
The Natural History of Integrons

 ,
,  and
and 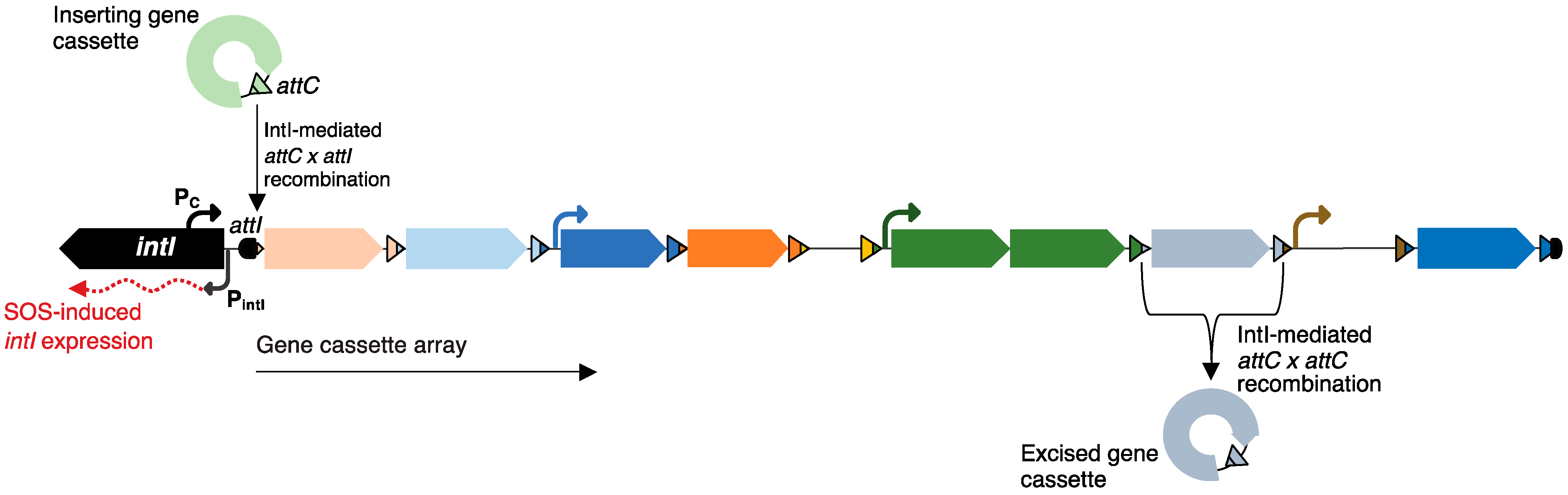{kind=link}
{kind=link}
Abstract
1. Introduction
2. Integrons: Beyond the Clinical Environment
2.1. Diversity and Distribution of Integrons
2.2. Environmental Clustering of Integrons
2.3. Functional Diversity of Gene Cassettes
3. Integrons in the Anthropocene
3.1. Mobilisation and the Rise of Class 1 Integrons
3.2. Success of Class 1 Integrons
3.3. Ongoing Integron-Driven Evolution in the Anthropocene
4. The Future of Integron Evolution
4.1. Natural Environments
4.2. Clinical Environments
4.3. Integrons as a Biotechnological Resource
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Stokes, H.W.; Hall, R.M. A novel family of potentially mobile DNA elements encoding site-specific gene-integration functions: Integrons. Mol. Microbiol. 1989, 3, 1669–1683. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Recchia, G.D.; Scaramuzzi, C.; Collis, C.M.; Stokes, H.; Hall, R.M. Definition of the attI1 site of class 1 integrons. Microbiology 2000, 146, 2855–2864. [Google Scholar] [CrossRef] [PubMed]
- Collis, C.M.; Hall, R.M. Expression of antibiotic resistance genes in the integrated cassettes of integrons. Antimicrob. Agents Chemother. 1995, 39, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, C.; Brassard, S.; Lapointe, J.; Roy, P.H. Diversity and relative strength of tandem promoters for the antibiotic-resistance genes of several integron. Gene 1994, 142, 49–54. [Google Scholar] [CrossRef]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef]
- Gillings, M.R. Integrons: Past, present, and future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Cury, J.; Jové, T.; Touchon, M.; Néron, B.; Rocha, E.P. Identification and analysis of integrons and cassette arrays in bacterial genomes. Nucleic Acids Res. 2016, 44, 4539–4550. [Google Scholar] [CrossRef]
- Cambray, G.; Guerout, A.-M.; Mazel, D. Integrons. Annu. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Tetu, S.G.; Gillings, M.R. Predicting the taxonomic and environmental sources of integron gene cassettes using structural and sequence homology of attC sites. Commun. Biol. 2021, 4, 946. [Google Scholar] [CrossRef]
- Gillings, M.R.; Holley, M.P.; Stokes, H.W.; Holmes, A.J. Integrons in Xanthomonas: A source of species genome diversity. Proceedings of the National Academy of Sciences of the United States of America 2005, 102, 4419–4424. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Guerout, A.-M.; Biskri, L.; Bouige, P.; Mazel, D. Comparative analysis of superintegrons: Engineering extensive genetic diversity in the Vibrionaceae. Genome Res. 2003, 13, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Vaisvila, R.; Morgan, R.D.; Posfai, J.; Raleigh, E.A. Discovery and distribution of super-integrons among Pseudomonads. Mol. Microbiol. 2001, 42, 587–601. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Guerout, A.-M.; Ploncard, P.; Dychinco, B.; Davies, J.; Mazel, D. The evolutionary history of chromosomal super-integrons provides an ancestry for multiresistant integrons. Proc. Natl. Acad. Sci. USA 2001, 98, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Michael, C.A.; Labbate, M. Gene cassette transcription in a large integron-associated array. BMC Genet. 2010, 11, 82. [Google Scholar] [CrossRef]
