
A Mediation Analysis to Identify Links between Gut Bacteria and Memory in Context of Human Milk Oligosaccharides

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Dietary Treatments
2.3. Behavior
2.4. Magnetic Resonance Imaging (MRI)
2.5. Hippocampal Gene Expression
2.6. DNA Extraction
2.7. PCR Amplification and Sequencing of 16S rRNA Genes
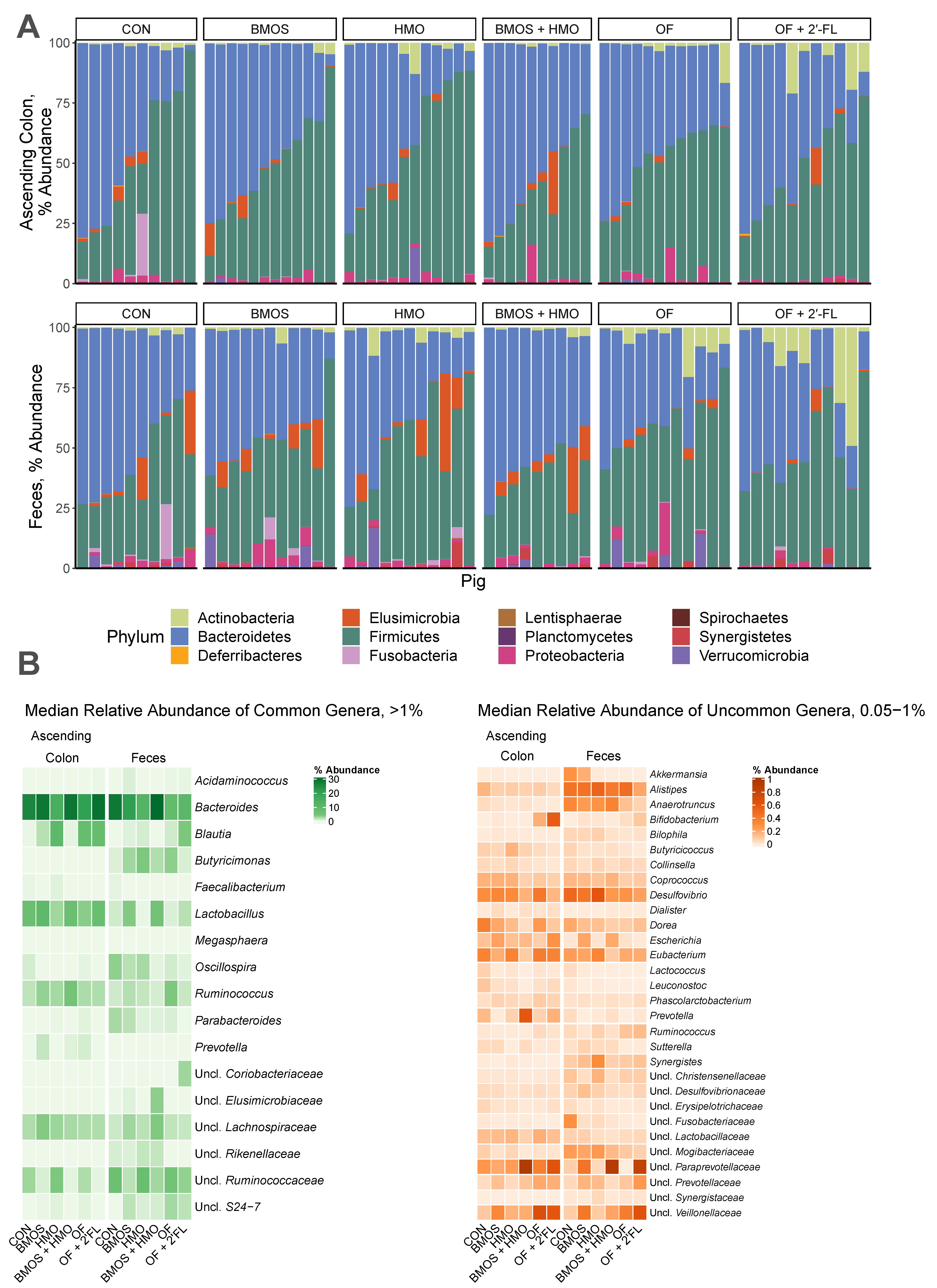
2.8. Sequence Processing
2.9. Statistical Analysis
2.9.1. Data Cleaning and Variable Selection
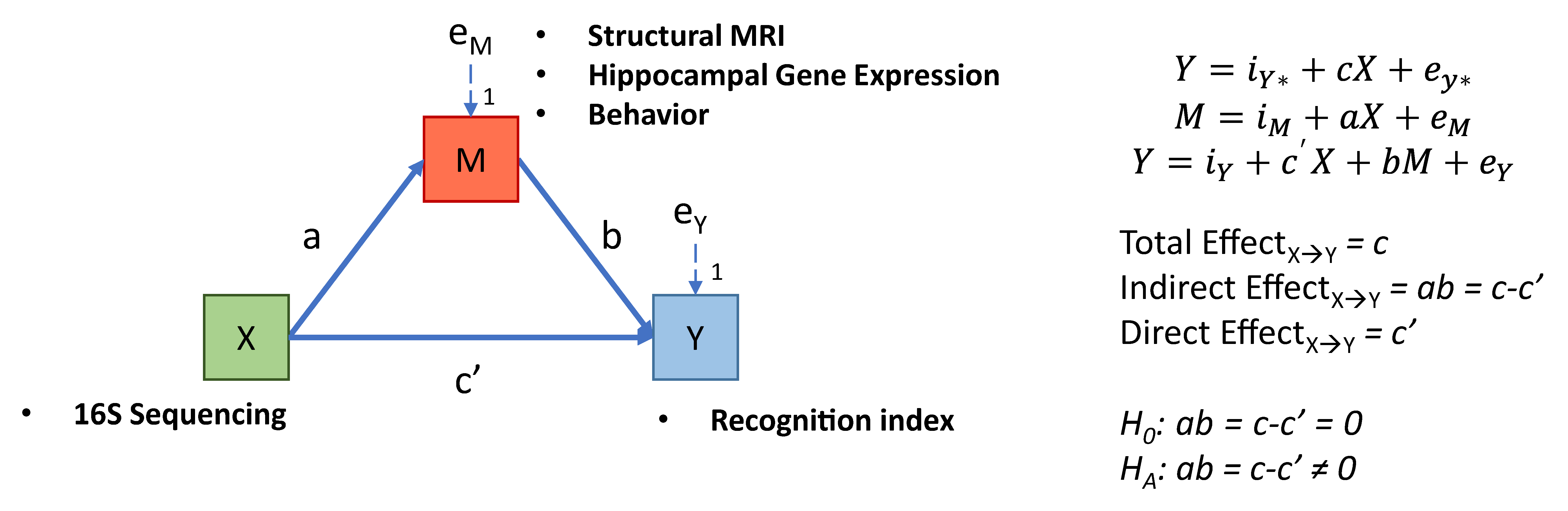
2.9.2. Mediation Analysis
3. Results
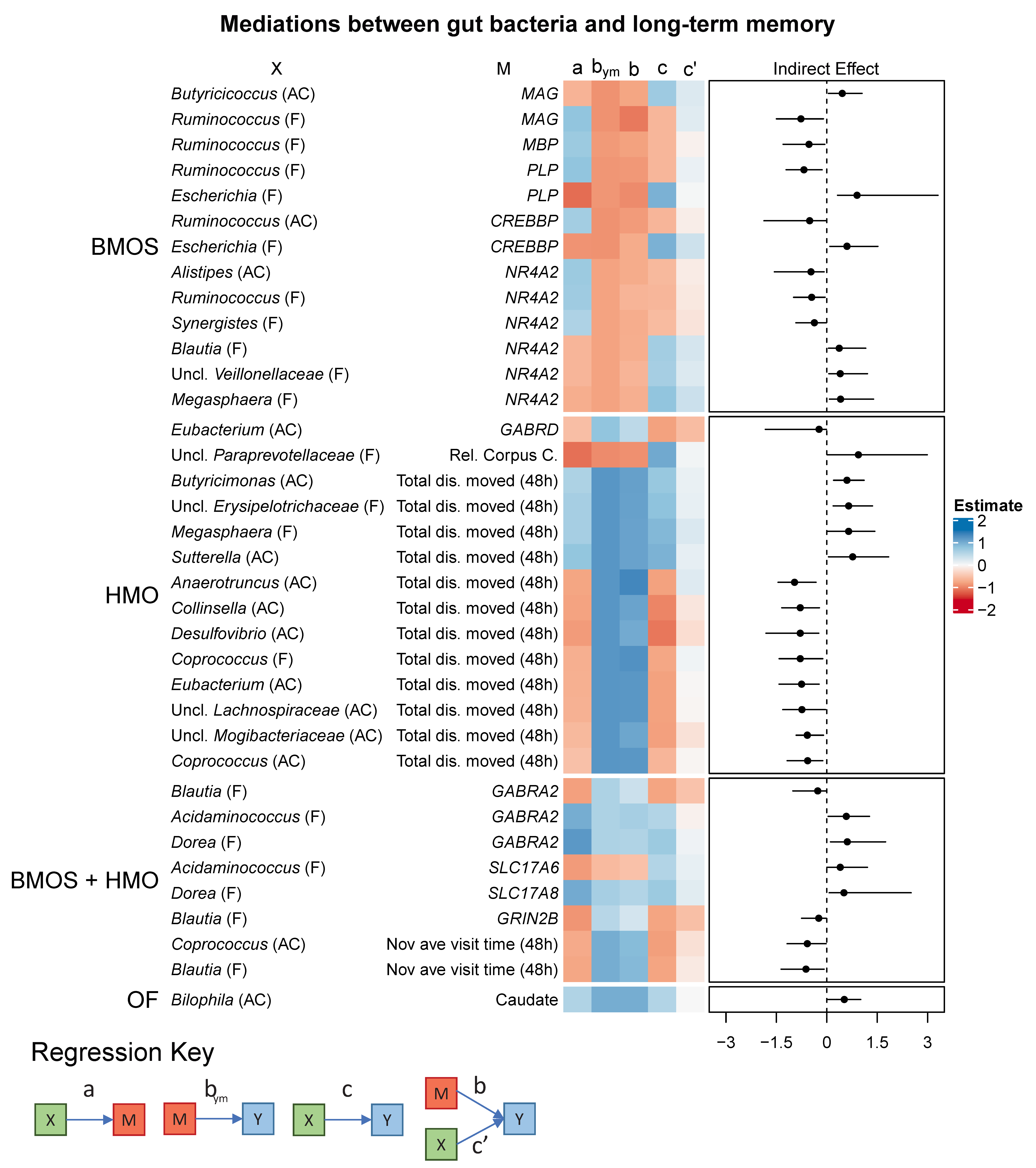
3.1. Short-Term Memory
3.2. Long-Term Memory
3.3. Suppressor Variables
4. Discussion
4.1. Evidence of Common Paths to Cognition
4.1.1. GABA and Glutamatergic Mediators
4.1.2. Myelination and Transcription Factors
4.1.3. Exploratory Behavior
4.1.4. Brain Volume
4.2. Potential Mechanisms of Action of Prebiotics
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Georgieff, M.K. Long-term Brain and Behavioral Consequences of Early Iron Deficiency. Nutr. Rev. 2011, 69, S43–S48. [Google Scholar] [CrossRef]
- Zeisel, S.H. Choline: Needed for Normal Development of Memory. J. Am. Coll. Nutr. 2000, 19, 528S–531S. [Google Scholar] [CrossRef]
- Mudd, A.T.; Dilger, R.N. Early-life nutrition and neurodevelopment: Use of the piglet as a translational model. Adv. Nutr. 2017, 8, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Prado, E.L.; Dewey, K.G. Nutrition and brain development in early life. Nutr. Rev. 2014, 72, 267–284. [Google Scholar] [CrossRef]
- Schneider, N.; Garcia-Rodenas, C.L. Early nutritional interventions for brain and cognitive development in preterm infants: A review of the literature. Nutrients 2017, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of milk and colostrum in non-human mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef]
- Jeurink, P.V.; van Esch, B.C.; Rijnierse, A.; Garssen, J.; Knippels, L.M. Mechanisms underlying immune effects of dietary oligosaccharides. Am. J. Clin. Nutr. 2013, 98, 572S–577S. [Google Scholar] [CrossRef]
- Donovan, S.M.; Comstock, S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016, 69 (Suppl. 2), 42–51. [Google Scholar] [CrossRef] [PubMed]
- Asakuma, S.; Hatakeyama, E.; Urashima, T.; Yoshida, E.; Katayama, T.; Yamamoto, K.; Kumagai, H.; Ashida, H.; Hirose, J.; Kitaoka, M. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J. Biol. Chem. 2011, 286, 34583–34592. [Google Scholar] [CrossRef] [PubMed]
