Atlantic Forest Malaria: A Review of More than 20 Years of Epidemiological Investigation

 , , ,
, , ,  and
and 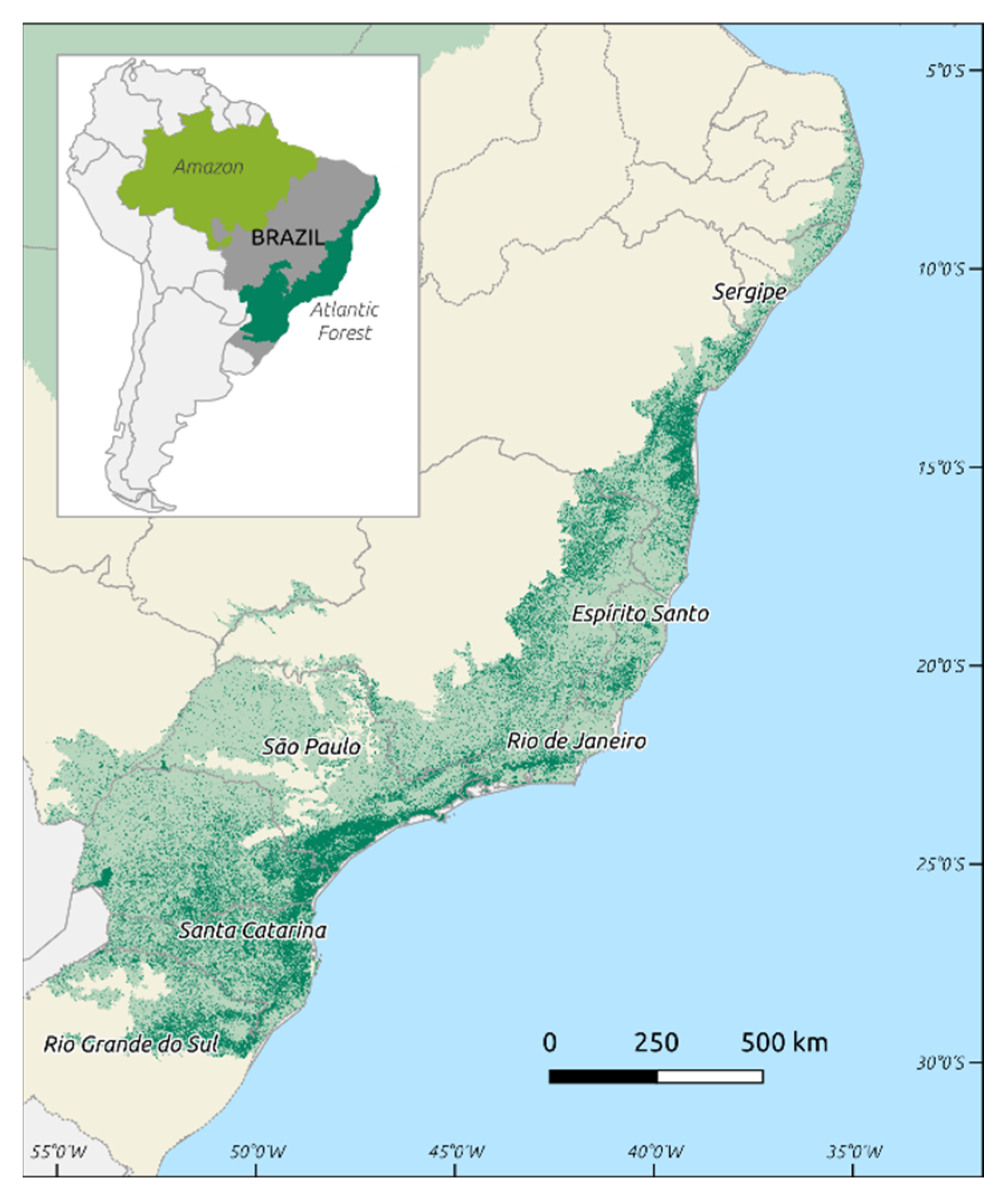{kind=link}
{kind=link}
Abstract
1. Introduction
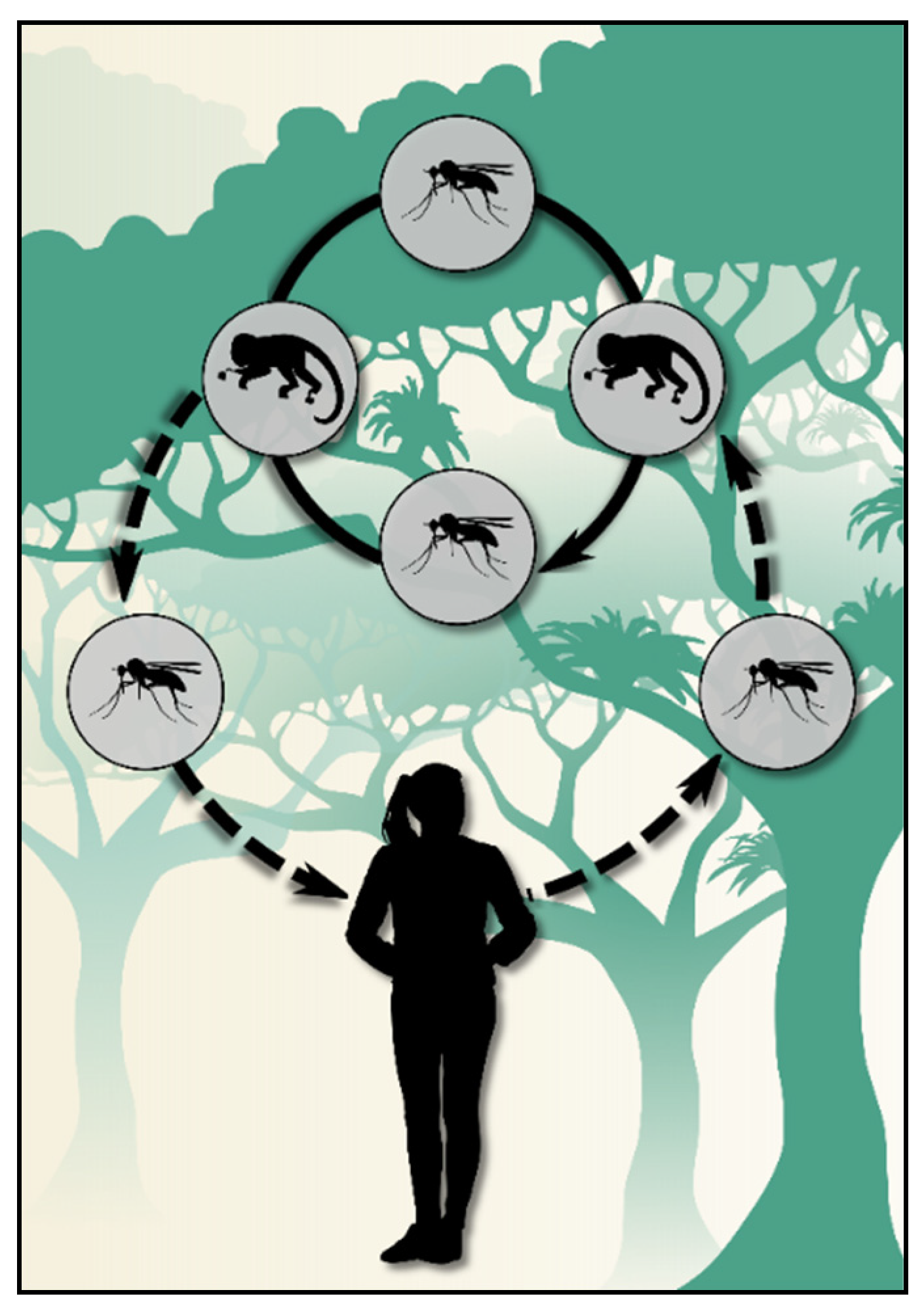
2. Simian Plasmodia and Their Relationship with Human Malaria in the Atlantic Forest
3. Vector Behavioral and Environmental Changes as Important Factors Related to Bromeliad-Malaria Transmission
4. Human Malaria
5. Surveillance
6. Concluding Remarks
7. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Malaria Report; World Health Organization: Geneva, Switzerland, 2019; 232p. [Google Scholar]
- Cerutti, C., Jr.; Boulos, M.; Coutinho, A.F.; Hatab, M.C.L.D.; Rezende, H.R.; Duarte, A.M.; Collins, H.; Malafronte, R.S. Epidemiologic aspects of the malaria transmission cycle in an area of very low incidence in Brazil. Malar. J. 2007, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.M.R.C.; Malafronte, R.S.; Cerutti, C., Jr.; Curado, I.; Paiva, B.R.; Maeda, A.Y.; Yamasaki, T.; Summa, M.E.L.; Neves, D.V.D.A.; Oliveira, S.G.; et al. Natural Plasmodium infections in Brazilian wild monkeys: Reservoirs for human infections? Acta Trop. 2008, 107, 79–85. [Google Scholar]
- Downs, W.G.; Pittendrigh, C.S. Bromelian malaria in Trinidad, British West Indies. Am. J. Trop. Med. 1946, 26, 47–66. [Google Scholar]
