
Agroecological Service Crops Drive Plant Mycorrhization in Organic Horticultural Systems

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiments Setup and Treatments
2.1.1. Case-Study on Organic Melon
2.1.2. Case-Study on Young Organic Citrus Trees
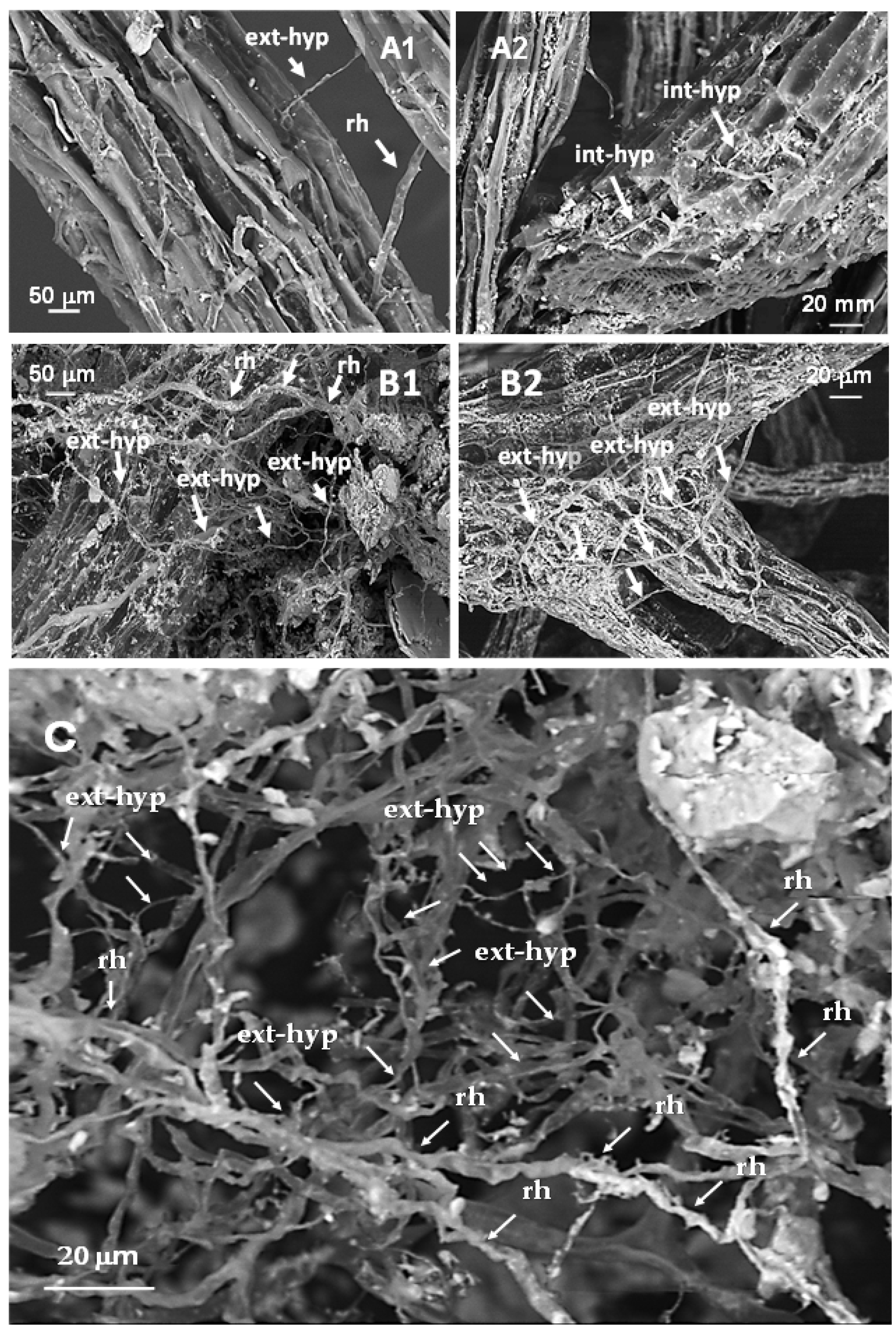
2.2. Determinations
- in the organic melon experiment, by sampling at melon ripening at the 47th day (June 2014) and at melon harvesting on the 56th (August 2014) day after transplanting (DAT), respectively. Total and by-species weed density (pp m−2) and cover (%, m2 /m2) were recorded by placing three randomly selected 0.25 × 0.25 m2 (June) and 1.0 × 1.0 m2 quadrats (August) within each plot.
- in the orange young trees experiment by sampling at barley boot (on 3rd February) and termination (on 16th April 2014), respectively. The total and by-species weed density were recorded by placing twelve randomly selected 1.0 × 1.0 m2 quadrats within each plot.