- Mudd, A.T.; Fleming, S.A.; Labhart, B.; Chichlowski, M.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Sialyllactose Influences Sialic Acid Concentrations in the Prefrontal Cortex and Magnetic Resonance Imaging Measures in Corpus Callosum of Young Pigs. Nutrients 2017, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, S.K.; Yatsunenko, T.; Li, D.; Dasgupta, S.; Yu, R.K.; Berg, B.M.; Chichlowski, M.; Odle, J. Dietary Isomers of Sialyllactose Increase Ganglioside Sialic Acid Concentrations in the Corpus Callosum and Cerebellum and Modulate the Colonic Microbiota of Formula-Fed Piglets. J. Nutr. 2016, 146, 200–208. [Google Scholar] [CrossRef]
- Tarr, A.J.; Galley, J.D.; Fisher, S.E.; Chichlowski, M.; Berg, B.M.; Bailey, M.T. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: Evidence for effects on the gut–brain axis. Brain. Behav. Immun. 2015, 50, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, E.; Ramirez, M.; Vazquez, E.; Barranco, A.; Gruart, A.; Delgado-Garcia, J.M.; Buck, R.; Rueda, R.; Martin, M.J. Oral supplementation of 2′-fucosyllactose during lactation improves memory and learning in rats. J. Nutr. Biochem. 2016, 31, 20–27. [Google Scholar] [CrossRef]
- Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.M.; Martínez-Lara, E.; Blanco, S.; Martín, M.J.; Castanys, E.; Buck, R.; et al. Effects of a human milk oligosaccharide, 2′-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J. Nutr. Biochem. 2015, 26, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, E.; Barranco, A.; Ramirez, M.; Gruart, A.; Delgado-Garcia, J.M.; Jimenez, M.L.; Buck, R.; Rueda, R. Dietary 2′-Fucosyllactose Enhances Operant Conditioning and Long-Term Potentiation via Gut-Brain Communication through the Vagus Nerve in Rodents. PLoS ONE 2016, 11, e0166070. [Google Scholar] [CrossRef]
- Jia, S.; Lu, Z.; Gao, Z.; An, J.; Wu, X.; Li, X.; Dai, X.; Zheng, Q.; Sun, Y. Chitosan oligosaccharides alleviate cognitive deficits in an amyloid-β1–42-induced rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 83, 416–425. [Google Scholar] [CrossRef]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human milk oligosaccharide 2′-fucosyllactose links feedings at 1 month to cognitive development at 24 months in infants of normal and overweight mothers. PLoS ONE 2020, 15, e0228323. [Google Scholar] [CrossRef]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.; Shanahan, F.; Dinan, T.; Cryan, J. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Liu, M.; Chen, J.; Pan, J.; Han, Y.; Liu, Y.; Cheng, K.; Zhou, C.; Wang, H.; et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci. Adv. 2019, 5, 1–11. [Google Scholar] [CrossRef]
