
Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Condition
2.2. Transposon Mutagenesis
2.3. Cloning of Mab_3083c and Msmeg_2685 and Creation of Mab_3083cD89K,H90A Mutant
2.4. Sliding Motility Assay
2.5. Aggregation Assay
2.6. Lipid Extraction and Thin-Layer Chromatography (TLC)
2.7. Intracellular Survival Assay
2.8. Lysozyme and H2O2 Susceptibility Assay
2.9. Sequence Alignment
2.10. Statistical Analysis
3. Results
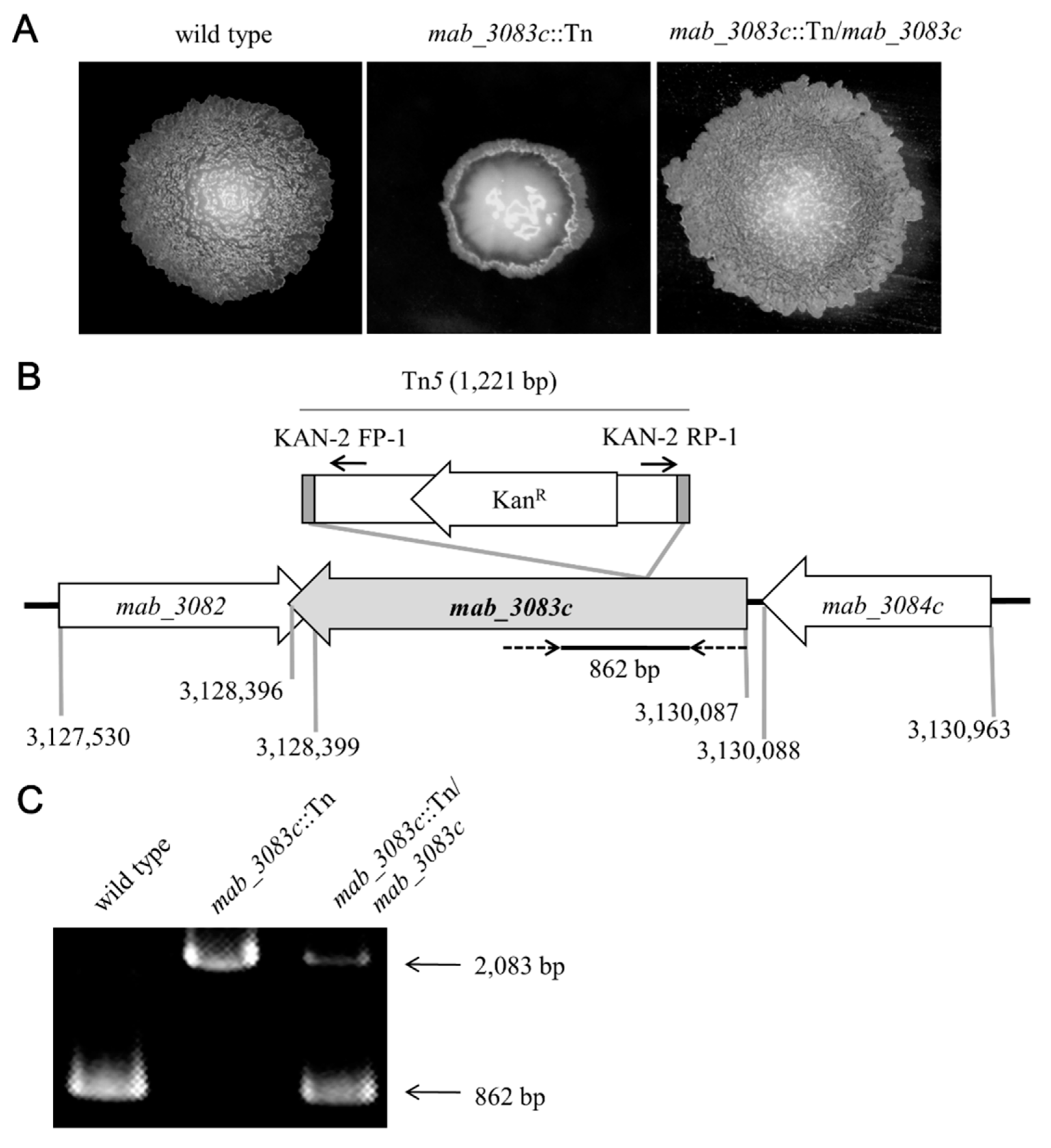
3.1. Characterization of the Mab_3083c::Tn Mutant
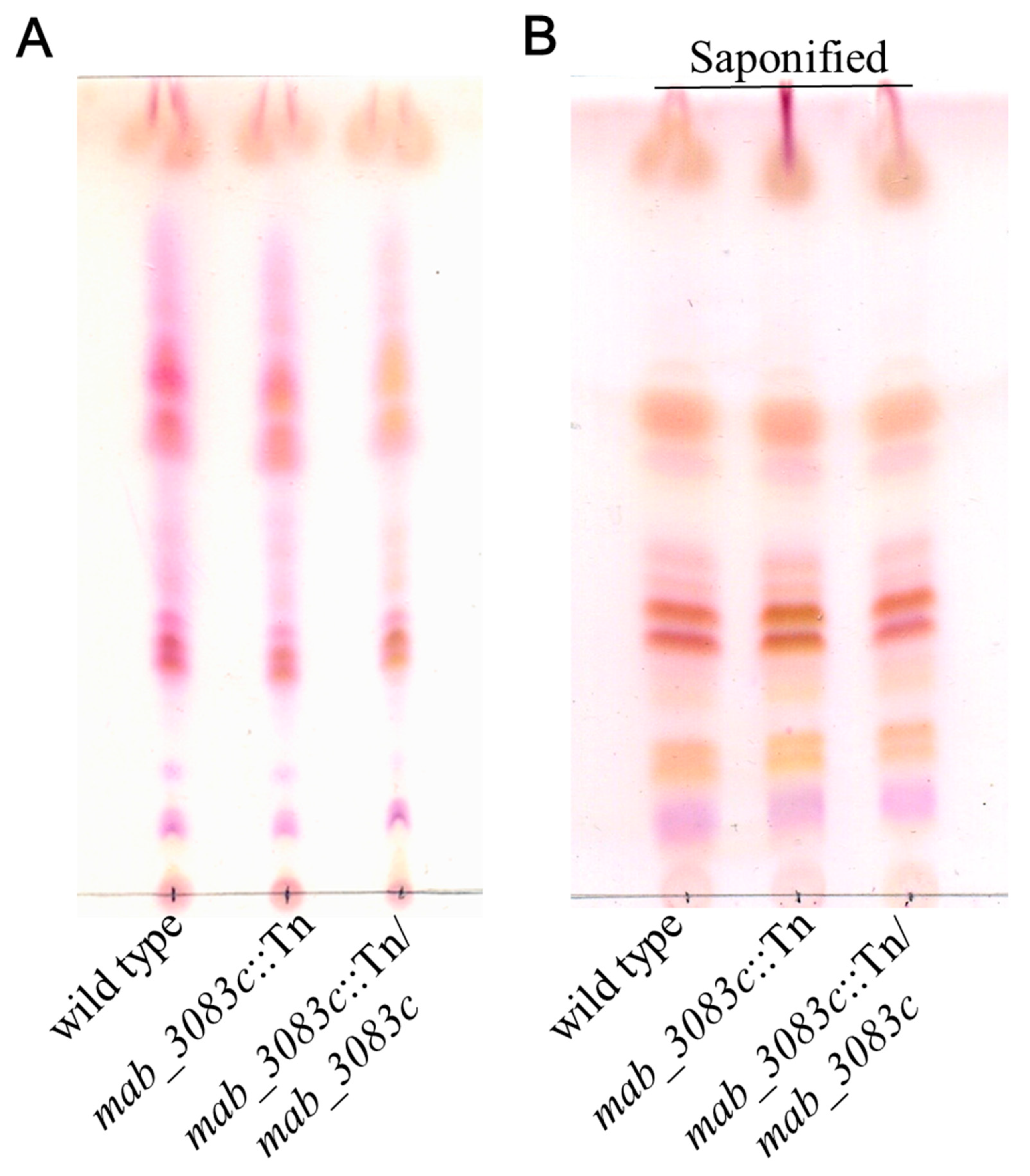
3.2. No Association of GPL with Colony Morphotype Switching of the mab_3083c::Tn Mutant
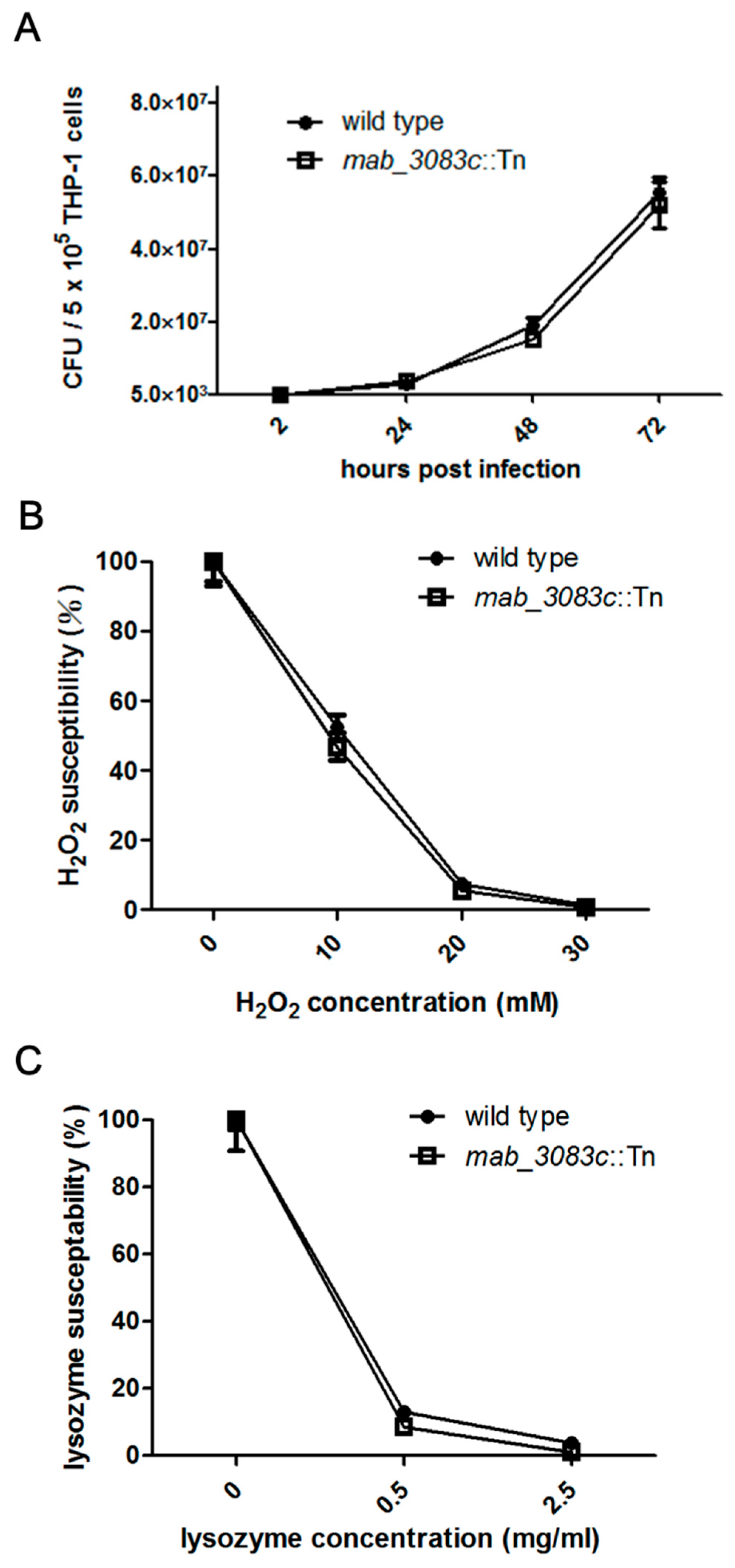
3.3. No Association of Mab_3083c with Intracellular Survival and Susceptibility to Hydrogen Peroxide and Lysozyme of M. abscessus
3.4. Higher Sliding Motility and Less Aggregation Capability of the Mab_3083c::Tn Mutant
3.5. Identification of Mab_3083c as a Homologue of Ribonuclease J
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Falkinham, J.O., III. Epidemiology of infection by nontuberculous mycobacteria. Clin. Microbiol. Rev. 1996, 9, 177–215. [Google Scholar] [CrossRef] [PubMed]
- McShane, P.J.; Glassroth, J. Pulmonary disease due to nontuberculous mycobacteria: Current state and new insights. Chest 2015, 148, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Griffith, D.E.; Girard, W.M.; Wallace, R.J. Clinical features of pulmonary disease caused by rapidly growing mycobacteria: An analysis of 154 patients. Am. Rev. Respir. Dis. 1993, 147, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Griffith, D.E.; Aksamit, T.; Iseman, M.; Olivier, K.; Ruoss, S.; Von Reyn, C.F.; Wallace, R.J.; Winthrop, K.; Brown-Elliott, B.A.; Catanzaro, A.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed]