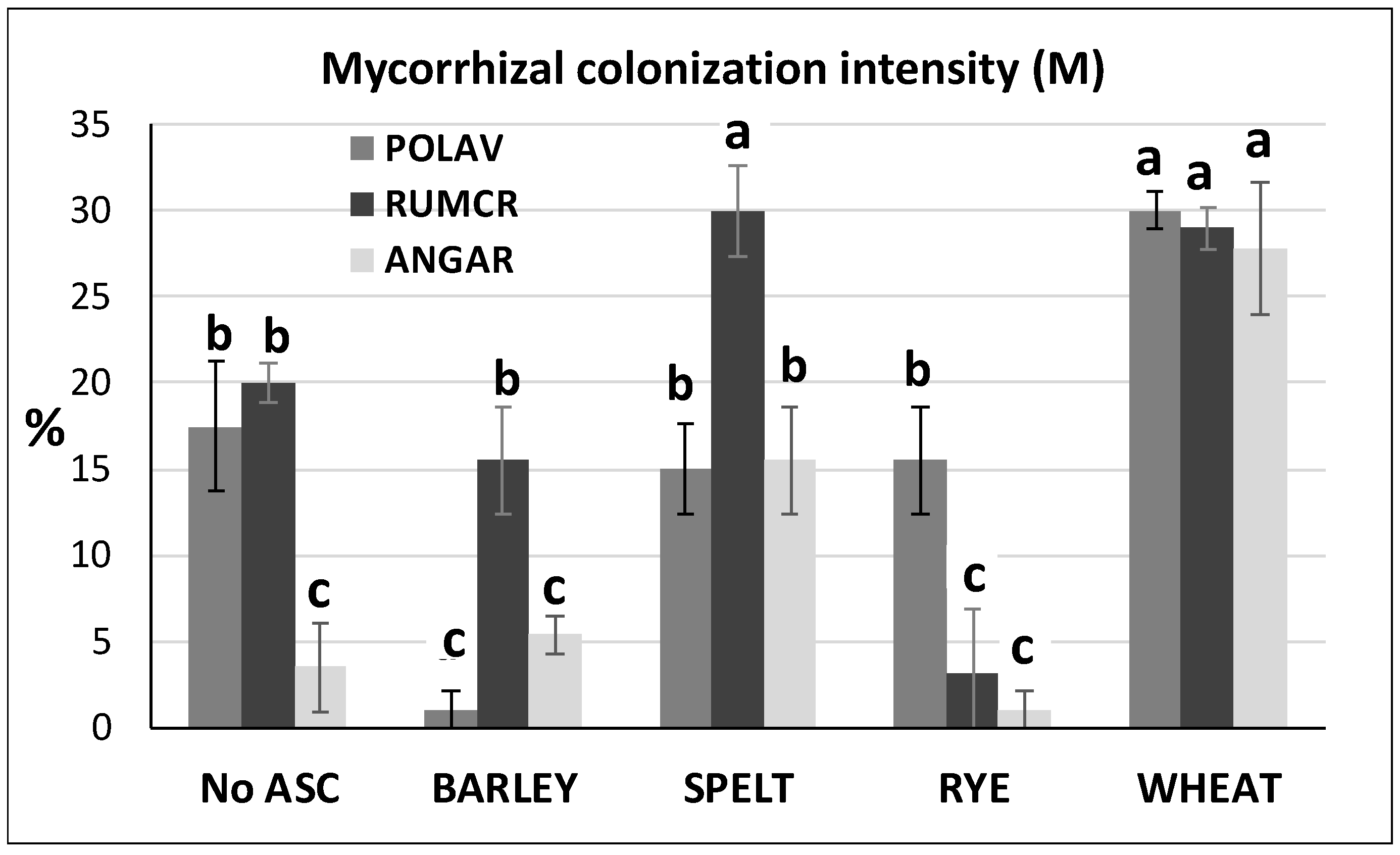
- in the organic melon system, for each treatment and each field replicate, root mycorrhizal colonization intensity (M%) was determined on melon and on three selected and most representative SAM weed species, Rumex crispus L. (RUMCR), Polygonum aviculare L. (POLAV) and Anagallis arvensis L. (ANGAR). The root apparatus of melon and weeds was sampled from the field by using stainless steel cylinders of 6 cm diameter and 20 cm length [10]. For each species, three root subsamples per plot were collected, then pooled to obtain n.1 root sample × n. 3 treatments × n. 3 blocks.
- in young organic orange trees, for each treatment and each field replicate, root mycorrhizal colonization intensity (M%) was determined on orange tree roots, by using stainless steel cylinders of 9-cm diameter and 30-cm length, to obtain n. 2 root samples × n. 2 treatments × n. 3 blocks.
- in the organic melon case-study, at harvest (13th August 2014), melon yield (Mg ha−1), fruit average weight (kg) and sugar content (Brix) were determined to evaluate the cash crop production and quality.
- in the organic orange trees case-study, after one month from ASC termination (12th May), the Soil Plant Analysis Development (SPAD) value, the foliar P content (g kg−1), the plant height (cm), the canopy diameter (cm) and the canopy volume (m3) were determined on all sampled trees.