- Tansirichaiya, S.; Mullany, P.; Roberts, A.P. Promoter activity of ORF-less gene cassettes isolated from the oral metagenome. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Guerin, É.; Da Re, S.; González-Zorn, B.; Ploy, M.-C.; Barbé, J.; Mazel, D.; Erill, I. Prevalence of SOS-mediated control of integron integrase expression as an adaptive trait of chromosomal and mobile integrons. Mob. DNA 2011, 2, 6. [Google Scholar] [CrossRef]
- Guerin, É.; Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Erill, I.; Da Re, S.; Gonzalez-Zorn, B.; Barbé, J.; Ploy, M.-C.; Mazel, D. The SOS response controls integron recombination. Science 2009, 324, 1034. [Google Scholar] [CrossRef]
- Strugeon, E.; Tilloy, V.; Ploy, M.-C.; Re, S.D.; Davies, J.E. The stringent response promotes antibiotic resistance dissemination by regulating integron integrase expression in biofilms. mBio 2016, 7, e00868-00816. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Geoghegan, J.L.; Alroy, J.; Gillings, M.R. High diversity and rapid spatial turnover of integron gene cassettes in soil. Environ. Microbiol. 2019, 21, 1567–1574. [Google Scholar] [CrossRef]
- Elsaied, H.; Stokes, H.; Nakamura, T.; Kitamura, K.; Fuse, H.; Maruyama, A. Novel and diverse integron integrase genes and integron-like gene cassettes are prevalent in deep-sea hydrothermal vents. Environ. Microbiol. 2007, 9, 2298–2312. [Google Scholar] [CrossRef]
- Antelo, V.; Giménez, M.; Azziz, G.; Valdespino-Castillo, P.; Falcón, L.I.; Ruberto, L.A.M.; Mac Cormack, W.P.; Mazel, D.; Batista, S. Metagenomic strategies identify diverse integron-integrase and antibiotic resistance genes in the Antarctic environment. MicrobiologyOpen 2021, 10, e1219. [Google Scholar] [CrossRef]
- Dias, M.F.; de Castro, G.M.; de Paiva, M.C.; de Paula Reis, M.; Facchin, S.; do Carmo, A.O.; Alves, M.S.; Suhadolnik, M.L.; de Moraes Motta, A.; Henriques, I. Exploring antibiotic resistance in environmental integron-cassettes through intI-attC amplicons deep sequencing. Braz. J. Microbiol. 2021, 52, 363–372. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.-A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, T.M.; Penesyan, A.; Pritchard, A.; Qi, Q.; Rajabal, V.; Tetu, S.G.; Gillings, M.R. Methods for the targeted sequencing and analysis of integrons and their gene cassettes from complex microbial communities. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- MacDonald, D.; Demarre, G.; Bouvier, M.; Mazel, D.; Gopaul, D.N. Structural basis for broad DNA-specificity in integron recombination. Nature 2006, 440, 1157–1162. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, M.; Demarre, G.; Mazel, D. Integron cassette insertion: A recombination process involving a folded single strand substrate. EMBO J. 2005, 24, 4356–4367. [Google Scholar] [CrossRef]