- Gacias, M.; Gaspari, S.; Santos, P.-M.G.; Tamburini, S.; Andrade, M.; Zhang, F.; Shen, N.; Tolstikov, V.; Kiebish, M.A.; Dupree, J.L.; et al. Microbiota-driven transcriptional changes in prefrontal cortex override genetic differences in social behavior. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Hanstock, T.L.; Clayton, E.H.; Li, K.M.; Mallet, P.E. Anxiety and aggression associated with the fermentation of carbohydrates in the hindgut of rats. Physiol. Behav. 2004, 82, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Hanstock, T.L.; Mallet, P.E.; Clayton, E.H. Increased plasma d-lactic acid associated with impaired memory in rats. Physiol. Behav. 2010, 101, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.L.; Xia, K.; Azcarate-Peril, M.A.; Goldman, B.D.; Ahn, M.; Styner, M.A.; Thompson, A.L.; Geng, X.; Gilmore, J.H.; Knickmeyer, R.C. Infant Gut Microbiome Associated With Cognitive Development. Biol. Psychiatry 2018, 83, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Golubeva, A.V.; Joyce, S.A.; Moloney, G.; Burokas, A.; Sherwin, E.; Arboleya, S.; Flynn, I.; Khochanskiy, D.; Moya-Pérez, A.; Peterson, V.; et al. Microbiota-related Changes in Bile Acid & Tryptophan Metabolism are Associated with Gastrointestinal Dysfunction in a Mouse Model of Autism. EBioMedicine 2017, 24, 166–178. [Google Scholar] [CrossRef]
- Luna, R.A.; Oezguen, N.; Balderas, M.; Venkatachalam, A.; Runge, J.K.; Versalovic, J.; Veenstra-VanderWeele, J.; Anderson, G.M.; Savidge, T.; Williams, K.C. Distinct Microbiome-Neuroimmune Signatures Correlate With Functional Abdominal Pain in Children With Autism Spectrum Disorder. CMGH 2017, 3, 218–230. [Google Scholar] [CrossRef]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef]
- Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression. Nutrients 2020, 12, 2131. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Human and Bovine Milk Oligosaccharides Elicit Improved Recognition Memory Concurrent with Alterations in Regional Brain Volumes and Hippocampal mRNA Expression. Front. Neurosci. 2020, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Monaco, M.H.; Hauser, J.; Yan, J.; Dilger, R.N.; Donovan, S.M. Bovine Milk Oligosaccharides and Human Milk Oligosaccharides Modulate the Gut Microbiota Composition and Volatile Fatty Acid Concentrations in a Neonatal Model. Microorganisms 2021, 2, 331–351. [Google Scholar]
- Niilo, L. Clostridium perfringens Type C Enterotoxemia. Can. Vet. J. 1988, 29, 658–664. [Google Scholar] [PubMed]
