
Maintaining Digestive Health in Diabetes: The Role of the Gut Microbiome and the Challenge of Functional Foods

 ,
, 

 , ,
, ,  , ,
, , 
 ,
,  and
and 
Abstract
1. Introduction-Diabetes as a Disease
2. Microbiome and Diabetes
2.1. Microbiome at a Glance
2.2. Mechanisms Connecting Gut Microbiota with Occurrence of T2DM
2.2.1. Short-Chain Fatty Acid Contribution
2.2.2. Lipid Metabolism
2.2.3. Disruption of Intestinal Barrier Function and Endotoxemia
2.2.4. Inflammation and the Immune System
2.2.5. Microbiota Traits in DM
3. Functional Foods and Diabetes
3.1. The Challenge of Functional Foods
3.2. Animal Studies
3.3. Human Studies
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGE | advanced glycosylation end products |
| BMS | burning mouth syndrome |
| D | type 2 iodothyronine deiodinase |
| DAG | diacylglycerol |
| DSHEA | Dietary Supplement Health and Education Act system |
| Epac | Exchange protein directly activated by cAMP |
| FBG | fasting blood glucose |
| FFAs | free fatty acids |
| FOSHU | Foods for Specialized Health Use |
| GDM | Gestational Diabetes Mellitus |
| GLP-1 | glucagon-like peptide-1 |
| TGR5 | G-protein-coupled bile acid receptor 5 |
| HFD | high fat diet |
| IKKb | b kinase inhibitor |
| IL | Interleukin |
| JNK | c-Jun-N-terminal kinases |
| LAB | lactic acid bacteria |
| LIP | liver inhibitory protein |
| LPL | lipoprotein lipase |
| LPS | lipopolysaccharides |
| MAPK | mitogen-activated protein kinase |
| PRRs | pattern recognition receptors |
| PYY | peptide tyrosine-tyrosine |
| SCFAs | Short-chain fatty acids |
| T1DM | type 1 diabetes mellitus |
| T2DM | type 2 diabetes mellitus |
| T3 | active tri-iodothyronine |
| TLR4 | Toll-like receptor 4 |
| TMA | trimethylamine |
| TNF | Tumor necrosis facto |
| VLDL | very low-density lipoproteins |
| WGS | Whole Genome Shotgun |
| XR | Farnesol X-receptor |
References
- World Health Organization. Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 19 November 2020).
- International Diabetes Federation. Facts and Figures. Available online: https://www.idf.org/aboutdiabetes/what-is-diabetes/facts-figures.html (accessed on 26 December 2020).
- Petersen, M.C.; Shulman, G.I. Mechanisms of insulin action and insulin resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Guo, C.; Chen, W.D.; Wang, Y.D. TGR5, Not only a metabolic regulator. Front. Physiol. 2016, 7, 646. [Google Scholar] [CrossRef]
- WebMD. Types of Diabetes Mellitus. Available online: https://www.webmd.com/diabetes/guide/types-of-diabetes-mellitus (accessed on 19 November 2020).
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Hasain, Z.; Mokhtar, N.M.; Kamaruddin, N.A.; Ismail, N.A.M.; Razalli, N.H.; Gnanou, J.V.; Ali, R.A.R. Gut Microbiota and Gestational Diabetes Mellitus: A Review of Host-Gut Microbiota Interactions and Their Therapeutic Potential. Front. Cell. Infect. Microbiol. 2020, 10, 188. [Google Scholar] [CrossRef]
- Bellamy, L.; Casas, J.P.; Hingorani, A.D.; Williams, D. Type 2 diabetes mellitus after gestational diabetes: A systematic review and meta-analysis. Lancet 2009, 373, 1773–1779. [Google Scholar] [CrossRef]
- Boulangé, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef]
- Sell, H.; Habich, C.; Eckel, J. Adaptive immunity in obesity and insulin resistance. Nat. Rev. Endocrinol. 2012, 8, 709–716. [Google Scholar] [CrossRef]
- Yoon, J.W.; Jun, H.S. Autoimmune destruction of pancreatic beta cells. Am. J. Ther. 2005, 12, 580–591. [Google Scholar] [CrossRef]
- Karlsson, F.A.; Berne, C.; Björk, E.; Kullin, M.; Li, Z.; Ma, J.Y.; Schölin, A.; Zhao, L. Beta-cell activity and destruction in type 1 diabetes. Ups. J. Med. Sci. 2000, 105, 85–95. [Google Scholar] [CrossRef]
- Filippi, C.M.; von Herrath, M.G. Viral trigger for type 1 diabetes: Pros and cons. Diabetes 2008, 57, 2863–2871. [Google Scholar] [CrossRef]
- Beik, P.; Ciesielska, M.; Kucza, M.; Kurczewska, A.; Kuźmińska, J.; Maćkowiak, B.; Niechciał, E. Prevention of Type 1 Diabetes: Past Experiences and Future Opportunities. J. Clin. Med. 2020, 9, 2805. [Google Scholar] [CrossRef]
- Stefanaki, C.; Peppa, M.; Mastorakos, G.; Chrousos, G.P. Examining the gut bacteriome, virome, and mycobiome in glucose metabolism disorders: Are we on the right track? Metab. Clin. Exp. 2017, 73, 52–66. [Google Scholar] [CrossRef]
- World Health Organization. Global Report on Diabetes. Available online: https://www.who.int/publications-detail-redirect/9789241565257 (accessed on 19 November 2020).
- Mauri-Obradors, E.; Estrugo-Devesa, A.; Jané-Salas, E.; Viñas, M.; López-López, J. Oral manifestations of Diabetes Mellitus. A systematic review. Med. Oral. Patol. Oral. Cir. Bucal. 2017, 22, e586–e594. [Google Scholar] [CrossRef]
- Connolly, V.; Unwin, N.; Sherriff, P.; Bilous, R.; Kelly, W. Diabetes prevalence and socioeconomic status: A population based study showing increased prevalence of type 2 diabetes mellitus in deprived areas. J. Epidemiol. Community Health 2000, 54, 173–177. [Google Scholar] [CrossRef]
- Rayappa, Ph.; Raju, K.; Kapur, A.; Björk, S.; Sylvest, C.; Kumar, D. The Impact of socio-economic factors on diabetes care. Int. J. Diab. Dev. Ctries. 1999, 19, 7–15. [Google Scholar]
- Anjana, R.M.; Deepa, M.; Pradeepa, R.; Mahanta, J.; Narain, K.; Das, H.K.; Adhikari, P.; Rao, P.V.; Saboo, B.; Kumar, A.; et al. Prevalence of diabetes and prediabetes in 15 states of India: Results from the ICMR–INDIAB population-based cross-sectional study. Lancet Diabetes Endocrinol. 2017, 5, 585–596. [Google Scholar] [CrossRef]
- Statista. Prevalence of Diabetes in Europe in 2019. Available online: https://www.statista.com/statistics/1081006/prevalence-of-diabetes-in-europe/ (accessed on 19 November 2020).