- Nessar, R.; Cambau, E.; Reyrat, J.M.; Murray, A.; Gicquel, B. Mycobacterium abscessus: A new antibiotic nightmare. J. Antimicrob. Chemother. 2012, 67, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Sethiya, J.P.; Sowards, M.A.; Jackson, M.; North, E.J. MmpL3 inhibition: A new approach to treat nontuberculous mycobacterial infections. Int. J. Mol. Sci. 2020, 21, 6202. [Google Scholar] [CrossRef]
- Olivier, K.N.; Weber, D.J.; Wallace, R.J., Jr.; Edwards, L.J.; Chakraborti, S.; Knowles, M.R.; Faiz, A.R.; Lee, J.-H.; Zhang, Y.; Brown-Elliot, B.A.; et al. Nontuberculous mycobacteria. I: Amulticenter prevalence study in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2003, 167, 828–834. [Google Scholar] [CrossRef]
- Roux, A.-L.; Catherinot, E.; Gutierrez, C.; Vincent, V.; Fauroux, B.; Rottman, M.; Guillemot, D.; Gaillard, J.-L.; Jean-Louis Herrmann for the OMA Group; Ripoll, F.; et al. Multicenter study of prevalence of nontuberculous mycobacteria in patients with cystic fibrosis in france. J. Clin. Microbiol. 2009, 47, 4124–4128. [Google Scholar] [CrossRef]
- Huang, W.-C.; Chiou, C.-S.; Chen, J.-H.; Shen, G.-H. Molecular epidemiology of Mycobacterium abscessus infections in a subtropical chronic ventilatory setting. J. Med. Microbiol. 2010, 59, 1203–1211. [Google Scholar] [CrossRef][Green Version]
- Huang, H.-L.; Cheng, M.-H.; Lu, P.-L.; Shu, C.-C.; Wang, J.-Y.; Wang, J.-T.; Chong, I.-W.; Lee, L.-N. Epidemiology and predictors of NTM pulmonary infection in Taiwan-A retrospective, five-year multicenter study. Sci. Rep. 2017, 7, 16300. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J., Jr. The clinical presentation, diagnosis, and therapy of cutaneous and pulmonary infections due to the rapidly growing mycobacteria, M. fortuitum and M. chelonae. Clin. Chest Med. 1989, 10, 419–429. [Google Scholar]