- Meli, F.; Puccio, G.; Cajozzo, C.; Ricottone, G.L.; Pecquet, S.; Sprenger, N.; Steenhout, P. Growth and safety evaluation of infant formulae containing oligosaccharides derived from bovine milk: A randomized, double-blind, noninferiority trial. BMC Pediatr. 2014, 14, 306. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.; Bolton, K.D.; Velaphi, S.; De Groot, N.; Emady-Azar, S.; Pecquet, S.; Steenhout, P. Early Benefits of a Starter Formula Enriched in Prebiotics and Probiotics on the Gut Microbiota of Healthy Infants Born to HIV+ Mothers: A Randomized Double-Blind Controlled Trial. Clin. Med. Insights Pediatr. 2016, 10, 119–130. [Google Scholar] [CrossRef]
- Radke, M.; Picaud, J.C.; Loui, A.; Cambonie, G.; Faas, D.; Lafeber, H.N.; De Groot, N.; Pecquet, S.S.; Steenhout, P.G.; Hascoet, J.M. Starter formula enriched in prebiotics and probiotics ensures normal growth of infants and promotes gut health: A randomized clinical trial. Pediatr. Res. 2017, 81, 622–631. [Google Scholar] [CrossRef]
- Puccio, G.; Alliet, P.; Cajozzo, C.; Janssens, E.; Corsello, G.; Sprenger, N.; Wernimont, S.; Egli, D.; Gosoniu, L.; Steenhout, P. Effects of infant formula with human milk oligosaccharides on growth and morbidity: A randomized multicenter trial. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Marriage, B.J.; Buck, R.H.; Goehring, K.C.; Oliver, J.S.; Williams, J.A. Infants Fed a Lower Calorie Formula with 2′FL Show Growth and 2′FL Uptake Like Breast-Fed Infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Lien, E.L.; Capeding, M.R.Z.; Fitzgerald, M.; Ramanujam, K.; Yuhas, R.; Northington, R.; Lebumfacil, J.; Wang, L.; DeRusso, P.A. Effects of Term Infant Formulas Containing High sn-2 Palmitate With and Without Oligofructose on Stool Composition, Stool Characteristics, and Bifidogenicity. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 440–448. [Google Scholar] [CrossRef]
- Wernimont, S.; Northington, R.; Kullen, M.J.; Yao, M.; Bettler, J. Effect of an α-lactalbumin-enriched infant formula supplemented with oligofructose on fecal microbiota, stool characteristics, and hydration status: A randomized, double-blind, controlled trial. Clin. Pediatr. 2015, 54, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Moustgaard, A.; Lind, N.M.; Hemmingsen, R.; Hansen, A.K. Spontaneous Object Recognition in the Göttingen Minipig. Neural Plast. 2002, 9, 255–259. [Google Scholar] [CrossRef]
- Kornum, B.R.; Thygesen, K.S.; Nielsen, T.R.; Knudsen, G.M.; Lind, N.M. The effect of the inter-phase delay interval in the spontaneous object recognition test for pigs. Behav. Brain Res. 2007, 181, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Gifford, A.K.; Cloutier, S.; Newberry, R.C. Objects as enrichment: Effects of object exposure time and delay interval on object recognition memory of the domestic pig. Appl. Anim. Behav. Sci. 2007, 107, 206–217. [Google Scholar] [CrossRef]