2.3. Statistical Analysis
3. Results
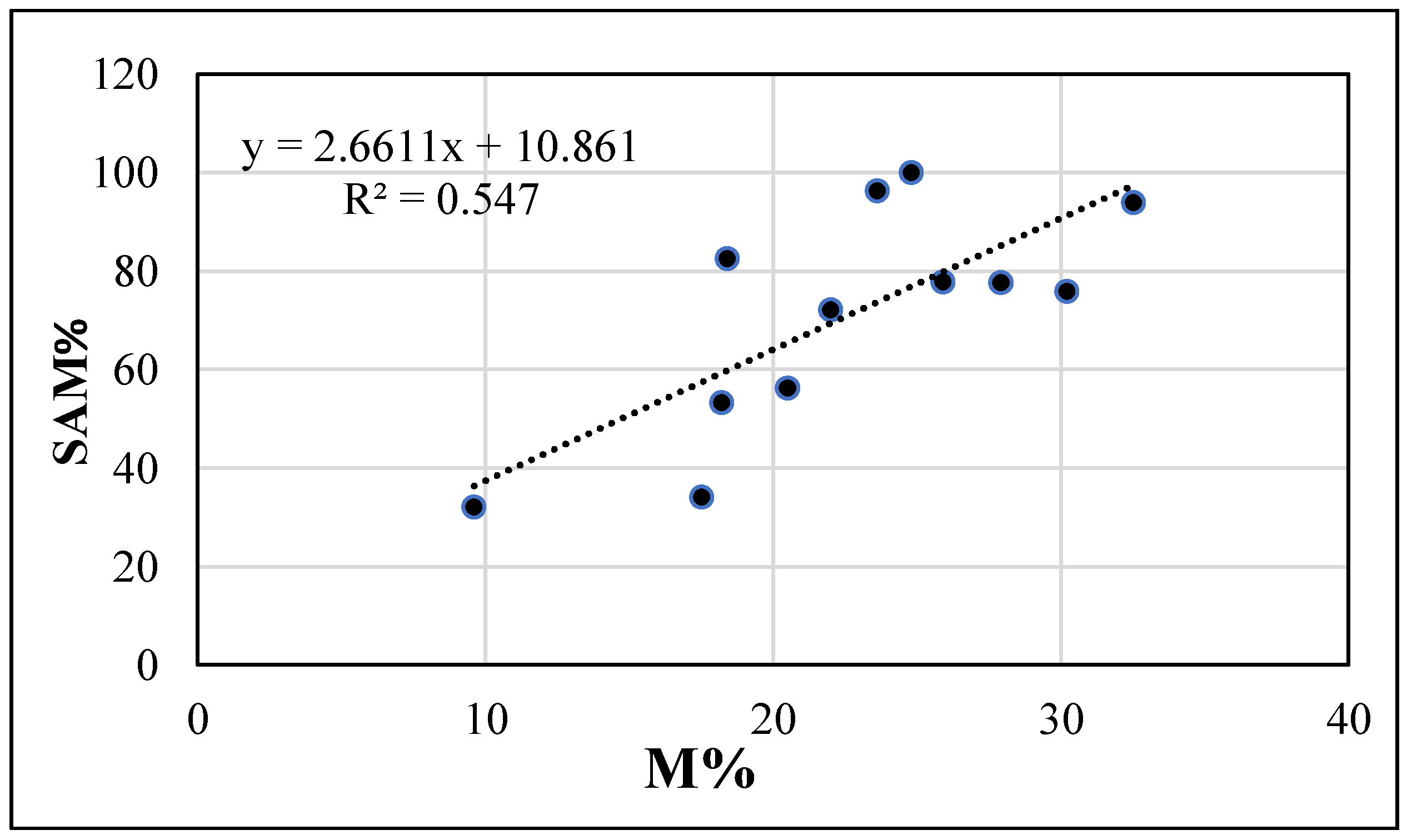
3.1. Case-Study on Organic Melon
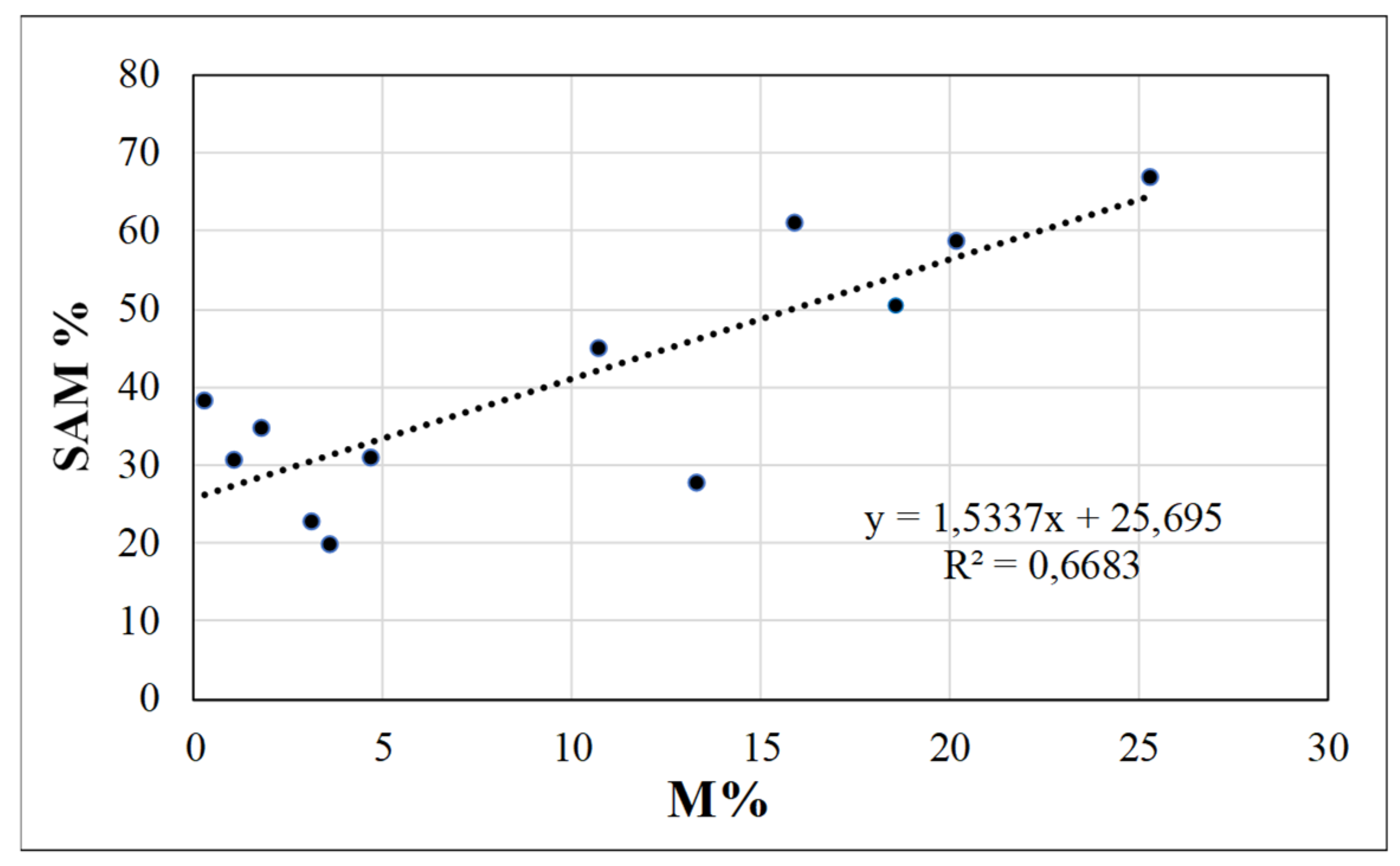
3.2. Case-Study on Young Organic Citrus Trees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flach, K.W. Low-input agriculture and soil conservation. J. Soil Water Cons. 1990, 45, 42–44, ISSN: 1941-3300.. [Google Scholar]
- Altieri, M.A. Organic farming. In Agroecology: The Science of Sustainable Agriculture, 2nd ed.; Altieri, M.A., Ed.; CRC Press–Taylor & Francis Group: London, UK, 1995; Part 2; pp. 179–204. ISBN 978-0-8133-1718-2. [Google Scholar]
- Raviv, M. Sustainability of organic farming. Hortic. Rev. 2010, 36, 289–333. [Google Scholar]
- Diacono, M.; Persiani, A.; Testani, E.; Montemurro, F.; Ciaccia, C. Recycling agricultural wastes and by-products in organic farming: Biofertilizer production, yield performance and carbon footprint analysis. Sustainability 2019, 11, 3824. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Alternative Pathways to 2050. Rome. 224 pp. Licence: CC BY-NC-SA 3.0 IGO. ISBN 978-92-5-130158-6. 2018. Available online: http://www.fao.org/publications/fofa/en/ (accessed on 23 November 2020).