- Li, B.; Ye, M.; Guo, Q.; Zhang, Z.; Yang, S.; Ma, W.; Yu, F.; Chu, H. Determination of MIC distribution and mechanisms of decreased susceptibility to bedaquiline among clinical isolates of mycobacterium abscessus. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Sha, W.; Weng, X.-H.; Xiao, H.-P.; He, G.-J. Investigation of drug-resistance to rifampin and rpoB gene sequence analysis of Mycobacterium abscessus. Zhonghua Jiehe He Huxi Zazhi. Chin. J. Tuberc. Respir. Dis. 2003, 26, 544–547. [Google Scholar]
- Alcaide, F.; Pfyffer, G.E.; Telenti, A. Role of embB in natural and acquired resistance to ethambutol in mycobacteria. Antimicrob. Agents Chemother. 1997, 41, 2270–2273. [Google Scholar] [CrossRef] [PubMed]
- Jarand, J.; Levin, A.; Zhang, L.; Huitt, G.; Mitchell, J.D.; Daley, C.L. Clinical and microbiologic outcomes in patients receiving treatment for mycobacterium abscessus pulmonary disease. Clin. Infect. Dis. 2011, 52, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Elidemir, O.; Heinle, J.; McKenzie, E.; Schecter, M.; Kaplan, S.; Dishop, M.; Kearney, D.; Mallory, G. Mycobacterium abscessusin cystic fibrosis lung transplant recipients: Report of 2 cases and risk for recurrence. Transpl. Infect. Dis. 2009, 11, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.; Ardito, F.; Fiscarelli, E.; La Sorda, M.; D’Argenio, P.; Ricciotti, G.; Fadda, G. Fatal pulmonary infection due to multidrug-resistant mycobacterium abscessus in a patient with cystic fibrosis. J. Clin. Microbiol. 2001, 39, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.C.; Gan, A.W.; Yam, A.; Tan, A.B.; Tay, S.C. Mycobacterium abscessus hand infections in immunocompetent fish handlers: Case report. J. Hand Surg. 2010, 35, 1142–1145. [Google Scholar] [CrossRef]
- Fregnan, G.B.; Smith, D.W. Description of various colony forms of mycobacteria. J. Bacteriol. 1962, 83, 819–827. [Google Scholar] [CrossRef]
- Recht, J.; Martínez, A.; Torello, S.; Kolter, R. Genetic analysis of sliding motility inmycobacterium smegmatis. J. Bacteriol. 2000, 182, 4348–4351. [Google Scholar] [CrossRef]