- Kouwenberg, A.-L.; Walsh, C.J.; Morgan, B.E.; Martin, G.M. Episodic-like memory in crossbred Yucatan minipigs (Sus scrofa). Appl. Anim. Behav. Sci. 2009, 117, 165–172. [Google Scholar] [CrossRef]
- Fleming, S.A.; Dilger, R.N. Young pigs exhibit differential exploratory behavior during novelty preference tasks in response to age, sex, and delay. Behav. Brain Res. 2017, 321, 50–60. [Google Scholar] [CrossRef]
- Fleming, S.A.; Monaikul, S.; Patsavas, A.J.; Waworuntu, R.V.; Berg, B.M.; Dilger, R.N. Dietary polydextrose and galactooligosaccharide increase exploratory behavior, improve recognition memory, and alter neurochemistry in the young pig. Nutr. Neurosci. 2017, 22, 499–512. [Google Scholar] [CrossRef]
- Fleming, S.; Chichlowski, M.; Berg, B.; Donovan, S.; Dilger, R. Dietary Sialyllactose Does Not Influence Measures of Recognition Memory or Diurnal Activity in the Young Pig. Nutrients 2018, 10, 395. [Google Scholar] [CrossRef]
- Jacob, R.M.; Mudd, A.T.; Alexander, L.S.; Lai, C.-S.; Dilger, R.N. Comparison of Brain Development in Sow-Reared and Artificially Reared Piglets. Front. Pediatr. 2016, 4, 95. [Google Scholar] [CrossRef]
- Radlowski, E.C.; Conrad, M.S.; Lezmi, S.; Dilger, R.N.; Sutton, B.; Larsen, R.; Johnson, R.W. A neonatal piglet model for investigating brain and cognitive development in small for gestational age human infants. PLoS ONE 2014, 9, e91951. [Google Scholar] [CrossRef]
- Mudd, A.T.; Getty, C.M.; Sutton, B.P.; Dilger, R.N. Perinatal choline deficiency delays brain development and alters metabolite concentrations in the young pig. Nutr. Neurosci. 2016, 19, 425–433. [Google Scholar] [CrossRef]
- Reznikov, E.A.; Comstock, S.S.; Hoeflinger, J.L.; Wang, M.; Miller, M.J.; Donovan, S.M. Dietary Bovine Lactoferrin Reduces Staphylococcus aureus in the Tissues and Modulates the Immune Response in Piglets Systemically Infected with S. aureus. Curr. Dev. Nutr. 2018, 2, nzy001. [Google Scholar] [CrossRef]
- Monaco, M.H.; Wang, M.; Pan, X.; Li, Q.; Richards, J.D.; Chichlowski, M.; Berg, B.M.; Dilger, R.N.; Donovan, S.M. Evaluation of Sialyllactose Supplementation of a Prebiotic-Containing Formula on Growth, Intestinal Development, and Bacterial Colonization in the Neonatal Piglet. Curr. Dev. Nutr. 2018, 2, nzy067. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R. A Language and Environment for Statistical Computing; Vienna, Austria. 2019. Available online: https://www.R-project.org/ (accessed on 1 January 2021).