- Jiménez, E.; Fernández, L.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef]
- Bezirtzoglou, E.; Romond, C. Effect of the feeding practices on the establishment of bacterial interactions in the intestine of the newborn delivered by cesarean section. J. Perinat. Med. 1990, 18, 139–143. [Google Scholar] [CrossRef]
- Gabriel, I.; Olejek, A.; Stencel-Gabriel, K.; Wielgoś, M. The influence of maternal vaginal flora on the intestinal colonization in newborns and 3-month-old infants. J. Matern. Fetal Neonatal. Med. 2018, 31, 1448–1453. [Google Scholar] [CrossRef]
- Mueller, N.T.; Whyatt, R.; Hoepner, L.; Oberfield, S.; Dominguez-Bello, M.G.; Widen, E.M.; Hassoun, A.; Perera, F.; Rundle, A. Prenatal exposure to antibiotics, cesarean section and risk of childhood obesity. Int. J. Obes. 2015, 39, 665–670. [Google Scholar] [CrossRef]
- NIH Human Microbiome Project—Home. Available online: https://www.hmpdacc.org/ (accessed on 29 November 2020).
- MetaHIT Consortium (Metagenomics of the Human Intestinal Tract Consortium)—Welcome Sanger Institute. Available online: https://www.sanger.ac.uk/resources/downloads/bacteria/metahit/ (accessed on 29 November 2020).
- Structure, Function and Diversity of the Healthy Human Microbiome. Nature. Available online: https://www.nature.com/articles/nature11234 (accessed on 29 November 2020).
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef]
- Brunkwall, L.; Orho-Melander, M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: From current human evidence to future possibilities. Diabetologia 2017, 60, 943–951. [Google Scholar] [CrossRef]
- Song, S.J.; Dominguez-Bello, M.G.; Knight, R. How delivery mode and feeding can shape the bacterial community in the infant gut. Can. Med. Assoc. J. 2013, 185, 373–374. [Google Scholar] [CrossRef]
- Lazar, V.; Ditu, L.M.; Pircalabioru, G.G.; Picu, A.; Petcu, L.; Cucu, N.; Chifiriuc, M.C. Gut Microbiota, Host Organism, and Diet Trialogue in Diabetes and Obesity. Front. Νutr. 2019, 6, 21. [Google Scholar] [CrossRef]
- Marrs, T.; Jo, J.H.; Perkin, M.R.; Rivett, D.W.; Witney, A.A.; Bruce, K.D.; Logan, K.; Craven, J.; Radulovic, S.; Versteeg, S.A.; et al. Gut microbiota development during infancy: Impact of introducing allergenic foods. J. Allergy Clin. Immunol. 2021, 147, 613–621. [Google Scholar] [CrossRef]
- Sarkar, A.; Yoo, J.Y.; Valeria Ozorio Dutra, S.; Morgan, K.H.; Groer, M. The Association between Early-Life Gut Microbiota and Long-Term Health and Diseases. J. Clin. Med. 2021, 10, 459. [Google Scholar] [CrossRef]
- Filippo, C.D.; Cavalieri, D.; Paola, M.D.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.P.; Li, H.B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Bufford, J.D.; Gern, J.E. The hygiene hypothesis revisited. Immunol. Allergy Clin. North. Am. 2005, 25, 247–262. [Google Scholar] [CrossRef]
- Patki, A. Eat dirt and avoid atopy: The hygiene hypothesis revisited. Indian J. Dermatol. Venereol. Leprol. 2007, 73, 2–4. [Google Scholar] [CrossRef]
- Stiemsma, L.T.; Reynolds, L.A.; Turvey, S.E.; Finlay, B.B. The hygiene hypothesis: Current perspectives and future therapies. Immunotargets Ther. 2015, 4, 143–157. [Google Scholar] [CrossRef]
- Larsson, E.; Tremaroli, V.; Lee, Y.S.; Koren, O.; Nookaew, I.; Fricker, A.; Nielsen, J.; Ley, R.E.; Backhed, F. Analysis of gut microbial regulation of host gene expression along the length of the gut and regulation of gut microbial ecology through MyD88. Gut Microb. 2012, 61, 1124–1131. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR–S6K pathway. Mucosal. Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef]
- Acharya, A.B.; Thakur, S.; Muddapur, M.V. Evaluation of serum interleukin-10 levels as a predictor of glycemic alteration in chronic periodontitis and type 2 diabetes mellitus. J. Indian Soc. Periodontol. 2015, 19, 388–392. [Google Scholar] [CrossRef]
- Wang, Z.; Shen, X.-H.; Feng, W.-M.; Ye, G.-F.; Qiu, W.; Li, B. Analysis of inflammatory mediators in prediabetes and newly diagnosed Type 2 diabetes patients. J. Diabetes Res. 2016, 2016, 7965317. [Google Scholar]
- Gérard, P. Gut microbiota and obesity. Cell Mol. Life Sci. 2016, 73, 147–162. [Google Scholar] [CrossRef]
- Bensalem, A.; Murtaza, B.; Hichami, A.; Khan, A.S.; Oulamara, H.; Merlen, G.; Berrichi, M.; Agli, A.N.; Tordjmann, T.; Khan, N.A. Bile acid receptor TGR5 is critically involved in preference for dietary lipids and obesity. J. Nutr. Biochem. 2020, 76, 108298. [Google Scholar] [CrossRef]
- Kumar, D.P.; Asgharpour, A.; Mirshahi, F.; Park, S.H.; Liu, S.; Imai, Y.; Nadler, J.L.; Grider, J.R.; Murthy, K.S.; Sanyal, A.J. Activation of transmembrane bile acid receptor TGR5 modulates pancreatic islet α cells to promote glucose homeostasis. J. Biol. Chem. 2016, 291, 6626–6640. [Google Scholar] [CrossRef]
- Jiao, Y.; Lu, Y.; Li, X.Y. Farnesoid X receptor: A master regulator of hepatic triglyceride and glucose homeostasis. Acta Pharmacol. Sin. 2015, 36, 44–50. [Google Scholar] [CrossRef]
- McMillin, M.; Grant, S.; Frampton, G.; Petrescu, A.D.; Kain, J.; Williams, E.; Haines, R.; Canady, L.; DeMorrow, S. FXR-Mediated cortical cholesterol accumulation contributes to the pathogenesis of type A hepatic encephalopathy. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 47–63. [Google Scholar] [CrossRef]
