, 84, e01031-18. [Google Scholar] [CrossRef]
- Snider, D.M.; Wagner-Riddle, C.; Spoelstra, J. Stable Isotopes Reveal Rapid Cycling of Soil Nitrogen after Manure Application. J. Environ. Qual. 2017, 46, 261–271. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, X.; Ju, X. Linkage between N2O Emission and Functional Gene Abundance in an Intensively Managed Calcareous Fluvo-Aquic Soil. Sci. Rep. 2017, 7, 43283. [Google Scholar] [CrossRef] [PubMed]
- Hallin, S.; Jones, C.M.; Schloter, M.; Philippot, L. Relationship between N-Cycling Communities and Ecosystem Functioning in a 50-Year-Old Fertilization Experiment. ISME J. 2009, 3, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Cui, P.; Fan, F.; Yin, C.; Song, A.; Huang, P.; Tang, Y.; Zhu, P.; Peng, C.; Li, T.; Wakelin, S.A.; et al. Long-Term Organic and Inorganic Fertilization Alters Temperature Sensitivity of Potential N2O Emissions and Associated Microbes. Soil Biol. Biochem. 2016, 93, 131–141. [Google Scholar] [CrossRef]
- Surey, R.; Lippold, E.; Heilek, S.; Sauheitl, L.; Henjes, S.; Horn, M.A.; Mueller, C.W.; Merbach, I.; Kaiser, K.; Böttcher, J.; et al. Differences in Labile Soil Organic Matter Explain Potential Denitrification and Denitrifying Communities in a Long-Term Fertilization Experiment. Appl. Soil Ecol. 2020, 153, 103630. [Google Scholar] [CrossRef]
- Saunders, O.E.; Fortuna, A.-M.; Harrison, J.H.; Cogger, C.G.; Whitefield, E.; Green, T. Gaseous Nitrogen and Bacterial Responses to Raw and Digested Dairy Manure Applications in Incubated Soil. Environ. Sci. Technol. 2012, 46, 11684–11692. [Google Scholar] [CrossRef]
- Luo, G.; Friman, V.-P.; Chen, H.; Liu, M.; Wang, M.; Guo, S.; Ling, N.; Shen, Q. Long-Term Fertilization Regimes Drive the Abundance and Composition of N-Cycling-Related Prokaryotic Groups via Soil Particle-Size Differentiation. Soil Biol. Biochem. 2018, 116, 213–223. [Google Scholar] [CrossRef]
- Tatti, E.; Goyer, C.; Zebarth, B.J.; Burton, D.L.; Giovannetti, L.; Viti, C. Short-Term Effects of Mineral and Organic Fertilizer on Denitrifiers, Nitrous Oxide Emissions and Denitrification in Long-Term Amended Vineyard Soils. Soil Sci. Soc. Am. J. 2013, 77, 113–122. [Google Scholar] [CrossRef]
- Dambreville, C.; Hallet, S.; Nguyen, C.; Morvan, T.; Germon, J.-C.; Philippot, L. Structure and Activity of the Denitrifying Community in a Maize-Cropped Field Fertilized with Composted Pig Manure or Ammonium Nitrate: Effect of Fertilization Regime on Denitrifiers. FEMS Microbiol. Ecol. 2006, 56, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, C.; Liu, X.; Lu, Y.; Wang, Y. Saline-Alkali Soil Applied with Vermicompost and Humic Acid Fertilizer Improved Macroaggregate Microstructure to Enhance Salt Leaching and Inhibit Nitrogen Losses. Appl. Soil Ecol. 2020, 156, 103705. [Google Scholar] [CrossRef]
- Dai, Z.; Li, Y.; Zhang, X.; Wu, J.; Luo, Y.; Kuzyakov, Y.; Brookes, P.C.; Xu, J. Easily Mineralizable Carbon in Manure-Based Biochar Added to a Soil Influences N2O Emissions and Microbial-N Cycling Genes. Land Degrad. Dev. 2019, 30, 406–416. [Google Scholar] [CrossRef]
- Wei, J.; Reichel, R.; Islam, M.S.; Wissel, H.; Amelung, W.; Brüggemann, N. Chemical Composition of High Organic Carbon Soil Amendments Affects Fertilizer-Derived N2O Emission and Nitrogen Immobilization in an Oxic Sandy Loam. Front. Environ. Sci. 2020, 8, 15. [Google Scholar] [CrossRef]
- Wang, H.; Xu, J.; Liu, X.; Zhang, D.; Li, L.; Li, W.; Sheng, L. Effects of Long-Term Application of Organic Fertilizer on Improving Organic Matter Content and Retarding Acidity in Red Soil from China. Soil Tillage Res. 2019, 195, 104382. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; He, P.; Zhou, W. Different Roles of Rhizosphere Effect and Long-Term Fertilization in the Activity and Community Structure of Ammonia Oxidizers in a Calcareous Fluvo-Aquic Soil. Soil Biol. Biochem. 2013, 57, 30–42. [Google Scholar] [CrossRef]