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: London, UK, 1997; ISBN 9780080559346. [Google Scholar]
- Van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Hill, J.O.; Simpson, R.J.; Ryan, M.H.; Chapman, D.F. Root hair morphology and mycorrhizal colonisation of pasture species in response to phosphorus and nitrogen nutrition. Crop. Past Sci. 2010, 61, 122–131. [Google Scholar] [CrossRef]
- Plaxton, W.; Lambers, H. Phosphorus Metabolism in Plants; Annual Plant Reviews; Wiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 379–387. ISBN 9781118958834. [Google Scholar]
- Trinchera, A.; Testani, E.; Ciaccia, C.Q.; Campanelli, G.; Leteo, F.; Canali, S. Effects induced by living mulch on rhizosphere interactions in organic artichoke: The cultivar’s adaptive strategy. Renew. Agr. Food Syst. 2016, 32, 214–223. [Google Scholar] [CrossRef]
- Miceli, A.; Romano, C.; Moncada, A.; Piazza, G.; Torta, L.; D’Anna, F.; Vetrano, F. Yield and quality of mini-watermelon as affected by grafting and mycorrhizal inoculum. J. Agr. Sci. Tech. 2016, 18, 505–516. [Google Scholar]
- Salvioli, A.; Novero, M.; Lacourt, I.; Bonfante, P. The Impact of Mycorrhizal Symbiosis on Tomato Fruit Quality. In Proceedings of the 16th IFOAM Organic World Congress, Modena, Italy, 16–20 June 2008; Available online: http://orgprints.org/view/projects/conference.html (accessed on 21 December 2020).
- Simard, S.W.; Durall, D.M. Mycorrhizal networks: A review of their extent, function, and importance. Can. J. Bot. 2004, 82, 1140–1165. [Google Scholar] [CrossRef]
- Trinchera, A.; Ciaccia, C.; Testani, E.; Baratella, V.; Campanelli, C.; Leteo, F.; Canali, S. Mycorrhiza-mediated interference between cover crop and weed in organic winter cereal agroecosystems: The mycorrhizal colonization intensity indicator. Ecol. Evol. 2019, 9, 5593–5604. [Google Scholar] [CrossRef] [PubMed]
- Essel, E.; Junhong, X.; Chaochao, D.; Zhengkai, P.; Jinbin, W.W.; Jicheng, S.; Jianhui, X.; Jeffrey, A.C.; Lingling, L. Bacterial and fungal diversity in rhizosphere and bulk soil under different long-term tillage and cereal/legume rotation. Soil Till. Res. 2019, 194, 104302. [Google Scholar] [CrossRef]
- Víllora, R.A.; Plaza, E.H.; Navarrete, L.; Sánchez, M.J.; Sánchez, A.M. Climate and tillage system drive weed communities’ functional diversity in a Mediterranean cereal-legume rotation. Agr. Ecosyst. Environ. 2019, 283, 106574. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Ineichen, K.; Ris, E.H.; Boller, T.; Wiemken, A. Community structure of arbuscular mycorrhizal fungi at different soil depthsin extensively and intensively managed agroecosystems. New Phyt. 2005, 165, 273–283. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; de Ruiter, P.C.; van der Putten, W.H.; Birkhofer, K.; Hemerik, L.; de Vries, F.T.; Bardgett, R.D.; Brady, M.V.; et al. Intensive agriculture reduces soil biodiversity across Europe. Glob. Chan. Biol. 2015, 21, 973–985. [Google Scholar] [CrossRef]
- Canali, S.; Diacono, M.; Campanelli, G.; Montemurro, F. Organic no-till with roller crimpers: Agro-ecosystem services and Applications in organic Mediterranean vegetable productions. Sustain. Agric. Res. 2015, 4, 70–79. [Google Scholar] [CrossRef]
- Ciaccia, C.; Testani, E.; Campanelli, G.; Sestili, S.; Leteo, F.; Tittarelli, F.; Riva, F.; Canali, S.; Trinchera, A. Ecological service providing crops effect on melon-weed competition and allelopathic interactions. Org. Agric. 2015, 5, 199–207. [Google Scholar] [CrossRef]
- Campanelli, G.; Testani, E.; Canali, S.; Ciaccia, C.; Leteo, F.; Trinchera, A. Effects of cereals as agroecological service crops and no-till on organic melon, weeds and N. dynamics. Biol. Agric. Hort. 2019, 35, 275–287. [Google Scholar] [CrossRef]
- Navarro-Miró, D.; Blanco-Moreno, J.M.; Ciaccia, C.; Chamorro, L.; Testani, E.; Kristensen, H.L.; Hefner, M.; Tamm, K.; Bender, I.; Jakop, M.; et al. Agroecological service crops managed with roller crimper reduce weed density and weed species richness in organic vegetable systems across Europe. Agron. Sust. Dev. 2019, 39, 55. [Google Scholar] [CrossRef]