- Xu, Y.; Li, F.; Zalzala, M.; Xu, J.; Gonzalez, F.J.; Adorini, L.; Lee, Y.K.; Yin, L.; Zhang, Y. Farnesoid X receptor activation increases reverse cholesterol transport by modulating bile acid composition and cholesterol absorption in mice. Hepatology 2016, 64, 1072–1085. [Google Scholar] [CrossRef]
- Perino, A.; Pols, T.W.; Nomura, M.; Stein, S.; Pellicciari, R.; Schoonjans, K. TGR5 reduces macrophage migration through mTOR-induced C/EBPβ differential translation. J. Clin. Investig. 2014, 124, 5424–5436. [Google Scholar] [CrossRef]
- Kumar, D.P.; Rajagopal, S.; Mahavadi, S.; Mirshahi, F.; Grider, J.R.; Murthy, K.S.; Sanyal, A.J. Activation of transmembrane bile acid receptor TGR5 stimulates insulin secretion in pancreatic cells. Biochem. Biophys. Res. Commun. 2012, 2012. 427, 600–605. [Google Scholar] [CrossRef]
- Chen, X.; Lou, G.; Meng, Z.; Huang, W. TGR5: A novel target for weight maintenance and glucose metabolism. Exp. Diabetes Res. 2011, 853501. [Google Scholar] [CrossRef]
- Meng, Y.; Li, X.; Zhang, J.; Wang, C.; Lu, F. Effects of different diets on microbiota in the small intestine mucus and weight regulation in rats. Sci. Rep. 2019, 9, 8500. [Google Scholar] [CrossRef]
- Brown, B.I. Nutritional management of metabolic endotoxemia: A clinical review. Altern. Ther. Health Med. 2017, 23, 42–54. [Google Scholar]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef]
- André, P.; Laugerette, F.; Féart, C. Metabolic Endotoxemia: A Potential Underlying Mechanism of the Relationship between Dietary Fat Intake and Risk for Cognitive Impairments in Humans? Nutrients 2019, 11, 1887. [Google Scholar] [CrossRef]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid-induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef]
- Cheru, L.; Saylor, C.F.; Lo, J. Gastrointestinal barrier breakdown and adipose tissue inflammation. Curr. Obes. Rep. 2019, 8, 165–174. [Google Scholar] [CrossRef]
- Sánchez de Medina, F.; Romero-Calvo, I.; Mascaraque, C.; Martínez-Augustin, O. Intestinal inflammation and mucosal barrier function. Inflamm. Bowel Dis. 2014, 20, 2394–2404. [Google Scholar] [CrossRef]
- Genser, L.; Aguanno, D.; Soula, H.A.; Dong, L.; Trystram, L.; Assmann, K.; Salem, J.E.; Vaillant, J.C.; Oppert, J.M.; Laugerette, F.; et al. Increased jejunal permeability in human obesity is revealed by a lipid challenge and is linked to inflammation and type 2 diabetes. J. Pathol. 2018, 246, 217–230. [Google Scholar] [CrossRef]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef]
- Honda, K.; Moto, M.; Uchida, N.; He, F.; Hashizume, N. Anti-diabetic effects of lactic acid bacteria in normal and type 2 diabetic mice. J. Clin. Biochem. Nutr. 2012, 51, 96–101. [Google Scholar] [CrossRef]
- Wang, G.; Si, Q.; Yang, S.; Jiao, T.; Zhu, H.; Tian, P.; Wang, L.; Li, X.; Gong, L.; Zhao, J.; et al. Lactic acid bacteria reduce diabetes symptoms in mice by alleviating gut microbiota dysbiosis and inflammation in different manners. Food Funct. 2020, 11, 5898–5914. [Google Scholar] [CrossRef]
- McArdle, M.A.; Finucane, O.M.; Connaughton, R.M.; McMorrow, A.M.; Roche, H.M. Mechanisms of Obesity-Induced Inflammation and Insulin Resistance: Insights into the Emerging Role of Nutritional Strategies. Front. Endocrinol. 2013, 2013 4, 1–23. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nagai, Y.; Takatsu, K. Activation and Regulation of the Pattern Recognition Receptors in Obesity-Induced Adipose Tissue Inflammation and Insulin Resistance. Nutrients 2013, 5, 3757–3778. [Google Scholar] [CrossRef]
- Schenk, S.; Saberi, M.; Olefsky, J.M. Insulin sensitivity: Modulation by nutrients and inflammation. J. Clin. Invest. 2008, 118, 2992–3002. [Google Scholar] [CrossRef]
- Janssens, S.; Beyaert, R. Role of Toll-Like Receptors in Pathogen Recognition. Clin. Microbiol. Rev. 2003, 16, 637–646. [Google Scholar] [CrossRef]
- Mogensen, T.H. Pathogen Recognition and Inflammatory Signaling in Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108, 4554–4561. [Google Scholar] [CrossRef]
- Carvalho, B.M.; Guadagnini, D.; Tsukumo, D.M.L.; Schenka, A.A.; Latuf-Filho, P.; Vassallo, J.; Dias, J.C.; Kubota, L.T.; Carvalheira, J.B.C.; Saad, M.J.A. Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia 2012, 55, 2823–2834. [Google Scholar] [CrossRef]
- Caricilli, A.M.; Picardi, P.K.; de Abreu, L.L.; Ueno, M.; Prada, P.O.; Ropelle, E.R. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLOS Biol. 2011, 9, e1001212. [Google Scholar] [CrossRef]
- Dasu, M.R.; Devaraj, S.; Park, S.; Jialal, I. Increased Toll-Like Receptor (TLR) Activation and TLR Ligands in Recently Diagnosed Type 2 Diabetic Subjects. Diabetes Care 2010, 33, 861–868. [Google Scholar] [CrossRef]
- Pellegrini, S.; Sordi, V.; Bolla, A.M.; Saita, D.; Ferrarese, R.; Canducci, F.; Clementi, M.; Invernizzi, F.; Mariani, A.; Bonfanti, R.; et al. Duodenal Mucosa of Patients With Type 1 Diabetes Shows Distinctive Inflammatory Profile and Microbiota. J. Clin. Endocrinol. Metab. 2017, 102, 1468–1477. [Google Scholar] [CrossRef]
- Cinek, O.; Kramna, L.; Mazankova, K.; Odeh, R.; Alassaf, A.; Ibekwe, M.U.; Ahmadov, G.; Elmahi, B.M.E.; Mekki, H.; Lebl, J.; et al. The bacteriome at the onset of type 1 diabetes: A study from four geographically distant African and Asian countries. Diabetes Res. Clin. Pract. 2018, 144, 51–62. [Google Scholar] [CrossRef]