- Zhou, X.; Fornara, D.; Wasson, E.A.; Wang, D.; Ren, G.; Christie, P.; Jia, Z. Effects of 44 Years of Chronic Nitrogen Fertilization on the Soil Nitrifying Community of Permanent Grassland. Soil Biol. Biochem. 2015, 91, 76–83. [Google Scholar] [CrossRef]
- Bárta, J.; Melichová, T.; Vaněk, D.; Picek, T.; Šantrůčková, H. Effect of PH and Dissolved Organic Matter on the Abundance of NirK and NirS Denitrifiers in Spruce Forest Soil. Biogeochemistry 2010, 101, 123–132. [Google Scholar] [CrossRef]
- Wolsing, M.; Prieme, A. Observation of High Seasonal Variation in Community Structure of Denitrifying Bacteria in Arable Soil Receiving Artificial Fertilizer and Cattle Manure by Determining T-RFLP of Nir Gene Fragments. FEMS Microbiol. Ecol. 2004, 48, 261–271. [Google Scholar] [CrossRef]
- Yin, C.; Fan, F.; Song, A.; Cui, P.; Li, T.; Liang, Y. Denitrification Potential under Different Fertilization Regimes Is Closely Coupled with Changes in the Denitrifying Community in a Black Soil. Appl. Microbiol. Biotechnol. 2015, 99, 5719–5729. [Google Scholar] [CrossRef]
- Insam, H.; Gómez-Brandón, M.; Ascher, J. Manure-Based Biogas Fermentation Residues—Friend or Foe of Soil Fertility? Soil Biol. Biochem. 2015, 84, 1–14. [Google Scholar] [CrossRef]
- Tambone, F.; Terruzzi, L.; Scaglia, B.; Adani, F. Composting of the Solid Fraction of Digestate Derived from Pig Slurry: Biological Processes and Compost Properties. Waste Manag. 2015, 35, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Möller, K.; Müller, T. Effects of Anaerobic Digestion on Digestate Nutrient Availability and Crop Growth: A Review: Digestate Nutrient Availability. Eng. Life Sci. 2012, 12, 242–257. [Google Scholar] [CrossRef]
- Cayuela, M.L.; Oenema, O.; Kuikman, P.J.; Bakker, R.R.; Van GROENIGEN, J.W. Bioenergy By-Products as Soil Amendments? Implications for Carbon Sequestration and Greenhouse Gas Emissions. GCB Bioenergy 2010, 2, 201–213. [Google Scholar] [CrossRef]
- Grave, R.A.; da Nicoloso, R.S.; Cassol, P.C.; da Silva, M.L.B.; Mezzari, M.P.; Aita, C.; Wuaden, C.R. Determining the Effects of Tillage and Nitrogen Sources on Soil N2O Emission. Soil Tillage Res. 2018, 175, 1–12. [Google Scholar] [CrossRef]
- Lazcano, C.; Gómez-Brandón, M.; Domínguez, J. Comparison of the Effectiveness of Composting and Vermicomposting for the Biological Stabilization of Cattle Manure. Chemosphere 2008, 72, 1013–1019. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Lazcano, C.; Domínguez, J. The Evaluation of Stability and Maturity during the Composting of Cattle Manure. Chemosphere 2008, 70, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Huang, S.; Sha, C.; Wu, J.; Cui, C.; Su, J.; Ruan, J.; Tan, J.; Tang, H.; Xue, J. Changes of Bacterial Community in Arable Soil after Short-Term Application of Fresh Manures and Organic Fertilizer. Environ. Technol. 2020, 1–11. [Google Scholar] [CrossRef]
- Li, P.; Lang, M.; Li, C.; Hao, X. Nitrous Oxide and Carbon Dioxide Emissions from Soils Amended with Compost and Manure from Cattle Fed Diets Containing Wheat Dried Distillers’ Grains with Solubles. Can. J. Soil Sci. 2016, 97, 522–531. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Kolbe, A.R.; Gómez-Brandón, M.; Pérez-Losada, M. Changes in the Composition and Function of Bacterial Communities during Vermicomposting May Explain Beneficial Properties of Vermicompost. Sci. Rep. 2019, 9, 9657. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Brandón, M.; Juárez, M.F.-D.; Zangerle, M.; Insam, H. Effects of Digestate on Soil Chemical and Microbiological Properties: A Comparative Study with Compost and Vermicompost. J. Hazard. Mater. 2016, 302, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Gul, S.; Whalen, J.K. Biochemical Cycling of Nitrogen and Phosphorus in Biochar-Amended Soils. Soil Biol. Biochem. 2016, 103, 1–15. [Google Scholar] [CrossRef]