- Trinchera, A.; Testani, E.; Ciaccia, C.; Campanelli, G.; Leteo, F.; Canali, S. Rhizosphere interactions in an organic horticultural cropping system: Effect of living mulch on artichoke mycorrhization. Acta Hortic. 2016, 1137, 11–18. [Google Scholar] [CrossRef]
- Trinchera, A.; Testani, E.; Ciaccia, C.; Tittarelli, F.; Canali, S. May barley and rye extracts have an allelopathic inhibition effect on weed seedling root development by suppressing mycorrhization? Acta Hortic. 2015, 1105, 361–368. [Google Scholar] [CrossRef]
- Ortas, I. Role of mycorrhizae on mineral nutrition of fruit trees. Acta Hortic. 2018, 1217, 271–284. [Google Scholar] [CrossRef]
- Cruz, A.F.; de Carvalho Pires, M.; Beda do Nascimento, L.K.; Gerosa, R.M.L.; Oliveira, S.A.; Blum, B.L.E.; Yamanishi, O.K. Cover cropping system and mulching can shape soil microbial status in fruit orchards. Sci. Agric. 2020, 77, e20180316. [Google Scholar] [CrossRef]
- Trinchera, A.; Torrisi, B.; Allegra, M.; Rinaldi, S.; Rea, E.; Intrigliolo, F.; Roccuzzo, G. Effects of organic fertilization on soil organic matter and root morphology and density of orange trees. Acta Hortic. 2015, 1065, 1807–1814. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K.; Ying-Ning, Z.; Malhotra, S.K. Mycorrhizas in citrus: Beyond soil fertility and plant nutrition. Ind. J. Agric. Sci. 2017, 87, 423–427. [Google Scholar]
- Njeru, E.; Avio, L.; Sbrana, C.; Turrini, A.; Bocci, G.; Barberi, P.; Giovannetti, M. First evidence for a major cover crop effect on arbuscular mycorrhizal fungi and organic maize growth. Agron. Sust. Dev. 2014, 34, 841–848. [Google Scholar] [CrossRef]
- Moora, M. Mycorrhizal traits and plant communities: Perspectives for integration. J. Veg. Sci. 2014, 25, 1126–1132. [Google Scholar] [CrossRef]
- UNESCO-FAO. Bioclimatic Map of the Mediterranean Zone: Ecological Study of the Mediterranean Zone, Explanatory Notes; UNESCO-FAO: Paris, France, 1963; Volume 21, Available online: https://www.worldcat.org/title/bioclimatic-map-of-the-mediterranean-zone-ecological-study-of-the-mediterranean-zone-explanatory-notes/oclc/974951 (accessed on 29 November 2020).
- U.S. Department of Agriculture. Soil Survey Laboratory Methods Manual. In Natural Resource Conservation Service. Soil Survey Investigations Report; N 42, vers. 3.0. Washington DC, USA; 1996. Available online: www.section508.gov (accessed on 21 December 2020).
- IUSS Working Group. WRB World Reference Base for Soil Resources 2014; International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; Available online: http://www.fao.org/3/i3794en/I3794en.pdf (accessed on 21 December 2020).
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. Discuss. 2007, 4, 439–473. [Google Scholar]
- Canali, S.; Trinchera, A.; Intrigliolo, F.; Pompili, L.; Nisini, L.; Mocali, S.; Torrisi, B. Effect of long term addition of composts and poultry manure on soil quality of citrus orchards in Southern Italy. Biol. Fertil. Soils 2004, 40, 206–210. [Google Scholar] [CrossRef]
- Bàrberi, P.; Bocci, G.; Carlesi, S.; Armengot, L.; Blanco-Moreno, J.M.; Sans, F.X. Linking species traits to agroecosystem services: A functional analysis of weed communities. Weed Res. 2018, 58, 76–88. [Google Scholar] [CrossRef]
- Grace, C.; Stribely, D.P. A safer procedure for routine staining of vesicular-arbuscular mycorrhizal fungi. Mycolog. Res. 1991, 95, 1160–1162. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kouch, J.; Gianinazzi-Pearson, V. Mesure du taux de colonization on VA d’un systeme radiculaire: Colonizat of method d’estimation ayantune signification fonctionelle. In Ier Seminaire Dijon; Les Mycorhizes: Phisiologie and Genetique; INRA: Paris, France, 1986; pp. 217–221. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2017. R Package Version 1.0.5. Available online: https://CRAN.Rproject.org/package=factoextra (accessed on 2 September 2020).