- Buery, J.C.; Rodrigues, P.T.; Natal, L.; Salla, L.C.; Loss, A.C.; Vicente, C.R.; Rezende, H.R.; Duarte, A.M.R.C.; Fux, B.; Malafronte, R.S.; et al. Mitochondrial genome of Plasmodium vivax/simium detected in an endemic region for malaria in the Atlantic Forest of Espírito Santo state, Brazil: Do mosquitoes, simians and humans harbour the same parasite? Malar. J. 2017, 16, 437. [Google Scholar] [CrossRef] [PubMed]
- Laporta, G.Z.; Burattini, M.N.; Levy, D.; Fukuya, L.A.; de Oliveira, T.M.; Maselli, L.M.; Conn, J.E.; Massad, E.; Bydlowski, S.P.; Sallum, M.A. Plasmodium falciparum in the southeastern Atlantic forest: A challenge to the bromeliad-malaria paradigm? Malar. J. 2015, 14, 181. [Google Scholar] [CrossRef]
- SOS Mata Atlântica, Instituto Nacional de Pesquisas Espaciais. Atlas dos Remanescentes Florestais da Mata Atlântica. Relatório Técnico do Período de 2017 a 2018. 2019. Available online: https://www.sosma.org.br/wp-content/uploads/2019/10/Atlas-mata-atlanticaDIGITAL.pdf (accessed on 5 November 2020).
- Buery, J.C.; Rezende, H.R.; Natal, L.; Silva, L.S.; Menezes, R.M.T.; Fux, B.; Malafronte, R.S.; Falqueto, A.; Cerutti, C., Jr. Ecological characterisation and infection of Anophelines (Diptera: Culicidae) of the Atlantic Forest in the southeast of Brazil over a 10 year period: Has the behaviour of the autochthonous malaria vector changed? Mem. Inst. Oswaldo Cruz 2018, 113, 111–118. [Google Scholar] [CrossRef]