- Leiva-Gea, I.; Sánchez-Alcoholado, L.; Martín-Tejedor, B.; Castellano-Castillo, D.; Moreno-Indias, I.; Urda-Cardona, A.; Tinahones, F.J.; Fernández-García, J.C.; Queipo-Ortuño, M.I. Gut Microbiota Differs in Composition and Functionality Between Children With Type 1 Diabetes and MODY2 and Healthy Control Subjects: A Case-Control Study. Diabetes Care 2018, 41, 2385–2395. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sorensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Zheng, C.; Chen, J.; Chu, F.; Zhu, J.; Jin, T. Inflammatory Role of TLR-MyD88 Signaling in Multiple Sclerosis. Front. Mol. Neurosci. 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Valladares, R.; Sankar, D.; Li, N.; Williams, E.; Lai, K.K.; Abdelgeliel, A.S.; Gonzalez, C.F.; Wasserfall, C.H.; Larkin, J.; Schatz, D. Lactobacillus johnsonii N6.2 Mitigates the Development of Type 1 Diabetes in BB-DP Rats. PLoS ONE 2010, 5, e10507. [Google Scholar] [CrossRef]
- Devaraj, S.; Hemarajata, P.; Versalovic, J. The Human Gut Microbiome and Body Metabolism: Implications for Obesity and Diabetes. Clin. Chem. 2013, 59, 617–628. [Google Scholar] [CrossRef]
- Schmidt, A.M. Diabetes Mellitus and Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1–11. [Google Scholar] [CrossRef]
- Liu, D.; Liu, L.; Hu, Z.; Song, Z.; Wang, Y.; Chen, Z. Evaluation of the oxidative stress-related genes ALOX5, ALOX5AP, GPX1, GPX3 and MPO for contribution to the risk of type 2 diabetes mellitus in the Han Chinese population. Diab. Vasc. Dis. Res. 2018, 15, 336–339. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chuang, L.M. The role of oxidative stress in the pathogenesis of type 2 diabetes: From molecular mechanism to clinical implication. Am. J. Transl. Res. 2010, 2, 316–331. [Google Scholar]
- Tilg, H.; Kaser, A. Gut microbiome, obesity, and metabolic dysfunction. J. Clin. Investig. 2011, 121, 2126–2132. [Google Scholar] [CrossRef]
- Everard, A.; Cani, P.D. Diabetes, obesity and gut microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 73–83. [Google Scholar] [CrossRef]
- Monjiote, D.P.; Leo, E.E.M.; Campos, M.R.S. Functional and Biological Potential of Bioactive Compounds in Foods for the Dietary Treatment of Type 2 Diabetes Mellitus. Functional Food—Improve Health through Adequate Food; Hueda, M.C., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Mancebo, F. Urbanism. In Encyclopedia of Community: From the Village to the Virtual World; Sage: New York, NY, USA, 2003; pp. 1428–1433. [Google Scholar]
- Stavropoulou, E.; Tsigalou, C.; Bezirtzoglou, E. Functions of the Human Intestinal Microbiota in Relation to Functional Foods. Erciyes Med. J. 2018, 40, 188–193. [Google Scholar] [CrossRef]
- Bezirtzoglou, E.; Stavropoulou, E. Immunology and probiotic impact of the newborn and young children intestinal microflora. Anaerobe 2011, 17, 369–374. [Google Scholar] [CrossRef]
- Food & Nutrition Magazine. The History and Health Benefits of Fermented Food. Available online: https://foodandnutrition.org/winter-2012/history-health-benefits-fermented-food/ (accessed on 29 November 2020).
- Smid, E.J.; Hugenholtz, J. Functional Genomics for Food Fermentation Processes. Annu. Rev. Food Sci. Technol. 2010, 1, 497–519. [Google Scholar] [CrossRef]
- Montet, D.; Ray, R. Fermented Foods: Part I: Biochemistry & Biotechnology. Fermented Foods; Routledge: Abingdon, UK, 2015. [Google Scholar]
- Adesulu-Dahunsi, A.T.; Dahunsi, S.O.; Olayanju, A. Synergistic microbial interactions between lactic acid bacteria and yeasts during production of Nigerian indigenous fermented foods and beverages. Food Control 2020, 110, 106963. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics as a weapon in the fight against COVID-19. Front. Nutr. 2020, 7, 614986. [Google Scholar] [CrossRef]
- Elbron, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Iwatani, S.; Yamamoto, N. Functional food products in Japan: A review. Food Sci. Hum. Well. 2019, 8, 96–101. [Google Scholar] [CrossRef]
- Nutraceutical and Functional Food Regulations in the United States and around the World; Elsevier: Amsterdam, The Netherlands, 2019. [CrossRef]
- Fukushima, Y.; Kawata, Y.; Hara, H.; Terada, A.; Mitsuoka, T. Effect of a probiotic formula on intestinal immunoglobulin A production in healthy children. Int. J. Food Microbiol. 1998, 42, 39–44. [Google Scholar] [CrossRef]
- Küster-Boluda, I.; Vidal-Capilla, I. Consumer attitudes in the election of functional foods. Spanish J. Mark. ESIC 2017, 21, 65–79. [Google Scholar] [CrossRef]
- Steinhagen, F.; Schmidt, S.V.; Schewe, J.C.; Peukert, K.; Klinman, D.M.; Bode, C. Immunotherapy in sepsis—Brake or accelerate? Pharmacol. Ther. 2020, 208, 107476. [Google Scholar] [CrossRef]
- Mirmiran, P.; Bahadoran, Z.; Azizi, F. Functional foods-based diet as a novel dietary approach for management of type 2 diabetes and its complications: A review. World J. Diabetes 2014, 5, 267–281. [Google Scholar] [CrossRef]
- Alkhatib, A.; Tsang, C.; Tiss, A.; Bahorun, T.; Arefanian, H.; Barake, R. Functional Foods and Lifestyle Approaches for Diabetes Prevention and Management. Nutrients 2017, 9, 1310. [Google Scholar] [CrossRef]
- Tsigalou, C.; Konstantinidis, T.; Paraschaki, A.; Stavropoulou, E.; Voidarou, C.; Bezirtzoglou, E. Mediterranean Diet as a Tool to Combat Inflammation and Chronic Diseases. An Overview. Biomedicines 2020, 8, 201. [Google Scholar] [CrossRef]
- GMFH—Best of Nutrition Diet. Available online: https://www.gutmicrobiotaforhealth.com/wp-content/uploads/2019/03/GMFH-BESTOF-NUTRITION-DIET.pdf (accessed on 29 November 2020).