- Tingley, D.; Yamamoto, T.; Hirose, K.; Keele, L.; Imai, K. mediation: R package for Causal Mediation Analysis. J. Stat. Softw. 2014, 59, 1–38. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Hauke, J.; Kossowski, T.M. Comparison of values of pearson’s and spearman’s correlation coefficients on the same sets of data. Quaest. Geogr. 2011, 30, 87–93. [Google Scholar] [CrossRef]
- Hayes, A.F.; Rockwood, N.J. Regression-based statistical mediation and moderation analysis in clinical research: Observations, recommendations, and implementation. Behav. Res. Ther. 2017, 98, 39–57. [Google Scholar] [CrossRef]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis, 2nd ed.; Little, T., Ed.; The Guilford Press: New York, NY, USA, 2018. [Google Scholar]
- Hines, R.M.; Hines, D.J.; Houston, C.M.; Mukherjee, J.; Haydon, P.G.; Tretter, V.; Smart, T.G.; Moss, S.J. Disrupting the clustering of GABAA receptor α2 subunits in the frontal cortex leads to reduced γ-power and cognitive deficits. Proc. Natl. Acad. Sci. USA 2013, 110, 16628–16633. [Google Scholar] [CrossRef]
- Ge, Y.; Dong, Z.; Bagot, R.C.; Howland, J.G.; Phillips, A.G.; Wong, T.P.; Wang, Y.T. Hippocampal long-term depression is required for the consolidation of spatial memory. Proc. Natl. Acad. Sci. USA 2010, 107, 16697–16702. [Google Scholar] [CrossRef]
- Brigman, J.L.; Wright, T.; Talani, G.; Prasad-Mulcare, S.; Jinde, S.; Seabold, G.K.; Mathur, P.; Davis, M.I.; Bock, R.; Gustin, R.M.; et al. Loss of GluN2B-Containing NMDA Receptors in CA1 Hippocampus and Cortex Impairs Long-Term Depression, Reduces Dendritic Spine Density, and Disrupts Learning. J. Neurosci. 2010, 30, 4590–4600. [Google Scholar] [CrossRef]
- Zhao, X.; Rosenke, R.; Kronemann, D.; Brim, B.; Das, S.R.; Dunah, A.W.; Magnusson, K.R. The effects of aging on N-methyl-D-aspartate receptor subunits in the synaptic membrane and relationships to long-term spatial memory. Neuroscience 2009, 162, 933–945. [Google Scholar] [CrossRef]
- Li, H.; Santos, M.S.; Park, C.K.; Dobry, Y.; Voglmaier, S.M. VGLUT2 Trafficking Is Differentially Regulated by Adaptor Proteins AP-1 and AP-3. Front. Cell. Neurosci. 2017, 11, 324. [Google Scholar] [CrossRef]
- Birgner, C.; Nordenankar, K.; Lundblad, M.; Mendez, J.A.; Smith, C.; le Grevès, M.; Galter, D.; Olson, L.; Fredriksson, A.; Trudeau, L.-E.; et al. VGLUT2 in dopamine neurons is required for psychostimulant-induced behavioral activation. Proc. Natl. Acad. Sci. USA 2010, 107, 389–394. [Google Scholar] [CrossRef]