- Fitter, A.H.; Peat, H.J. The ecological flora database. J. Ecol. 1994, 82, 415–425. [Google Scholar] [CrossRef]
- Barni, E.; Siniscalco, C. Vegetation dynamics and arbuscular mycorrhiza in old-field successions of the western Italian Alps. Mycorrhiza 2000, 10, 63–72. [Google Scholar] [CrossRef]
- Dickie, I.A.; Martinez-Garcia, L.B.; Koele, N.; Grelet, G.A.; Tylianakis, J.M.; Peltzer, D.A.; Richardson, S.J. Mycorrhizas and mycorrhizal fungal communities throughout ecosystem development. Plant Soil 2013, 367, 11–39. [Google Scholar] [CrossRef]
- Ciaccia, C.; Canali, S.; Campanelli, G.; Testani, E.; Montemurro, F.; Leteo, F.; Delate, K. Effect of roller-crimper technology on weed management in organic zucchini production in a Mediterranean climate zone. Renew. Agric. Food Syst. 2016, 31, 111–121. [Google Scholar] [CrossRef]
- Ciaccia, C.; Armengot Martinez, L.; Testani, E.; Leteo, F.; Campanelli, G.; Trinchera, A. Weed functional diversity as affected by agroecological service crops and no-till in a mediterranean Organic vegetable system. Plants 2020, 9, 689. [Google Scholar] [CrossRef]
- Derkowska, E.; Sas-Paszt, L.; Sumorok, B.; Szwonek, E.; Sawomir, G. The influence of mycorrhization and organic mulches on mycorrhizal frequency in apple and strawberry roots. J. Fruit Ornam. Plant Res. 2008, 16, 227–242. [Google Scholar]
- Bertholdsson, N.O. Variation in allelopathic activity over 100 years of barley selection and breeding. Weed Res. 2004, 44, 78–86. [Google Scholar] [CrossRef]
- Lebecque, S.; Crowet, J.; Lins, L.; Delory, B.M.; du Jardin, P.; Fauconnier, M.L.; Deleu, M. Interaction between the barley allelochemical compounds gramine and hordenine and artificial lipid bilayers mimicking the plant plasma membrane. Sci. Rep. 2018, 8, 9784. [Google Scholar] [CrossRef]
- Testani, E.; Ciaccia, C.; Campanelli, G.; Leteo, F.; Salvati, L.; Canali, S. Mulch-based no-tillage effects on weed community and management in an organic vegetable system. Agronomy 2019, 9, 594. [Google Scholar] [CrossRef]
- Armengot, L.; Blanco-Moreno, J.M.; Bàrberi, P.; Bocci, G.; Carlesi, S.; Aendekerk, R.; Bernerb, A.; Celettee, F.; Grossef, M.; Huitingg, H.; et al. Tillage as a driver of change in weed communities: A functional perspective. Agric. Ecosyst. Environ. 2016, 222, 276–285. [Google Scholar] [CrossRef]
- Canali, S.; Campanelli, G.; Ciaccia, C.; Leteo, F.; Testani, E.; Montemurro, F. Conservation tillage strategy based on the roller crimper technology for weed control in Mediterranean vegetable organic cropping systems. Eur. J. Agron. 2013, 50, 11–18. [Google Scholar] [CrossRef]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crop. Res. 2015, 183, 56–68. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Kirnak, H.; Tas, I. Mycorrhizal colonisation improves fruit yield and water use efficiency in watermelon (Citrullus lanatus Thunb.) grown under well-watered and water-stressed conditions. Plant Soil 2003, 253, 287–292. [Google Scholar] [CrossRef]
- Douds, D.D., Jr.; Nagahashi, G.; Reider, C.; Hepperly, P.R. Inoculation with arbuscular mycorrhizal fungi increases the yield of potatoes in a high P soil. Biolog. Agric. Hortic. 2007, 25, 67–78. [Google Scholar] [CrossRef]
- Conversa, G.; Lazzizera, C.; Bonasia, A.; Elia, A. Yield and phosphorus uptake of a processing tomato crop grown at different phosphorus levels in a calcareous soil as affected by mycorrhizal inoculation under field conditions. Biol. Fertil. Soil 2012, 49, 1–13. [Google Scholar] [CrossRef]