- Biskri, L.; Bouvier, M.; Guérout, A.-M.; Boisnard, S.; Mazel, D. Comparative study of class 1 integron and Vibrio cholerae superintegron integrase activities. J. Bacteriol. 2005, 187, 1740–1750. [Google Scholar] [CrossRef] [PubMed]
- Koenig, J.E.; Bourne, D.G.; Curtis, B.; Dlutek, M.; Stokes, H.W.; Doolittle, W.F.; Boucher, Y. Coral-mucus-associated Vibrio integrons in the Great Barrier Reef: Genomic hotspots for environmental adaptation. ISME J. 2011, 5, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Boucher, Y.; Labbate, M.; Koenig, J.E.; Stokes, H.W. Integrons: Mobilizable platforms that promote genetic diversity in bacteria. Trends Microbiol. 2007, 15, 301–309. [Google Scholar] [CrossRef]
- Buongermino Pereira, M.; Österlund, T.; Eriksson, K.M.; Backhaus, T.; Axelson-Fisk, M.; Kristiansson, E. A comprehensive survey of integron-associated genes present in metagenomes. BMC Genom. 2020, 21, 495. [Google Scholar] [CrossRef]
- Iqbal, N.; Guérout, A.-M.; Krin, E.; Le Roux, F.; Mazel, D. Comprehensive functional analysis of the 18 Vibrio cholerae N16961 toxin-antitoxin systems substantiates their role in stabilizing the superintegron. J. Bacteriol. 2015, 197, 2150–2159. [Google Scholar] [CrossRef] [PubMed]
- Szekeres, S.; Dauti, M.; Wilde, C.; Mazel, D.; Rowe-Magnus, D.A. Chromosomal toxin–antitoxin loci can diminish large-scale genome reductions in the absence of selection. Mol. Microbiol. 2007, 63, 1588–1605. [Google Scholar] [CrossRef] [PubMed]
- Guérout, A.-M.; Iqbal, N.; Mine, N.; Ducos-Galand, M.; Van Melderen, L.; Mazel, D. Characterization of the phd-doc and ccd toxin-antitoxin cassettes from Vibrio superintegrons. J. Bacteriol. 2013, 195, 2270–2283. [Google Scholar] [CrossRef] [PubMed]
- Wilbaux, M.; Mine, N.; Guérout, A.-M.; Mazel, D.; Van Melderen, L. Functional interactions between coexisting toxin-antitoxin systems of the ccd family in Escherichia coli O157: H7. J. Bacteriol. 2007, 189, 2712–2719. [Google Scholar] [CrossRef] [PubMed]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S.; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef]
- Gillings, M.R.; Paulsen, I.T. Microbiology of the Anthropocene. Anthropocene 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Paulsen, I.T.; Sajjad, A.; Tetu, S.G.; Gillings, M.R. A novel family of Acinetobacter mega-plasmids are disseminating multi-drug resistance across the globe while acquiring location-specific accessory genes. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene cassettes and cassette arrays in mobile resistance integrons. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Chow, L.; Asher, A.J.; Waldron, L.S.; Gillings, M.R. Evolution of class 1 integrons: Mobilization and dispersal via food-borne bacteria. PLoS ONE 2017, 12, e0179169. [Google Scholar] [CrossRef] [PubMed]
- Sköld, O. Sulfonamide resistance: Mechanisms and trends. Drug Resist. Updates 2000, 3, 155–160. [Google Scholar] [CrossRef]