- Rezende, H.R.; Soares, R.M.; Cerutti, C., Jr.; Alves, I.C.; Natal, D.; Urbinatti, P.R.; Yamasaki, T.; Falqueto, A.; Malafronte, R.S. Entomological characterization and natural infection of anophelines in an area of the Atlantic Forest with autochthonous malaria cases in mountainous region of Espírito Santo state, Brazil. Neotrop. Entomol. 2009, 38, 272–280. [Google Scholar] [CrossRef]
- Rezende, H.R.; Falqueto, A.; Urbinatti, P.R.; de Menezes, R.M.; Natal, D.; Cerutti, C., Jr. Comparative study of distribution of anopheline vectors (Diptera: Culicidae) in areas with and without malaria transmission in the highlands of an extra-Amazonian region in Brazil. J. Med. Entomol. 2013, 50, 598–602. [Google Scholar] [CrossRef]
- Kirchgatter, K.; Tubaki, R.M.; Malafronte, R.S.; Alves, I.C.; Lima, G.F.M.C.; Guimarães, L.O.; Zampaulo, R.A.; Wunderlich, G. Anopheles (Kerteszia) cruzii (Diptera: Culicidae) in peridomiciliary area during asymptomatic malaria transmission in the Atlantic Forest: Molecular identification of blood-meal sources indicates humans as primary intermediate hosts. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 403–409. [Google Scholar] [CrossRef]
- Demari-Silva, B.; Laporta, G.Z.; Oliveira, T.M.P.; Sallum, M.A.M. Plasmodium infection in Kerteszia cruzii (Diptera: Culicidae) in the Atlantic tropical rain forest, southeastern Brazil. Infect. Genet. Evol. 2020, 78, 104061. [Google Scholar] [CrossRef]
- Consoli, R.A.G.B.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Editora FIOCRUZ: Rio de Janeiro, Brazil, 1994; 228p, ISBN 85-85676-03-5. Available online: http://books.scielo.org (accessed on 30 October 2020).