- Ménard, C.; Quirion, R.; Vigneault, E.; Bouchard, S.; Ferland, G.; El Mestikawy, S.; Gaudreau, P. Glutamate presynaptic vesicular transporter and postsynaptic receptor levels correlate with spatial memory status in aging rat models. Neurobiol. Aging 2015, 36, 1471–1482. [Google Scholar] [CrossRef]
- Amilhon, B.; Lepicard, E.; Renoir, T.; Mongeau, R.; Popa, D.; Poirel, O.; Miot, S.; Gras, C.; Gardier, A.M.; Gallego, J.; et al. VGLUT3 (Vesicular Glutamate Transporter Type 3) Contribution to the Regulation of Serotonergic Transmission and Anxiety. J. Neurosci. 2010, 30, 2198–2210. [Google Scholar] [CrossRef]
- Fazekas, C.L.; Balázsfi, D.; Horváth, H.R.; Balogh, Z.; Aliczki, M.; Puhova, A.; Balagova, L.; Chmelova, M.; Jezova, D.; Haller, J.; et al. Consequences of VGluT3 deficiency on learning and memory in mice. Physiol. Behav. 2019, 112688. [Google Scholar] [CrossRef]
- Tabouy, L.; Getselter, D.; Ziv, O.; Karpuj, M.; Tabouy, T.; Lukic, I.; Maayouf, R.; Werbner, N.; Ben-Amram, H.; Nuriel-Ohayon, M.; et al. Dysbiosis of microbiome and probiotic treatment in a genetic model of autism spectrum disorders. Brain. Behav. Immun. 2018, 73, 310–319. [Google Scholar] [CrossRef]
- Hoban, A.E.; Stilling, R.M.; Ryan, F.J.; Shanahan, F.; Dinan, T.G.; Claesson, M.J.; Clarke, G.; Cryan, J.F. Regulation of prefrontal cortex myelination by the microbiota. Transl. Psychiatry 2016, 6, e774. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Lu, L.; Yu, Y.; Cluette-Brown, J.; Martin, C.R.; Claud, E.C. Effects of Intestinal Microbiota on Brain Development in Humanized Gnotobiotic Mice. Sci. Rep. 2018, 8, 5443. [Google Scholar] [CrossRef]
- Heijtz, R.D.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, L.; Aceto, S.; Agnisola, C.; De Paolo, S.; Dipineto, L.; Stilling, R.M.; Dinan, T.G.; Cryan, J.F.; Menna, L.F.; Fioretti, A. Probiotic modulation of the microbiota-gut-brain axis and behaviour in zebrafish. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Lu, J.; Synowiec, S.; Lu, L.; Yu, Y.; Bretherick, T.; Takada, S.; Yarnykh, V.; Caplan, J.; Caplan, M.; Claud, E.C.; et al. Microbiota influence the development of the brain and behaviors in C57BL/6J mice. PLoS ONE 2018, 13, e0201829. [Google Scholar] [CrossRef]
- Sampson, T.R.; Mazmanian, S.K. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe 2015, 17, 565–576. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bovine Milk Oligosaccharides 2 | 2′Fucosyllactose 3 | Lacto-N-Neotetraose 3 | Oligofructose 4 | Additional Lactose | Total OS | Total Lactose | Total Carbohydrate | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Per kg | Per L | Per kg | Per L | Per kg | Per L | Per kg | Per L | Per kg | Per L | Per kg | Per L | Per kg | Per L | Per kg | Per L |