- Domingues, S.; Da Silva, G.J.; Nielsen, K.M. Global dissemination patterns of common gene cassette arrays in class 1 integrons. Microbiology 2015, 161, 1313–1337. [Google Scholar] [CrossRef] [PubMed]
- Sütterlin, S.; Bray, J.E.; Maiden, M.C.; Tano, E. Distribution of class 1 integrons in historic and contemporary collections of human pathogenic Escherichia coli. PLoS ONE 2020, 15, e0233315. [Google Scholar] [CrossRef]
- Vit, C.; Richard, E.; Fournes, F.; Whiteway, C.; Eyer, X.; Lapaillerie, D.; Parissi, V.; Mazel, D.; Loot, C. Cassette recruitment in the chromosomal Integron of Vibrio cholerae. Nucleic Acids Res. 2021, 49, 5654–5670. [Google Scholar] [CrossRef] [PubMed]
- Böhm, M.-E.; Razavi, M.; Flach, C.-F.; Larsson, D. A novel, integron-regulated, class C β-lactamase. Antibiotics 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Böhm, M.-E.; Razavi, M.; Marathe, N.P.; Flach, C.-F.; Larsson, D.J. Discovery of a novel integron-borne aminoglycoside resistance gene present in clinical pathogens by screening environmental bacterial communities. Microbiome 2020, 8, 114. [Google Scholar] [CrossRef]
- Marathe, N.P.; Berglund, F.; Razavi, M.; Pal, C.; Dröge, J.; Samant, S.; Kristiansson, E.; Larsson, D.J. Sewage effluent from an Indian hospital harbors novel carbapenemases and integron-borne antibiotic resistance genes. Microbiome 2019, 7, 97. [Google Scholar] [CrossRef]
- Razavi, M.; Marathe, N.P.; Gillings, M.R.; Flach, C.-F.; Kristiansson, E.; Larsson, D.J. Discovery of the fourth mobile sulfonamide resistance gene. Microbiome 2017, 5, 160. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Geoghegan, J.L.; Tetu, S.G.; Gillings, M.R. The peril and promise of integrons: Beyond antibiotic resistance. Trends Microbiol. 2020, 28, 455–464. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Gillings, M.; Simonet, P.; Stekel, D.; Banwart, S.; Penuelas, J. Microbial mass movements. Science 2017, 357, 1099–1100. [Google Scholar] [CrossRef]
- Escudero, J.A.; Loot, C.; Mazel, D. Integrons as adaptive devices. In Molecular Mechanisms of Microbial Evolution; Rampelotto, P.H., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 199–239. [Google Scholar]
- Souque, C.; Escudero, J.A.; MacLean, R.C. Integron activity accelerates the evolution of antibiotic resistance. eLife 2021, 10, e62474. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, K. Drugs in the environment: Emission of drugs, diagnostic aids and disinfectants into wastewater by hospitals in relation to other sources–a review. Chemosphere 2001, 45, 957–969. [Google Scholar] [CrossRef]
- Campagnolo, E.R.; Johnson, K.R.; Karpati, A.; Rubin, C.S.; Kolpin, D.W.; Meyer, M.T.; Esteban, J.E.; Currier, R.W.; Smith, K.; Thu, K.M. Antimicrobial residues in animal waste and water resources proximal to large-scale swine and poultry feeding operations. Sci. Total Environ. 2002, 299, 89–95. [Google Scholar] [CrossRef]