- Tomova, A.; Bukovsky, I.; Rembert, E.; Yonas, W.; Alwarith, J.; Barnard, N.; Kahleova, H. The Effects of Vegetarian and Vegan Diets on Gut Microbiota. Front. Nutr. 2019, 6, 47. [Google Scholar] [CrossRef]
- Tonucci, L.B.; Dos Santos, K.M.O.; Ferreira, C.L.D.L.F.; Ribeiro, S.M.R.; De Oliveira, L.L.; Martino, H.S.D. Gut microbiota and probiotics: Focus on diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2017, 57, 2296–2309. [Google Scholar] [CrossRef]
- Yao, K.; Zeng, L.; He, Q.; Wang, W.; Lei, J.; Zou, X. Effect of Probiotics on Glucose and Lipid Metabolism in Type 2 Diabetes Mellitus: A Meta-Analysis of 12 Randomized Controlled Trials. Med. Sci. Monit. 2017, 23, 3044–3053. [Google Scholar] [CrossRef]
- Silva, J.C.P.; Jones, J.G. Improving Metabolic Control Through Functional Foods. Curr. Med. Chem. 2020, 26, 3424–3438. [Google Scholar] [CrossRef]
- Kerry, R.G.; Das, G.; Golla, U.; Rodriguez-Torres, M.D.; Shin, H.; Patra, J.K. Engineered probiotic and prebiotic nutraceutical supplementations in combating non-communicable disorders: A review. Curr. Pharm. Biotechnol. 2020, 21, 33050862. [Google Scholar] [CrossRef]
- King, A.J. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef]
- Al-Awar, A.; Kupai, K.; Veszelka, M.; Szucs, G.; Attieh, Z.; Murlasits, Z.; Varga, C. Experimental Diabetes Mellitus in Different Animal Models. J. Diabetes Res. 2016, 2016, 9051426. [Google Scholar] [CrossRef]
- Xue, M.; Liua, Y.; Xu, H.; Zhou, Z.; Ma, Y.; Sun, T.; Liu, M.; Zhang, H.; Liang, H. Propolis modulates the gut microbiota and improves the intestinal mucosal barrier function in diabetic rats. Biomed. Pharmacother. 2019, 118, 109393. [Google Scholar] [CrossRef]
- Terzo, S.; Mulè, F.; Caldara, G.F.; Baldassano, S.; Puleio, R.; Vitale, M.; Cassata, G.; Ferrantelli, V.; Amato, A. Pistachio Consumption Alleviates Inflammation and Improves Gut Microbiota Composition in Mice Fed a High-Fat Diet. Int. J. Mol. Sci. 2020, 21, 365. [Google Scholar] [CrossRef]
- Yanni, AE.; Mitropoulou, G.; Prapa, I.; Agrogiannis, G.; Kostomitsopoulos, N.; Bezirtzoglou, E.; Kourkoutas, Y.; Karathanos, V.T. Functional modulation of gut microbiota in diabetic rats following dietary intervention with pistachio nuts (Pistacia vera L.). Metab. Open 2020, 7, 100040. [Google Scholar] [CrossRef]
- Zhong, Y.; Nyman, M.; Fak, F. Modulation of gut microbiota in rats fed high-fat diets by processing whole-grain barley to barley malt. Mol. Nutr. Food Res. 2015, 59, 2066–2076. [Google Scholar]
- Bhandarkar, N.S.; Brown, L.; Panchal, S.K. Chlorogenic acid attenuates high-carbohydrate, high-fat diet-induced cardiovascular, liver, and metabolic changes in rats. Nutr. Res. 2018, 62, 78–88. [Google Scholar] [CrossRef]
- Zhang, H.H.; Liu, J.; Lv, Y.J.; Jiang, Y.I.; Pan, J.X.; Zhu, Y.J.; Huang, M.G.; Zhang, S.K. Changes in Intestinal Microbiota of Type 2 Diabetes in Mice in Response to Dietary Supplementation With Instant Tea or Matcha. Can. J. Diabetes 2020, 44, 44–52. [Google Scholar] [CrossRef]
- Surono, I.S.; Wardana, A.A.; Waspodo, P.; Saksono, B.; Verhoeven, J.; Venema, K. Effect of functional food ingredients on gut microbiota in a rodent diabetes model. Nutr. Metab. 2020, 17, 77. [Google Scholar] [CrossRef]
- Sane, F.; Scuotto, A.; Pierrat, V.; Kacet, N.; Hober, D.; Romond, M.B. Diabetes progression and alterations in gut bacterial translocation: Prevention by diet supplementation with human milk in NOD mice. J. Nutr. Biochem. 2018, 62, 108–122. [Google Scholar] [CrossRef]
- Shikano, A.; Kuda, T.; Shibayama, J.; Toyama, A.; Ishida, Y.; Takahashi, H.; Kimura, B. Effects of lactobacillus plantarum uruma-SU4 fermented green loofah on plasma lipid levels and gut microbiome of high-fat diet fed mice. Food Res. Int. 2019, 121, 817–824. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, R.K.; Malhotra, S.; Pothuraju, R.; Shandilya, U.K. Lactobacillus rhamnosus NCDC17 ameliorates type-2 diabetes by improving gut function, oxidative stress and inflammation in high-fat-diet fed and streptozotocin treated rats. Benef. Microbes 2017, 8, 243–255. [Google Scholar] [CrossRef]
- Cabello-Olmo, Μ.; Oneca, Μ.; Torre, P.; Sainz, N.; Moreno-Aliaga, M.J.; Guruceaga, E.; Díaz, J.V.; Encio, I.J.; Barajas, M.; Araña, M. A Fermented Food Product Containing Lactic Acid Bacteria Protects ZDF Rats from the Development of Type 2 Diabetes. Nutrients 2019, 11, 2530. [Google Scholar] [CrossRef]
- Hu, T.G.; Wen, P.; Shen, W.Z.; Liu, F.; Li, Q.; Li, E.N.; Liao, S.T.; Wu, H.; Zou, Y.X. Effect of 1-Deoxynojirimycin Isolated from Mulberry Leaves on Glucose Metabolism and Gut Microbiota in a Streptozotocin-Induced Diabetic Mouse Model. J. Nat. Prod. 2019, 82, 2189–2200. [Google Scholar] [CrossRef]
- Shah, N.J.; Swami, O.C. Role of probiotics in diabetes: A review of their rationale and efficacy. EMJ Diabet. 2017, 5, 104–110. [Google Scholar]
- Dolpady, J.; Sorini, C.; Di Pietro, C.; Cosorich, I.; Ferrarese, R.; Saita, D.; Clementi, M.; Canducci, F.; Falcone, M. Oral Probiotic VSL#3 Prevents Autoimmune Diabetes by Modulating Microbiota and Promoting Indoleamine 2,3-Dioxygenase-Enriched Tolerogenic Intestinal Environment. J. Diabetes Res. 2016, 2016, 7569431. [Google Scholar] [CrossRef]