- Zhu, K.; Christel, W.; Bruun, S.; Jensen, L.S. The Different Effects of Applying Fresh, Composted or Charred Manure on Soil N2O Emissions. Soil Biol. Biochem. 2014, 74, 61–69. [Google Scholar] [CrossRef]
- Cayuela, M.L.; van Zwieten, L.; Singh, B.P.; Jeffery, S.; Roig, A.; Sánchez-Monedero, M.A. Biochar’s Role in Mitigating Soil Nitrous Oxide Emissions: A Review and Meta-Analysis. Agric. Ecosyst. Environ. 2014, 191, 5–16. [Google Scholar] [CrossRef]
- Pujol Pereira, E.I.; Léchot, J.; Feola Conz, R.; da Silva Cardoso, A.; Six, J. Biochar Enhances Nitrous Oxide Reduction in Acidic but Not in Near-Neutral pH Soil. Soil Syst. 2019, 3, 69. [Google Scholar] [CrossRef]
- Ruiz Diaz, D.A.; Sawyer, J.E.; Mallarino, A.P. Poultry Manure Supply of Potentially Available Nitrogen with Soil Incubation. Agron. J. 2008, 100, 1310–1317. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, H.; Yuan, W.; Williams, D.; Walker, J.T.; Shi, W. Is Biochar-Manure Co-Compost a Better Solution for Soil Health Improvement and N2O Emissions Mitigation? Soil Biol. Biochem. 2017, 113, 14–25. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Wu, D.; Kong, X.; Bol, R.; Petersen, S.O.; Jensen, L.S.; Liu, S.; Brüggemann, N.; Glud, R.N.; Larsen, M.; et al. Effects of Cattle Slurry and Nitrification Inhibitor Application on Spatial Soil O2 Dynamics and N2O Production Pathways. Soil Biol. Biochem. 2017, 114, 200–209. [Google Scholar] [CrossRef]
- Duan, P.; Zhang, Q.; Zhang, X.; Xiong, Z. Mechanisms of Mitigating Nitrous Oxide Emissions from Vegetable Soil Varied with Manure, Biochar and Nitrification Inhibitors. Agric. For. Meteorol. 2019, 278, 107672. [Google Scholar] [CrossRef]
- Lazicki, P.; Geisseler, D.; Lloyd, M. Nitrogen Mineralization from Organic Amendments Is Variable but Predictable. J. Environ. Qual. 2020, 49, 483–495. [Google Scholar] [CrossRef] [PubMed]
- VanderZaag, A.C.; Jayasundara, S.; Wagner-Riddle, C. Strategies to Mitigate Nitrous Oxide Emissions from Land Applied Manure. Anim. Feed Sci. Technol. 2011, 166–167, 464–479. [Google Scholar] [CrossRef]
- Bowles, T.M.; Hollander, A.D.; Steenwerth, K.; Jackson, L.E. Tightly-Coupled Plant-Soil Nitrogen Cycling: Comparison of Organic Farms across an Agricultural Landscape. PLoS ONE 2015, 10, e0131888. [Google Scholar] [CrossRef]
- Aguilera, E.; Lassaletta, L.; Sanz-Cobena, A.; Garnier, J.; Vallejo, A. The Potential of Organic Fertilizers and Water Management to Reduce N2O Emissions in Mediterranean Climate Cropping Systems. A Review. Agric. Ecosyst. Environ. 2013, 164, 32–52. [Google Scholar] [CrossRef]
- Bhowmik, A.; Fortuna, A.-M.; Cihacek, L.J.; Bary, A.I.; Cogger, C.G. Use of Biological Indicators of Soil Health to Estimate Reactive Nitrogen Dynamics in Long-Term Organic Vegetable and Pasture Systems. Soil Biol. Biochem. 2016, 103, 308–319. [Google Scholar] [CrossRef]
- Lazcano, C.; Barrios-Masias, F.H.; Jackson, L.E. Arbuscular Mycorrhizal Effects on Plant Water Relations and Soil Greenhouse Gas Emissions under Changing Moisture Regimes. Soil Biol. Biochem. 2014, 74, 184–192. [Google Scholar] [CrossRef]
- Bender, S.F.; Plantenga, F.; Neftel, A.; Jocher, M.; Oberholzer, H.-R.; Köhl, L.; Giles, M.; Daniell, T.J.; van der Heijden, M.G. Symbiotic Relationships between Soil Fungi and Plants Reduce N2O Emissions from Soil. ISME J. 2014, 8, 1336–1345. [Google Scholar] [CrossRef]