- Chow, L.K.; Ghaly, T.M.; Gillings, M.R. A survey of sub-inhibitory concentrations of antibiotics in the environment. J. Environ. Sci. 2021, 99, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Elsaied, H.; Stokes, H.W.; Kitamura, K.; Kurusu, Y.; Kamagata, Y.; Maruyama, A. Marine integrons containing novel integrase genes, attachment sites, attI, and associated gene cassettes in polluted sediments from Suez and Tokyo Bays. ISME J. 2011, 5, 1162–1177. [Google Scholar] [CrossRef]
- Koenig, J.E.; Sharp, C.; Dlutek, M.; Curtis, B.; Joss, M.; Boucher, Y.; Doolittle, W.F. Integron gene cassettes and degradation of compounds associated with industrial waste: The case of the Sydney tar ponds. PLoS ONE 2009, 4, e5276. [Google Scholar] [CrossRef]
- Elsaied, H.; Stokes, H.W.; Yoshioka, H.; Mitani, Y.; Maruyama, A. Novel integrons and gene cassettes from a Cascadian submarine gas-hydrate-bearing core. FEMS Microbiol. Ecol. 2014, 87, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R.; Labbate, M.; Sajjad, A.; Giguère, N.J.; Holley, M.P.; Stokes, H. Mobilization of a Tn402-like class 1 integron with a novel cassette array via flanking miniature inverted-repeat transposable element-like structures. Appl. Environ. Microbiol. 2009, 75, 6002–6004. [Google Scholar] [CrossRef] [PubMed]
- Nivina, A.; Grieb, M.S.; Loot, C.; Bikard, D.; Cury, J.; Shehata, L.; Bernardes, J.; Mazel, D. Structure-specific DNA recombination sites: Design, validation, and machine learning-based refinement. Sci. Adv. 2020, 6, eaay2922. [Google Scholar] [CrossRef]
- Bikard, D.; Julie-Galau, S.; Cambray, G.; Mazel, D. The synthetic integron: An in vivo genetic shuffling device. Nucleic Acids Res. 2010, 38, e153. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A. Integrase-directed recovery of functional genes from genomic libraries. Nucleic Acids Res. 2009, 37, e118. [Google Scholar] [CrossRef][Green Version]
- Kan, A.; Joshi, N.S. Towards the directed evolution of protein materials. MRS Commun. 2019, 9, 441–455. [Google Scholar] [CrossRef]
- Folke, C.; Polasky, S.; Rockström, J.; Galaz, V.; Westley, F.; Lamont, M.; Scheffer, M.; Österblom, H.; Carpenter, S.R.; Chapin, F.S.; et al. Our future in the Anthropocene biosphere. Ambio 2021, 50, 834–869. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghaly, T.M.; Gillings, M.R.; Penesyan, A.; Qi, Q.; Rajabal, V.; Tetu, S.G. The Natural History of Integrons. Microorganisms 2021, 9, 2212. https://doi.org/10.3390/microorganisms9112212
Ghaly TM, Gillings MR, Penesyan A, Qi Q, Rajabal V, Tetu SG. The Natural History of Integrons. Microorganisms. 2021; 9(11):2212. https://doi.org/10.3390/microorganisms9112212
Chicago/Turabian StyleGhaly, Timothy M., Michael R. Gillings, Anahit Penesyan, Qin Qi, Vaheesan Rajabal, and Sasha G. Tetu. 2021. "The Natural History of Integrons" Microorganisms 9, no. 11: 2212. https://doi.org/10.3390/microorganisms9112212
APA StyleGhaly, T. M., Gillings, M. R., Penesyan, A., Qi, Q., Rajabal, V., & Tetu, S. G. (2021). The Natural History of Integrons. Microorganisms, 9(11), 2212. https://doi.org/10.3390/microorganisms9112212