- Morshedi, M.; Saghafi-Asl, M.; Hosseinifard, E.S. The potential therapeutic effects of the gut microbiome manipulation by symbiotic containing-Lactobacillus plantarum on neuropsychological performance of diabetic rats. Transl. Med. 2020. 18, 1–14. [CrossRef]
- Farida, E.; Nuraida, L.; Giriwono, P.E.; Jenie, B.L.E. Lactobacillus rhamnosus Reduces Blood Glucose Level through Downregulation of Gluconeogenesis Gene Expression in Streptozotocin-Induced Diabetic Rats. International Journal of Food Science 2020, 2020, 6108575. [Google Scholar] [CrossRef]
- Tian, P.; Li, B.; He, C.; Song, W.; Hou, A.; Tian, S.; Meng, X.; Lia, K.; Shan, Y. Antidiabetic (type 2) effects of Lactobacillus G15 and Q14 in rats through regulation of intestinal permeability and microbiota. Food Funct. 2016, 7, 3789–3797. [Google Scholar] [CrossRef]
- Li, K.K.; Tian, P.J.; Wang, S.D.; Lei, P.; Qu, L.; Huang, J.P.; Shan, Y.J.; Li, B.L. Targeting gut microbiota: Lactobacillus alleviated type 2 diabetes via inhibiting LPS secretion and activating GPR43 pathway. J. Funct. Foods 2017, 38, 561–570. [Google Scholar] [CrossRef]
- Li, X.; Wang, E.; Yin, B.; Fang, D.; Chen, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of Lactobacillus casei CCFM419 on insulin resistance and gut microbiota in type 2 diabetic mice. Benef 2017, 8, 421–432. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, X.; Guo, J.; He, Q.; Li, H.; Song, Y.; Zang, H. Lactobacillus casei reduces susceptibility to type 2 diabetes via microbiota-mediated body chloride ion influx. Sci. Rep. 2014, 4, 5654. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus casei CCFM419 attenuated type 2 diabetes via a gut microbiota dependent mechanism. Food Funct. 2017, 8, 3155–3164. [Google Scholar] [CrossRef]
- Allen, S.J.; Jordan, S.; Storey, M.; Thornton, C.A.; Gravenor, M.; Garaiova, I.; Plummer, S.F.; Wang, D.; Morgan, G. Dietary supplementation with lactobacilli and bifidobacteria is well tolerated and not associated with adverse events during late pregnancy and early infancy. J. Nutr. 2010, 140, 483–488. [Google Scholar] [CrossRef]
- Kassaian, N.; Feizi, A.; Aminorroaya, A.; Amini, M. Probiotic and synbiotic supplementation could improve metabolic syndrome in prediabetic adults: A randomized controlled trial. Diabetes Metab. Syndr. 2019, 13, 2991–2996. [Google Scholar] [CrossRef]
- Khalili, L.; Alipour, B.; Jafar-Abadi, M.A.; Faraji, I.; Hassanalilou, T.; Mesgari Abbasi, M.; Vaghef-Mehrabany, E.; Sani, A.M. The Effects of Lactobacillus casei on Glycemic Response, Serum Sirtuin1 and Fetuin-A Levels in Patients with Type 2 Diabetes Mellitus: A Randomized Controlled Trial. Iran Biomed. J. 2019, 23, 68–77. [Google Scholar] [CrossRef]
- Medina-Vera, I.; Sanchez-Tapia, M.; Noriega-López, L.; Granados-Portillo, O.; Guevara-Cruz, M.; Flores-López, A.; Avila-Nava, A.; Fernández, M.L.; Tovar, A.R.; Torres, N. A dietary intervention with functional foods reduces metabolic endotoxaemia and attenuates biochemical abnormalities by modifying faecal microbiota in people with type 2 diabetes. Diabetes Metab. 2019, 45, 122–131. [Google Scholar] [CrossRef]
- Sabico, S.; Al-Mashharawi, A.; Al-Daghri, N.M.; Wani, K.; Amer, O.E.; Hussain, D.S.; Ansari, A.M.G.; Masoud, M.S.; Alokail, M.S.; McTernan, P.G. Effects of a 6-month multi-strain probiotics supplementation in endotoxemic, inflammatory and cardiometabolic status of T2DM patients: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1561–1569. [Google Scholar] [CrossRef]
- Sato, J.; Kanazawa, A.; Azuma, K.; Ikeda, F.; Goto, H.; Komiya, K.; Kanno, R.; Tamura, Y.; Asahara, T.; Takahashi, T.; et al. Probiotic reduces bacterial translocation in type 2 diabetes mellitus: A randomised controlled study. Sci. Rep. 2017, 7, 12115. [Google Scholar] [CrossRef]
- Tonucci, L.B.; Dos Santos, O.K.M.; de Oliveira, L.L.; Ribeiro, R.S.M.; Martino, D.H.S. Clinical application of probiotics in type 2 diabetes mellitus: A randomized, double-blind, placebo-controlled study. Clin. Nutr. 2017, 36, 85–92. [Google Scholar] [CrossRef]
- Yanni, A.E.; Kartsioti, K.; Karathanos, V.T. The role of yogurt consumption in the management of type 2 diabetes. Food Funct. 2020, 11, 10306–10316. [Google Scholar]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef]


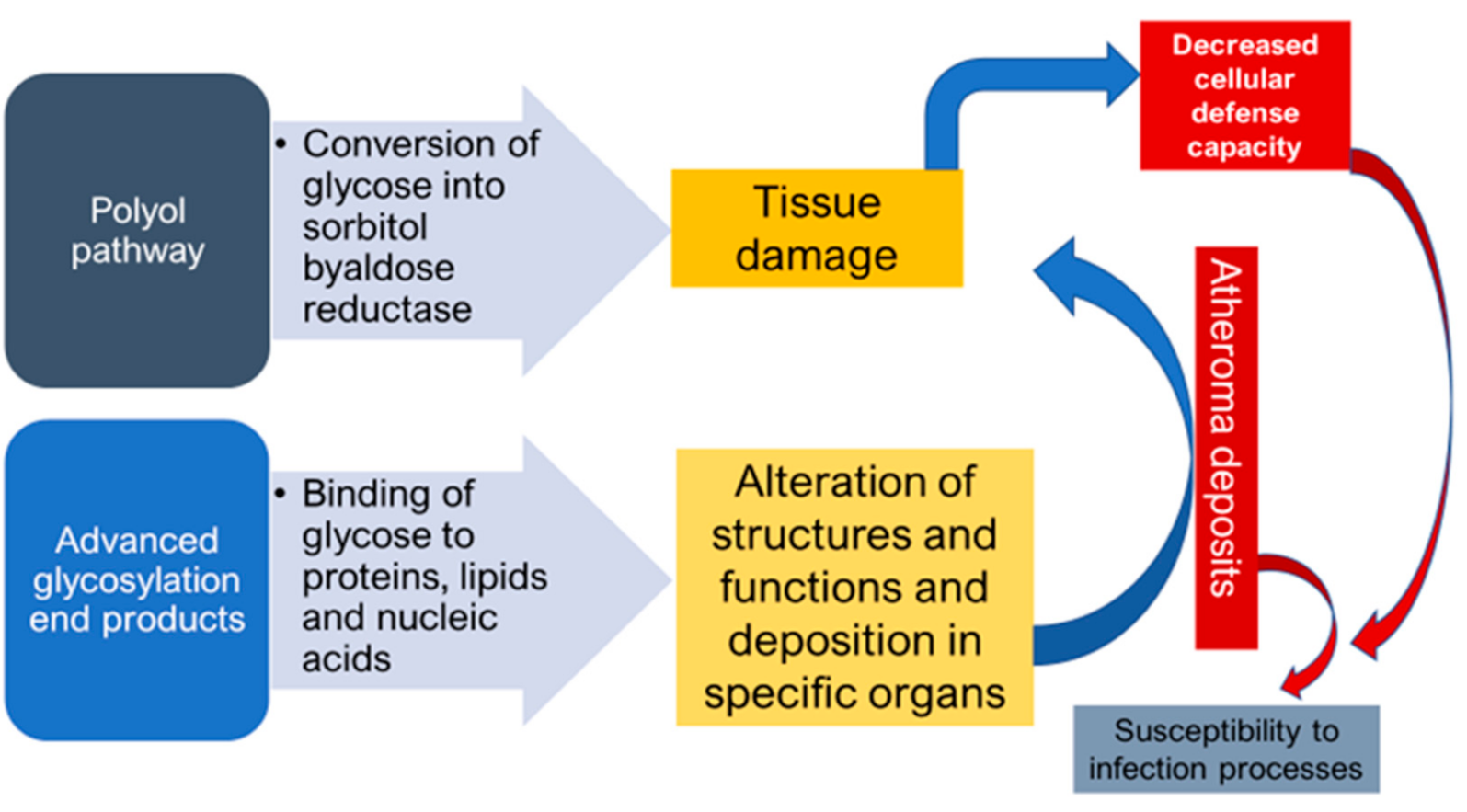{kind=link}
| Type of Compound | DietarySupplement | Animal Model | Outcomes In Immune System | Outcomes in Microbiota | Reference |
|---|---|---|---|---|---|
| Prebiotics | Propolis | STZ-induced diabetic Sprague-Dawley rats (T2DM), male | 240 mg/kg propolis led to lower FBG levels. Insulin resistance reduced after propolis treatment. Propolis treatment could repair the intestinal mucosal injury | Overall structure of the gut microbiota in diabetic rats was shifted toward that in normal rats. Lactobacillus genera were predominant in the control and propolis treatment groups. Significant down regulation of the abundances of Blautia, Fusicatenibacter and ClostridiumXlVa in the model group. | [115] |
| High fat diet group supplemented with pistachio nuts | C57BL/6J (B6), male | Decreased amounts of TNF-α and IL-1βin serum (HFD-P vs HFD). Inflammatory foci, IL-1β, CCL-2 gene expression were lower in the liver of HFD-P vs HFD. Improvement of inflammation in obese mice. | Lower Firmicutes/Bacteroidetes in HFD-P vs HFD/ Parabacteroides, Dorea, Allobaculum, Turicibacter, Lactobacillus and Anaeroplasma increased with pistachio. Oscillospira, Desulfovibrio, Coprobacillus and Biophila reduced with pistachio supplementation. | [116] | |