- Howard, S.T.; Rhoades, E.; Recht, J.; Pang, X.; Alsup, A.; Kolter, R.; Lyons, C.R.; Byrd, T.F. Spontaneous reversion of Mycobacterium abscessus from a smooth to a rough morphotype is associated with reduced expression of glycopeptidolipid and reacquisition of an invasive phenotype. Microbiology 2006, 152, 1581–1590. [Google Scholar] [CrossRef]
- Deshayes, C.; Laval, F.; Montrozier, H.; Daffé, M.; Etienne, G.; Reyrat, J.-M. A glycosyltransferase involved in biosynthesis of triglycosylated glycopeptidolipids in mycobacterium smegmatis: Impact on surface properties. J. Bacteriol. 2005, 187, 7283–7291. [Google Scholar] [CrossRef]
- Catherinot, E.; Roux, A.-L.; Rottman, M.; Gaillard, J.-L.; Herrmann, J.-L.; Macheras, E.; Hubert, D.; Matmar, M.; Dannhoffer, L.; Chinet, T.; et al. Acute respiratory failure involving an R variant of mycobacterium abscessus. J. Clin. Microbiol. 2008, 47, 271–274. [Google Scholar] [CrossRef]
- Byrd, T.F.; Lyons, C.R. Preliminary characterization of aMycobacterium abscessus mutant in human and murine models of infection. Infect. Immun. 1999, 67, 4700–4707. [Google Scholar] [CrossRef]
- Ripoll, F.; Deshayes, C.; Pasek, S.; Laval, F.; Beretti, J.-L.; Biet, F.; Risler, J.-L.; Daffé, M.; Etienne, G.; Gaillard, J.-L.; et al. Genomics of glycopeptidolipid biosynthesis in Mycobacterium abscessus and M. chelonae. Bmc Genom. 2007, 8, 114. [Google Scholar] [CrossRef]
- Gutiérrez, A.V.; Viljoen, A.; Ghigo, E.; Herrmann, J.-L.; Kremer, L. Glycopeptidolipids, a double-edged sword of the mycobacterium abscessus complex. Front. Microbiol. 2018, 9, 1145. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, E.R.; Archambault, A.S.; Greendyke, R.; Hsu, F.-F.; Streeter, C.; Byrd, T.F. Mycobacterium abscessusGlycopeptidolipids mask underlying cell wall phosphatidyl-myo-inositol mannosides blocking induction of human macrophage TNF-α by preventing interaction with TLR2a. J. Immunol. 