- Forattini, O.P.; Gomes, A.C.; Santos, J.L.F.; Kakitani, I.; Marucci, D. Freqüência ao ambiente humano e dispersão de mosquitos Culicidae em área adjacente à Mata Atlântica primitiva da planície. Revista de Saúde Pública 1990, 2, 101–107. [Google Scholar]
- Goldman, I.F.; Qari, S.H.; Millet, P.G.; Collins, W.E.; Lal, A.A. Circumsporozoite protein gene of Plasmodium simium, a Plasmodium vivax-like monkey malaria parasite. Mol. Biochem. Parasitol. 1993, 57, 177–180. [Google Scholar] [CrossRef]
- Escalante, A.A.; Freeland, D.E.; Collins, W.E.; Lal, A.A. The evolution of primate malaria parasites based on the gene encoding cytochrome b from the linear mitochondrial genome. Proc. Natl. Acad. Sci. USA 1998, 95, 8124–8129. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, M.C.; Durand, P.; Gauthier, C.; Patot, S.; Billotte, N.; Menegon, M.; Severini, C.; Ayala, F.J.; Renaud, F. Meager genetic variability of the human malaria agent Plasmodium vivax. Proc. Natl. Acad. Sci. USA 2004, 101, 14455–14460. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Tazi, L.; Ayala, F.J. Plasmodium vivax: Recent world expansion and genetic identity to Plasmodium simium. Proc. Natl. Acad. Sci. USA 2005, 102, 15523–15528. [Google Scholar] [CrossRef]
- Tazi, L.; Ayala, F.J. Unresolved direction of host transfer of Plasmodium vivax v. P. simium and P. malariae v. P. brasilianum. Infect. Genet. Evol. 2011, 11, 209–221. [Google Scholar] [CrossRef]
- Lee, K.S.; Divis, P.C.S.; Zakaria, S.K.; Matusop, A.; Julin, R.A.; Conway, D.J.; Cox-Singh, J.; Singh, B. Plasmodium knowlesi: Reservoir hosts and tracking the emergence in humans and macaques. PLoS. Pathog. 2011, 7, 1002015. [Google Scholar] [CrossRef]
- Ta, T.H.; Hisam, S.; Lanza, M.; Jiram, A.I.; Ismail, N.; Rubio, J.M. First case of a naturally acquired human infection with Plasmodium cynomolgi. Malar. J. 2014, 13, 68. [Google Scholar] [CrossRef]
- Brasil, P.; Zalis, M.G.; Pina-Costa, A.; Siqueira, A.M.; Bianco Júnior, C.; Silva, S.; Pelaio-Machado, M.; Alvarenga, D.A.M.; Santelli, A.C.F.S.; Albuquerque, H.G.; et al. Outbreak of human malaria caused by Plasmodium simium in the Atlantic Forest in Rio de Janeiro: A molecular epidemiological investigation. Lancet Glob. Health 2017, 5, 1038–1046. [Google Scholar] [CrossRef]
- Abreu, F.V.S.; Santos, E.D.; Mello, A.R.L.; Gomes, L.R.; Alvarenga, D.A.M.A.; Gomes, M.Q.; Vargas, W.P.; Bianco-Júnior, C.; Pina-Costa, A.; Teixeira, D.S.; et al. Howler monkeys are the reservoir of malarial parasites causing zoonotic infections in the Atlantic forest of Rio de Janeiro. PLoS Negl. Trop. Dis. 2019, 13, e0007906. [Google Scholar] [CrossRef]
- Rodrigues, P.T.; Valdivia, H.O.; Oliveira, T.C.; Alves, J.M.P.; Duarte, A.M.R.C.; Cerutti, C., Jr.; Buery, J.C.; Brito, C.F.A.; Souza, J.C.; Hirano, Z.M.B.; et al. Human migration and the spread of malaria parasites to the New World. Sci. Rep. 2018, 8, 1993. [Google Scholar] [CrossRef] [PubMed]