| Pistachio nuts | STZ-induced diabetic Wistar rats (T1DM), male | - | Elevated levels of lactobacilli and bifidobacteria in jejunum, ileum and cecum of diabetic animals. Increased fecal lactobacilli and bifidobacteria counts and decreased enterococci after 4 weeks of pistachio diet (in healthy and diabetic animals). | [117] | |
| Whole grain barley / barley malt | Wistar rats, male | - | Lower Firmicutes/ Bacteroidetes increased Actinobacteria and Verrucomicrobia, after barley supplementation. Akkermansia, Ruminococcus, Blautia, Biophila, Turicibacter, Roseburia higher after barley malt intake. | [118] | |
| Corn starch diet with chlorogenic acid | Wistar rats, male | Decreased inflammation and fat deposition in the liver along with reduced plasma liver enzyme activities of obese rats. | Increased diversity of gut microbiota. Rats showed decreased abundance of Actinobacteria and Bacteriodetes with increased abundance of Firmicutes. | [119] | |
| Taro starch, beet juice, L. plantarum IS-10506 | STZ-induced diabetic Sprague Dawley rats (T1DM), male | - | The RA of these six OTUs, Christensenellaceae R-7 group, Prevotella 9, an unknown species of the Prevotellaceae family, Prevotellaceae UCG-001 and Ruminococcaceae UCG-005 and Ruminococcus 1 were all increased after starch feeding. | [121] | |
| Probiotics | L. plantarum (probiotic), inulin (prebiotic) or in combination (symbiotic) | STZ-induced diabetic Wistar rats (T2DM), male | Improvement of oxidative stress status in hippocampus and prefrontal cortex. Neuropsychological improvement.Cognitive impairment is reversed after symbiotic administration. | Probiotic and synbiotic treatment led to increase of Firmicutes phylum and in Lactobacillales family, while Clostridiales and Bacteroidales were decreased. | [127] |
| VSL#3 containing Bifidobacteriaceae, Lactobacillaceae, and Streptococcus thermophilus, alone or in combination with Retinoic Acid. | Nod mice | Protection from diabetes progression Reduced degree of insulitis in histological examination. Inhibition of IL-1β expression. Enhancement of indoleamine-2,3- dioxygenase (IDO) and IL-33. | Increased Lactobacillacae, clostridia and Rikenellaceae after VSL#3 treatment. Decreased abundance of Bacteroidetes strain S24-7. | [128] | |
| L. rhamnosus BSL and L. rhamnosus R23 | STZ-induced diabetic Sprague-Dawley rats (T2DM), male | Fasting blood glucose (FBG) was significantly reduced after probiotic administration. Improved glucose tolerance and glucose control after probiotic administration. Decrease in Total Cholesterol (TC) | Increased LAB levels after 30 days of probiotic supplementation | [130] | |
| L. paracasei subsp. paracasei G15 and/or L. casei Q14 isolated from dairy food alone or in combination with metformin | STZ-induced diabetic Wistar rats in high-fat diet (T2DM), male | Restored glucose intolerance in all treatment groups after 6 weeks. FBG decreased after 13 weeks in all treatment groups. Probiotics reduced plasma LPS. The lactobacillus and metformin treatments significantly reduced both IL-1β and IL-8 levels. | Separated clustering of microbiota in each group. Microbiota of group supplemented with L. casei Q14 was located near the healthy group. | [131] | |
| L. paracasei subsp. paracasei G15 and/or L. casei Q14 isolated from dairy food | STZ-induced diabetic Wistar rats in high-fat diet (T2DM), male | Restored glucose tolerance and suppressed total cholesterol and triacylglycerol (TAG) level, after 6 weeks of probiotic administration. Hyperinsulinemia was ameliorated with insulin and glucagon levels being lower after probiotic ingestion. | Abundances of Lactobacillus and Bifidobacterium,Clostridiumleptum, Bacteroides, Prevotella, were increased after probiotic treatment. | [132] | |
| L.casei CCFM419 | STZ-induced diabetic C57BL/6J mice in high-fat diet (T2DM), male | Ingestion of L. casei CCFM419 led to lower FBG, reduced 2-h postprandial blood glucose. Improved insulin sensitivity by decreased fasting insulin concentration and HOMA-IR value. Decreased levels of TNF-α and IL-6 and IL-10. Reduced LDL-C and increased HDL-C levels. | The abundance of Allobaculum and Bacteriodes were increased after probiotic treatment. Decreased Firmicutes and increased Bacteroidetes. | [133] | |
| L. casei | STZ-induced diabetic Sprague-Dawley rats in high-fat sucrose diet (HFS) (T2DM), male | L. casei reduced the endotoxin LPS production induced by STZ. L. casei Zhang ingestion prevents from the onset and development of glycemia in both fasting and postprandial 2 h blood glucose levels and OGTT levels. Inhibition of pro inflammatory cytokines (IFN-c and TNF-α) after probiotic administration. | Caecal Bifidobacterium and Lactobacillus were more abundant in probiotic-treated rats in HFS diet than the plain HFS group. Higher C. coccoides–E.rectale group and C. scindens members in HFS than the probiotic HFS and control rats. | [134] | |