2009, 183, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-H.; Shen, G.-H.; Lin, C.-H.; Liau, J.-R.; Lai, H.-C.; Hu, S.-T. Mab_3168c, a putative acetyltransferase, enhances adherence, intracellular survival and antimicrobial resistance of mycobacterium abscessus. PLoS ONE 2013, 8, e67563. [Google Scholar] [CrossRef]
- Li, H.; Havens, W.M.; Nibert, M.L.; Ghabrial, S.A. RNA sequence determinants of a coupled termination-reinitiation strategy for downstream open reading frame translation in helminthosporium victoriae virus 190s and other victoriviruses (family totiviridae). J. Virol. 2011, 85, 7343–7352. [Google Scholar] [CrossRef]
- Martínez, A.; Torello, S.; Kolter, R. Sliding motility in mycobacteria. J. Bacteriol. 1999, 181, 7331–7338. [Google Scholar] [CrossRef] [PubMed]
- Naka, T.; Nakata, N.; Fujiwara, N.; Maeda, S.; Yamamoto, R.; Doe, M.; Mizuno, S.; Niki, M.; Kobayashi, K.; Ogura, H.; et al. Structure and host recognition of serotype 13 glycopeptidolipid from mycobacterium intracellulare. J. Bacteriol. 2011, 193, 5766–5774. [Google Scholar] [CrossRef][Green Version]
- Chen, C.-C.; Tsai, S.-H.; Lu, C.-C.; Hu, S.-T.; Wu, T.-S.; Huang, T.-T.; Saïd-Sadier, N.; Ojcius, D.M.; Lai, H.-C. Activation of an NLRP3 inflammasome restricts mycobacterium kansasii infection. PLoS ONE 2012, 7, e36292. [Google Scholar] [CrossRef]
- Shin, D.-M.; Jeon, B.-Y.; Friedman, R.L.; Jo, E.-K.; Lee, H.-M.; Jin, H.S.; Yuk, J.-M.; Song, C.-H.; Lee, S.-H.; Lee, Z.-W.; et al. Mycobacterium tuberculosis eis regulates autophagy, inflammation, and cell death through redox-dependent signaling. PLoS Pathog. 2010, 6, e1001230. [Google Scholar] [CrossRef]
- Taverniti, V.; Forti, F.; Ghisotti, D.; Putzer, H. Mycobacterium smegmatis RNase J is a 5′-3′ exo-/endoribonuclease and both RNase J and RNase E are involved in ribosomal RNA maturation. Mol. Microbiol. 2011, 82, 1260–1276. [Google Scholar] [CrossRef]