- Bowles, T.M.; Jackson, L.E.; Loeher, M.; Cavagnaro, T.R. Ecological Intensification and Arbuscular Mycorrhizas: A Meta-Analysis of Tillage and Cover Crop Effects. J. Appl. Ecol. 2017, 54, 1785–1793. [Google Scholar] [CrossRef]
- Krauss, M.; Krause, H.-M.; Spangler, S.; Kandeler, E.; Behrens, S.; Kappler, A.; Mäder, P.; Gattinger, A. Tillage System Affects Fertilizer-Induced Nitrous Oxide Emissions. Biol. Fertil. Soils 2017, 53, 49–59. [Google Scholar] [CrossRef]
- Marquina, S.; Pérez, T.; Donoso, L.; Giuliante, A.; Rasse, R.; Herrera, F. NO, N2O and CO2 Soil Emissions from Venezuelan Corn Fields under Tillage and No-Tillage Agriculture. Nutr. Cycl. Agroecosyst. 2015, 101, 123–137. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen Mineralization: Challenges of a Changing Paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Baldrian, P. The Known and the Unknown in Soil Microbial Ecology. FEMS Microbiol. Ecol. 2019, 95, fiz005. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.K.; Hofmockel, K.S. Soil Microbiomes and Climate Change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.M.; Abell, G.; Philippot, L.; Prosser, J.; et al. Microbes as Engines of Ecosystem Function: When Does Community Structure Enhance Predictions of Ecosystem Processes? Front. Microbiol. 2016, 7, 214. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Transformation | Enzyme | Gen | Microorganism |
|---|---|---|---|
| NH4+ -> NH2OH | Ammonia monooxygenase | amoA | Bacteria and Archaea |
| NH2OH -> NO | Hydroxylamine oxidoreductase | hao | Bacteria |
| NH2OH -> N2O | Cytochrome P460 | cyt P460 | Bacteria |
| NO -> NO2− | Dinitrite reductase | nirK | Bacteria |
| NO2− -> NO3− | Nitrite oxidoreductase | nxr | Bacteria |
| NO2− -> NH4+ | Periplasmic nitrite reductase | nrf | Bacteria |
| NO3− -> NO2− | Membrane bound dissimilatory nitrate reductase | narG | Fungi, Bacteria |
| Periplasmatic dissimilatory nitrate reductase | nap | Fungi, Bacteria | |
| NO2− -> NO | Cd1 nitrite reductase | nirS | Bacteria |
| Copper nitrite reductase | nirK | Bacteria, fungi | |
| NO -> N2O | Nitric oxide reductase | norB | Bacteria |
| Quinol nitric oxide reductase | qnorB | Bacteria | |
| Cytochrome P450nor nitric oxide reductase | P450nor | Fungi | |
| Nitrosocyanin | ncyA | Bacteria | |
| N2O -> N2 | Nitrous oxide reductase | nosZ Clade I, nosZ Clade II | Bacteria, Fungi |
| Nitrosocyanin | ncyA | Bacteria |
| Nitrification | Denitrification | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| amo | hao | nxr | narG | nap | nirS | nirK | norB | nosZ | ncyA | References | |
| Manure | ↑ | ? | ? | ↑ | ? | ↑ | ↑ | ? | ↑ | ? | [37,56,68,71,72,73] |
| Digestate | ↑ | ? | ? | ↑ | ? | ? | ? | ? | ↑ | ? | [74] |
| Compost | ↑ | ? | ? | ↑ | ? | ↑ | ↑ | ↓↑ | ↑ | ? | [70,75,76,77] |
| Vermicompost | ↑ | ? | ? | ? | ? | ↓ | ↓ | ? | ↑ | ? | [78] |
| Biochar | ? | ? | ? | ? | ? | ↑ | ↑ | ? | ↑ | ? | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazcano, C.; Zhu-Barker, X.; Decock, C. Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review. Microorganisms 2021, 9, 983. https://doi.org/10.3390/microorganisms9050983
Lazcano C, Zhu-Barker X, Decock C. Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review. Microorganisms. 2021; 9(5):983. https://doi.org/10.3390/microorganisms9050983
Chicago/Turabian StyleLazcano, Cristina, Xia Zhu-Barker, and Charlotte Decock. 2021. "Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review" Microorganisms 9, no. 5: 983. https://doi.org/10.3390/microorganisms9050983
APA StyleLazcano, C., Zhu-Barker, X., & Decock, C. (2021). Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review. Microorganisms, 9(5), 983. https://doi.org/10.3390/microorganisms9050983