| L. rhamnosus CCFM0528 | STZ-induced diabetic C57BL/6J mice in high-fat diet (T2DM), male | Amelioration of insulin resistance, glucose tolerance, FBG and postprandial 2-h blood glucose. Decreased TNF-α and IL-6. Increased GLP-1. | Increased Bacteroidetes and decreased Firmicutes.Increased Bifidobacterium, Lactobacillus, Allobaculum and Bacteroides. | [135] | |
| Fermented products | Green loofah L. cylindrica homogenate (LH) and fermentedLH (FL) with Lactococcus lactis subsp. lactis Uruma-SU1 and L. plantarum Uruma-SU4, isolated from algal beach casts. | male Kwl: ddY mice in high-fat diet | TC, LDL-C, and the ratio LDL-C/HDL-C were lower after FL. | Caecal L. johnsonii and C. disporicum were increased through the consumption of fermented loofah. | [123] |
| Fermented milk by inoculation of skim milkwith probiotic cultures (L. rhamnosus NCDC 17 and L. rhamnosus GG) at 1% (v/v) | STZ-induced diabetic Wistar rats in high-fat diet (T2DM), male | L. rhamnosus NCDC 17 supplementation decreased expression levels of TNF-α and IL-6 genes, increased mRNA of the adiponectin gene. LGG and L. rhamnosus NCDC 17 decreased FGB levels. Lower insulin levels after probiotic treatment. | Both probiotic treatments increased the population of total bacteria. HFD + L. rhamnosus NCDC 17 group had higher populations of Eubacterium rectale-C. coccoides, Bacteroides, lactobacilli and bifidobacteria. | [124] | |
| Non dairy fermented food product | Zucker diabetic fatty (ZDF) rats, male | Decreased glucose absorption in treated group. Decreased blood glucose with the FFP. | Enriched microbial diversity after FFP administration. Increased Firmicutes, Saturella, Proteus, Alistipes, Anaerococcus in FFP group and Streptococcacae, Anaerococcus and Streptococcus, Barnesiella and Blautia in control group. | [125] | |
| Mixed fermentation by L. fermentum and Saccharomyces cerevisiae was used to enhance DNJ extraction efficiency from mulberry leaves | STZ-induced diabetic Kunming mice, male | - | Relieved gut dysbiosis in diabetic mice by promoting the growth of Lactobacillus, Lachnospiraceae NK4A136 group, Oscillibacter, Lachnospiraceae, Alistipes, and Bifidobacterium. | [126] | |
| Other products | Corn starch tea (instant or matcha) | Kunming mice, female | Particularly increased levels of Coriobacteriaceae, Lactobacillaceae, Prevotellaceae and Bifidobacteriaceae, and decreased Bacteroidaceae, Ruminococcaceae, Helicobacteraceae and Enterobacteriaceae. | [120] | |
| Human milk | Nod mice, female | Prevention of diabetes onset and progression. | Elevated fecal Bifidobacterium and Akkermansia by human milk. Cecal B. fragilis and E. coli lower after HM.Higher cecal and colonic B. vulgatus by human milk intake. | [122] |
| Fermented Food/Probiotics | Outcome in the Immune System | Reference |
|---|---|---|
| fermented milk containing L. acidophilus La-5 and B. animalis subsp lactis BB-12 | improved the glycemic control in T2DM subjects decrease in inflammatory cytokines (TNF-a and resistin) and increase of the acetic acid | [142] |
| The probiotic group consumed Lactobacillus casei strain Shirota-fermented milk | Probiotic administration reduced bacterial translocation and altered the gut microbiota in Japanese patients with T2DM | [141] |
| 6-months multi-strain probiotics supplementation in T2DM | beneficial role of probiotics in inflammation, promising adjuvant anti-diabetes therapy | [140] |
| reduced-energy diet with a dietary portfolio (DP) comprising high-fiber, polyphenol-rich and vegetable-protein functional foods | Increase in Faecalibacterium prausnitzii and Akkermansia muciniphila, two bacterial species known to have anti-inflammatory effects. Dietary intervention with functional foods significantly modified fecal microbiota by increasing alpha diversity. | [139] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezirtzoglou, E.; Stavropoulou, E.; Kantartzi, K.; Tsigalou, C.; Voidarou, C.; Mitropoulou, G.; Prapa, I.; Santarmaki, V.; Kompoura, V.; Yanni, A.E.; et al. Maintaining Digestive Health in Diabetes: The Role of the Gut Microbiome and the Challenge of Functional Foods. Microorganisms 2021, 9, 516. https://doi.org/10.3390/microorganisms9030516
Bezirtzoglou E, Stavropoulou E, Kantartzi K, Tsigalou C, Voidarou C, Mitropoulou G, Prapa I, Santarmaki V, Kompoura V, Yanni AE, et al. Maintaining Digestive Health in Diabetes: The Role of the Gut Microbiome and the Challenge of Functional Foods. Microorganisms. 2021; 9(3):516. https://doi.org/10.3390/microorganisms9030516
Chicago/Turabian StyleBezirtzoglou, Eugenia, Elisavet Stavropoulou, Konstantina Kantartzi, Christina Tsigalou, Chrysa Voidarou, Gregoria Mitropoulou, Ioanna Prapa, Valentini Santarmaki, Vasiliki Kompoura, Amalia E. Yanni, and et al. 2021. "Maintaining Digestive Health in Diabetes: The Role of the Gut Microbiome and the Challenge of Functional Foods" Microorganisms 9, no. 3: 516. https://doi.org/10.3390/microorganisms9030516
APA StyleBezirtzoglou, E., Stavropoulou, E., Kantartzi, K., Tsigalou, C., Voidarou, C., Mitropoulou, G., Prapa, I., Santarmaki, V., Kompoura, V., Yanni, A. E., Antoniadou, M., Varzakas, T., & Kourkoutas, Y. (2021). Maintaining Digestive Health in Diabetes: The Role of the Gut Microbiome and the Challenge of Functional Foods. Microorganisms, 9(3), 516. https://doi.org/10.3390/microorganisms9030516