- Hunt-Serracin, A.C.; Parks, B.J.; Boll, J.; Boutte, C.C. Mycobacterium abscessus cells have altered antibiotic tolerance and surface glycolipids in artificial cystic fibrosis sputum medium. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Halloum, I.; Carrère-Kremer, S.; Blaise, M.; Viljoen, A.; Bernut, A.; Le Moigne, V.; Vilchèze, C.; Guérardel, Y.; Lutfalla, G.; Herrmann, J.-L.; et al. Deletion of a dehydratase important for intracellular growth and cording renders rough Mycobacterium abscessus avirulent. Proc. Nat. Acad. Sci. USA 2016, 113, E4228–E4237. [Google Scholar] [CrossRef]
- Davidson, L.B.; Nessar, R.; Kempaiah, P.; Perkins, D.J.; Byrd, T.F. Mycobacterium abscessus glycopeptidolipid prevents respiratory epithelial TLR2 signaling as measured by HβD2 gene expression and IL-8 release. PLoS ONE 2011, 6, e29148. [Google Scholar] [CrossRef]
- Figaro, S.; Durand, S.; Gilet, L.; Cayet, N.; Sachse, M.; Condon, C. Bacillus subtilis mutants with knockouts of the genes encoding ribonucleases RNAse Y and rnase J1 are viable, with major defects in cell morphology, sporulation, and competence. J. Bacteriol. 2013, 195, 2340–2348. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Leong, V.; Ortega, J.; Elliot, M.A. Development, antibiotic production, and ribosome assembly in streptomyces venezuelae are impacted by RNAse J and RNAse III deletion. J. Bacteriol. 2014, 196, 4253–4267. [Google Scholar] [CrossRef]
- Bugrysheva, J.V.; Scott, J.R. Regulation of virulence gene expression in Streptococcus pyogenes: Determinants of differential mRNA decay. Rna Biol. 2010, 7, 569–572. [Google Scholar] [CrossRef][Green Version]