- Bowman, G.; Shirley, C.; Cramer, C. Managing Cover Crops Profitably, 2nd ed.; Sustainable Agriculture Publications: Burlington, VT, USA, 1998; p. 212. [Google Scholar]
- Liu, L.; Gitz, D.C., III.; McClure, J.W. Effects of UV-B on flavonoids, ferulic acid, growth and photosynthesis in barley primary leaves. Physiol. Plantarum 1995, 93, 725–733. [Google Scholar] [CrossRef]
- Dhima, K.V.; Vasilakoglou, I.B.; Eleftherohorinos, I.G.; Lithourgidis, A.S. Allelopathic potential of vwinter cereals and their cover crop mulch effect on grass weed suppression and corn development. Crop. Sci. 2006, 46, 345–352. [Google Scholar] [CrossRef]
- Ben-Hammouda, M.; Ghorbal, H.; Kremer, R.J.; Oueslati, O. Allelopathic effects of barley extracts on germination and seedlings growth of bread and durum wheats. Agronomie 2001, 21, 65–71. [Google Scholar] [CrossRef]
- Hamido, S.A.; Morgan, K.T. Effect of various irrigation rates on growth and root development of young citrus trees in high-density planting. Plants 2020, 9, 1462. [Google Scholar] [CrossRef] [PubMed]
- Vatovec, C.; Nicholas, J.; Huerd, S.C. Responsiveness of certain agronomic weed species to arbuscular mycorrhizal fungi. Renew. Agric. Food Sys. 2005, 20, 181–189. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
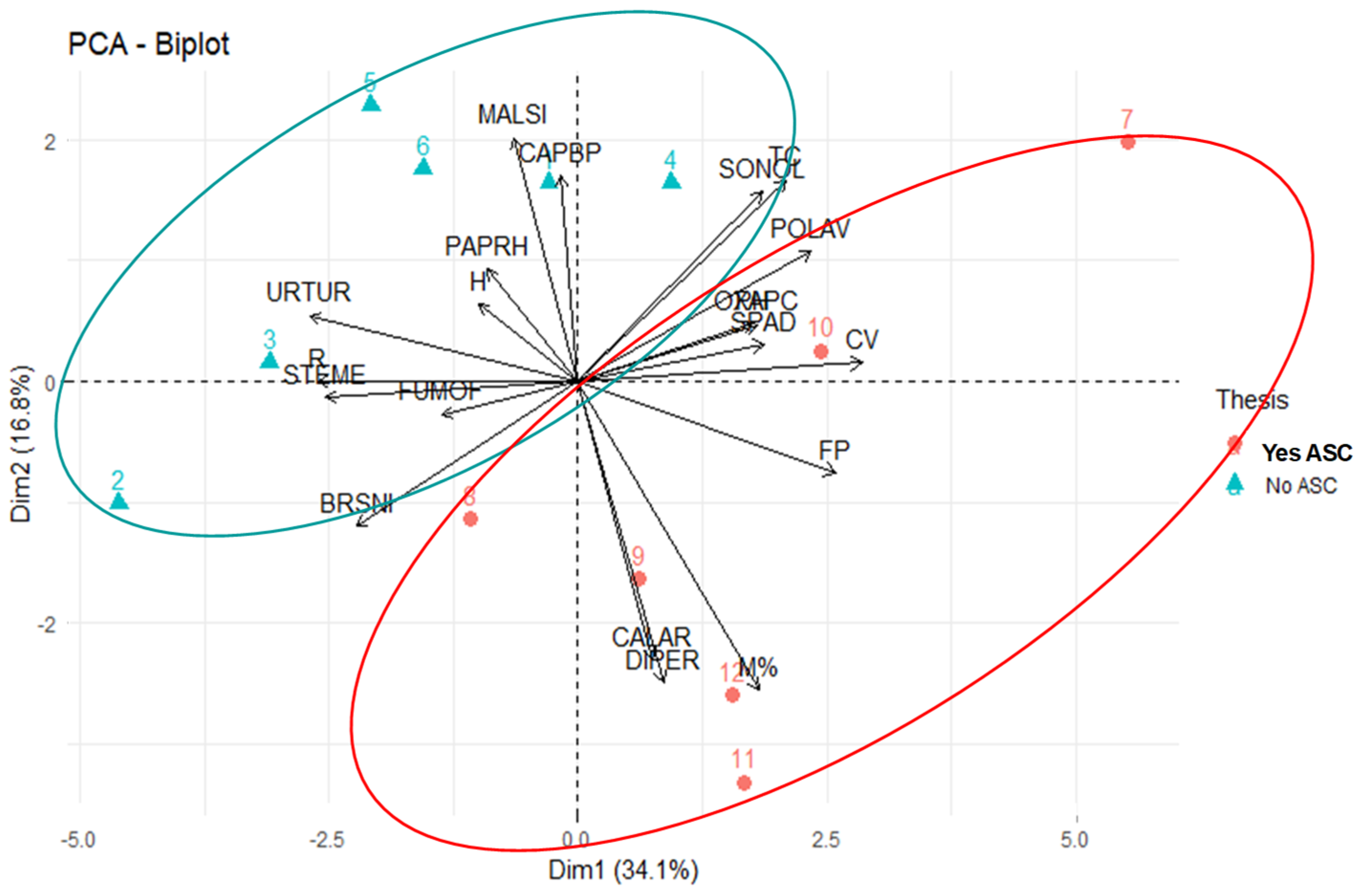{kind=link}
| Species | EPPO 1 code | Raunkiaer 2 BG | Habit | N 12 | SAM | Vegetable System | Orange System | Reference | ||
|---|---|---|---|---|---|---|---|---|---|---|
| June | Aug. | Feb. | Apr. | |||||||
| Aster squamatus | ASTSQ | T 3 Scap 7 | Erect | 7 | * | * | ||||
| Amaranthus retroflexus | AMARE | T Scap | Erect | 9 | * | * | * | * | ||
| Anagallis arvensis | ANGAR | T Rept 8 | Prostrate | 6 | X | * | * | * | * | A |
| Beta vulgaris | BETVU | H 4 Scap | Rosette | 5 | * | * | ||||
| Bromus sterilis | BROST | T Scap | Erect | 5 | X | * | * | A | ||
| Brassica nigra | BRSNI | T Scap | Erect | 4 | * | * | ||||
| Calendula arvensis | CALAR | T Scap | Erect | 5 | X | * | * | A | ||
| Capsella bursa-pastoris | CAPBP | H Bienne 8 | Rosette | 4 | * | * | ||||
| Chondrilla juncea | CHOJU | H 4 Scap | Erect | x | X | * | * | A | ||
| Convolvolus arvensis | CONAR | G 5 Rhiz 9 | Prostrate/creeping | 5 | X | * | * | * | * | A |
| Conyza canadensis | ERICA | T Scap | Erect | 7 | X | * | * | A | ||
| Diplotaxis erucoides | DIPER | T Scap | Erect | 5 | * | * | ||||
| Echinochloa crus-galli | ECHCG | T Scap | Erect | 8 | * | * | ||||
| Fumaria officinalis | FUMOF | T Scap | Erect | 6 | * | * | ||||
| Glebionis segetum | CHYSE | T Scap | Erect | 5 | X | * | * | A | ||
| Helminthotheca echioides | PICEC | T Scap | Erect | 2 | X | * | * | A | ||
| Lamium amplexicaule | LAMAM | T Scap | Erect | 7 | * | * | ||||
| Lolium perenne | LOLPE | H Caesp 10 | Erect | 7 | X | * | * | A | ||
| Malva silvestris | MALSI | H Scap | Erect | 8 | * | * | ||||
| Mercurialis annua | MERAN | T Scap | Erect | 8 | X | * | * | A | ||
| Oxalis pes-caprae | OXAPC | G Bulb 10 | Prostrate | 5 | * | * | ||||
| Papaver rhoeas | PAPRH | T Scap | Erect | x | X | * | * | A | ||
| Plantago media | PLAME | H Ros 11 | Rosulate | 3 | X | * | * | A | ||
| Poa annua | POAAN | T Caesp | Erect | 8 | X | * | * | A | ||
| Polygonum aviculare | POLAV | T Rept | Prostrate/creeping | 1 | X | * | * | * | * | B |
| Portulaca oleracea | POROL | T Scap | Prostrate/creeping | 7 | X | * | * | A | ||
| Raphanus raphanistrum | RAPRA | T Scap | Erect | 5 | * | * | ||||