| Formulated | ||||||||||||||||
| CON | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 69.30 | 13.86 | 0 | 0 | 412.79 | 82.56 | 412.79 | 82.56 |
| BMOS | 61.98 | 12.40 | 0 | 0 | 0 | 0 | 0 | 0 | 61.98 | 1.47 | 61.98 | 12.40 | 350.82 | 70.16 | 412.79 | 82.56 |
| HMO | 0 | 0 | 4.92 | 0.98 | 2.41 | 0.48 | 0 | 0 | 7.33 | 12.40 | 7.33 | 1.47 | 405.47 | 81.09 | 412.79 | 82.56 |
| BMOS + HMO | 61.98 | 12.40 | 4.92 | 0.98 | 2.41 | 0.48 | 0 | 0 | 0 | 0 | 69.30 | 13.86 | 343.49 | 68.70 | 412.79 | 82.56 |
| OF | 0 | 0 | 0 | 0 | 0 | 0 | 26.08 | 5.22 | 61.98 | 8.64 | 26.08 | 5.22 | 386.71 | 77.34 | 412.79 | 82.56 |
| OF + 2′-FL | 0 | 0 | 4.92 | 0.98 | 0 | 0 | 26.08 | 5.22 | 38.30 | 7.66 | 31.00 | 6.20 | 381.79 | 76.36 | 412.79 | 82.56 |
| Analyzed | ||||||||||||||||
| CON | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | NQ | NQ | 0 | 0 | NQ | NQ | NQ | NQ |
| BMOS | 28.95 | 5.79 | 0 | 0 | 0 | 0 | 0 | 0 | NQ | NQ | 28.95 | 5.79 | NQ | NQ | NQ | NQ |
| HMO | 0 | 0 | 4.05 | 0.81 | 2.10 | 0.42 | 0 | 0 | NQ | NQ | 6.15 | 1.23 | NQ | NQ | NQ | NQ |
| BMOS + HMO | 28.75 | 5.75 | 5.00 | 1.00 | 2.65 | 0.53 | 0 | 0 | NQ | NQ | 36.40 | 7.28 | NQ | NQ | NQ | NQ |
| OF | 0 | 0 | 0 | 0 | 0 | 0 | 18.10 | 3.62 | NQ | NQ | 18.10 | 3.62 | NQ | NQ | NQ | NQ |
| OF + 2′-FL | 0 | 0 | 5.60 | 1.12 | 0 | 0 | 17.05 | 3.41 | NQ | NQ | 22.65 | 4.53 | NQ | NQ | NQ | NQ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleming, S.A.; Hauser, J.; Yan, J.; Donovan, S.M.; Wang, M.; Dilger, R.N. A Mediation Analysis to Identify Links between Gut Bacteria and Memory in Context of Human Milk Oligosaccharides. Microorganisms 2021, 9, 846. https://doi.org/10.3390/microorganisms9040846
Fleming SA, Hauser J, Yan J, Donovan SM, Wang M, Dilger RN. A Mediation Analysis to Identify Links between Gut Bacteria and Memory in Context of Human Milk Oligosaccharides. Microorganisms. 2021; 9(4):846. https://doi.org/10.3390/microorganisms9040846
Chicago/Turabian StyleFleming, Stephen A., Jonas Hauser, Jian Yan, Sharon M. Donovan, Mei Wang, and Ryan N. Dilger. 2021. "A Mediation Analysis to Identify Links between Gut Bacteria and Memory in Context of Human Milk Oligosaccharides" Microorganisms 9, no. 4: 846. https://doi.org/10.3390/microorganisms9040846
APA StyleFleming, S. A., Hauser, J., Yan, J., Donovan, S. M., Wang, M., & Dilger, R. N. (2021). A Mediation Analysis to Identify Links between Gut Bacteria and Memory in Context of Human Milk Oligosaccharides. Microorganisms, 9(4), 846. https://doi.org/10.3390/microorganisms9040846