- Madhugiri, R.; Evguenieva-Hackenberg, E. RNase J is involved in the 5′-end maturation of 16S rRNA and 23S rRNA inSinorhizobium meliloti. FEBS Lett. 2009, 583, 2339–2342. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Condon, C.; Bechhofer, D.H. Regulated RNA stability in the Gram positives. Curr. Opin. Microbiol. 2011, 14, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.K.; Kushner, S.R. Regulation of mRNA decay in bacteria. Annu. Rev. Microbiol. 2016, 70, 25–44. [Google Scholar] [CrossRef]
- Even, S.; Pellegrini, O.; Zig, L.; Labas, V.; Vinh, J.; Brechemmier-Baey, D.; Putzer, H. Ribonucleases J1 and J2: Two novel endoribonucleases in B. subtilis with functional homology to E. coli RNase E. Nucleic Acids Res. 2005, 33, 2141–2152. [Google Scholar] [CrossRef]
- Dorléans, A.; De La Sierra-Gallay, I.L.; Piton, J.; Zig, L.; Gilet, L.; Putzer, H.; Condon, C. Molecular basis for the recognition and cleavage of RNA by the bifunctional 5′–3′ exo/Endoribonuclease RNAse J. Structure 2011, 19, 1252–1261. [Google Scholar] [CrossRef]
- Mäder, U.; Zig, L.; Kretschmer, J.; Homuth, G.; Putzer, H. mRNA processing by RNases J1 and J2 affects Bacillus subtilis gene expression on a global scale. Mol. Microbiol. 2008, 70, 183–196. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.-Y.; Tsai, S.-H.; Chen, J.-W.; Wang, Y.-C.; Hu, S.-T.; Chen, Y.-Y. Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus. Microorganisms 2021, 9, 676. https://doi.org/10.3390/microorganisms9040676
Liu T-Y, Tsai S-H, Chen J-W, Wang Y-C, Hu S-T, Chen Y-Y. Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus. Microorganisms. 2021; 9(4):676. https://doi.org/10.3390/microorganisms9040676
Chicago/Turabian StyleLiu, Ting-Yu, Sheng-Hui Tsai, Jenn-Wei Chen, Yu-Ching Wang, Shiau-Ting Hu, and Yih-Yuan Chen. 2021. "Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus" Microorganisms 9, no. 4: 676. https://doi.org/10.3390/microorganisms9040676
APA StyleLiu, T.-Y., Tsai, S.-H., Chen, J.-W., Wang, Y.-C., Hu, S.-T., & Chen, Y.-Y. (2021). Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus. Microorganisms, 9(4), 676. https://doi.org/10.3390/microorganisms9040676