| Rumex crispus | RUMCR | H Scap | Erect | 5 | X | * | * | B | ||
| Senecio vulgare | SENVU | T Scap | Erect | 8 | X | * | * | A | ||
| Setaria viridis | SETVI | T Scap | Erect | 7 | X | * | * | A | ||
| Sonchus oleraceus | SONOL | T Scap | Erect | 6 | X | * | * | * | * | A |
| Stellaria media | STEME | T Rept | Prostrate | 8 | X | * | * | * | * | B |
| Trifolium repens | TRFRP | Ch 6 Rept | Prostrate | 7 | X | * | * | C | ||
| Urtica urens | URTUR | T Scap | Erect | 8 | * | * | ||||
| Verbascum sinuatum | VESSI | H Bienne | Rosette | 7 | * | * | ||||
| Veronica persica | VERPE | T Scap | Prostrate | 6 | X | * | * | * | * | A |
| Melon | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| June 2014 (Melon Ripening) | August 2014 (Melon Harvesting) | |||||||||||
| SAM (%) | Density (pp m−2) | R | H | SAM (%) | Cover (%) | R | H | M% | Yield (Mg ha−1) | Fruit Weight (kg fruit−1) | Sugar Content (Brix) | |
| ASC | ||||||||||||
| No | 66.3 | 345 a | 4.3 | 0.90 b | 40.8 b | 87.5 | 6.3 b | 1.15 b | 15.9 b | 10.5 b | 1.5 | 7.4 b |
| Yes | 44.4 | 196 b | 5.7 | 1.36 a | 83.6 a | 79.1 | 9.5 a | 1.65 a | 24.5 a | 12.2 a | 1.2 | 8.0 a |
| Sig. | n.s. | ** | n.s. | * | *** | n.s. | * | * | ** | * | n.s. | * |
| ASC spec. | ||||||||||||
| Barley | 42.7 | 238 ab | 6.3 | 1.33 | 73.6 | 79.2 | 10.0 ab | 1,63 | 17.6 b | 8.27 | 0.97 | 8.07 |
| Rye | 49.6 | 139 b | 5.3 | 1.63 | 91.8 | 79.2 | 8.3 b | 1,77 | 27.0 ab | 13.76 | 1.17 | 7.95 |
| Spelt | 46.8 | 116 b | 4.7 | 1.23 | 84.6 | 70.8 | 8.0 b | 1,44 | 20.7 b | 16.22 | 1.49 | 7.75 |
| Wheat | 38.6 | 292 a | 6.3 | 1.37 | 84.4 | 87.5 | 11.7 a | 1,76 | 32.8 a | 10.69 | 1.32 | 8.45 |
| Sig. | n.s. | ** | n.s. | n.s. | n.s. | n.s. | * | n.s. | * | n.s. | n.s. | n.s. |
| Young Orange Trees | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| February 2014 (DAS 135) | April 2014 (DAS 187) | |||||||||||||
| SAM(%) | Density (pp m−2) | R | e | SAM (%) | Density (pp m−2) | R | H | M% | SPAD | Foliar P (g kg−1) | Plant Height (cm) | Canopy Diameter (cm) | Canopy Volume (m3) | |
| ASC | ||||||||||||||
| No | 40.8 a | 396 a | 5.3 b | 0.77 a | 35.4 | 192 a | 3.6 b | 1.8 | 2.4 b | 74.37 | 1.50 b | 157 | 88 | 0.65 b |
| Yes | 18.7 b | 238 b | 7.7 a | 0.64 b | 43.7 | 41 b | 6.5 a | 1.5 | 17.2 a | 75.59 | 1.71 a | 164 | 95 | 0.83 a |
| Sig. | *** | *** | ** | *** | n.s. | *** | * | n.s. | *** | n.s. | ** | n.s. | n.s. | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trinchera, A.; Testani, E.; Roccuzzo, G.; Campanelli, G.; Ciaccia, C. Agroecological Service Crops Drive Plant Mycorrhization in Organic Horticultural Systems. Microorganisms 2021, 9, 410. https://doi.org/10.3390/microorganisms9020410
Trinchera A, Testani E, Roccuzzo G, Campanelli G, Ciaccia C. Agroecological Service Crops Drive Plant Mycorrhization in Organic Horticultural Systems. Microorganisms. 2021; 9(2):410. https://doi.org/10.3390/microorganisms9020410
Chicago/Turabian StyleTrinchera, Alessandra, Elena Testani, Giancarlo Roccuzzo, Gabriele Campanelli, and Corrado Ciaccia. 2021. "Agroecological Service Crops Drive Plant Mycorrhization in Organic Horticultural Systems" Microorganisms 9, no. 2: 410. https://doi.org/10.3390/microorganisms9020410
APA StyleTrinchera, A., Testani, E., Roccuzzo, G., Campanelli, G., & Ciaccia, C. (2021). Agroecological Service Crops Drive Plant Mycorrhization in Organic Horticultural Systems. Microorganisms, 9(2), 410. https://doi.org/10.3390/microorganisms9020410