- de Alvarenga, D.A.M.; Culleton, R.; de Pina-Costa, A.; Rodrigues, D.F.; Bianco, C., Jr.; Silva, S.; Nunes, A.J.D.; de Souza, J.C., Jr.; Hirano, Z.M.B.; Moreira, S.B.; et al. An assay for the identification of Plasmodium simium infection for diagnosis of zoonotic malaria in the Brazilian Atlantic Forest [published correction appears in Sci. Rep. 2019, 9, 17521]. Sci. Rep. 2018, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Buery, J.C.; Duarte, A.M.R.C.; Alencar, F.E.C.; Furieri, C.; Mendes, S.L.; Loss, A.C.; Vicente, C.R.; Fux, B.; Rezende, H.R.; Cravo, P.V.; et al. Detection of Plasmodium vivax in a liver sample of a howler-monkey: One evidence more in favour of the identity between Plasmodium simium and P. vivax. bioRxiv 2020. [Google Scholar] [CrossRef]
- Curado, I.; Duarte, A.M.R.C.; Lal, A.A.; Oliveira, S.G.; Kloetzel, J.K. Antibodies anti Bloodstream and Circumsporozoite Antigens (Plasmodium vivax and Plasmodium malariae/P. brasilianum) in areas of very low malaria endemicity in Brazil. Mem. Inst. Oswaldo Cruz 1997, 92, 235–243. [Google Scholar] [CrossRef]
- Arruda, M.E.; Nardin, E.H.; Nussenzweig, R.S.; Cochrane, A.H. Sero-epidemiological studies of malaria in indian tribes and monkeys of the Amazon Basin of Brazil. Am. J. Trop. Med. Hyg. 1989, 41, 379–385. [Google Scholar] [CrossRef]
- Deane, L.M. Simian malaria in Brazil. Mem. Inst. Oswaldo Cruz 1992, 87 (Suppl. S3), 1–20. [Google Scholar]
- Fandeur, T.; Volney, B.; Peneau, C.; De Thoisy, B. Monkeys of the rainforest in French Guiana are natural reservoirs for P. brasilianum/P. malariae. Parasitology 2000, 120, 11–21. [Google Scholar] [CrossRef]
- Volney, B.; Pouliquen, J.F.; De Thoisy, B.; Fandeur, T. A sero-epidemiological study of malaria in human and monkey population in French Guiana. Acta Trop. 2002, 82, 11–23. [Google Scholar] [CrossRef]
- Duarte, A.M.; Porto, M.A.; Curado, I.; Malafronte, R.S.; Hoffmann, E.H.; Oliveira, S.G.; Silva, A.M.; Kloetzel, J.K.; Gomes, A.C. Widespread occurrence of antibodies against circumsporozoite protein and against blood forms of Plasmodium vivax, P. falciparum and P. malariae in Brazilian wild monkeys. J. Med. Primatol. 2006, 35, 87–96. [Google Scholar] [CrossRef]
- Yamasaki, T.; Duarte, A.M.; Curado, I.; Summa, M.E.L.; Neves, D.V.D.A.; Wunderlich, G.; Malafronte, R.S. Detection of etiological agents of malaria in howler monkeys from Atlantic Forests, rescued in regions of São Paulo city, Brazil. J. Med. Primatol. 2011, 40, 392–400. [Google Scholar] [CrossRef]
- Erkenswick, G.A.; Watsa, M.; Pacheco, M.A.; Escalante, A.A.; Parker, P.G. Chronic Plasmodium brasilianum infections in wild Peruvian tamarins. PLoS ONE 2017, 12, e0184504. [Google Scholar] [CrossRef]
- Rondón, S.; León, C.; Link, A.; González, C. Prevalence of Plasmodium parasites in non-human primates and mosquitoes in areas with different degrees of fragmentation in Colombia. Malar. J. 2019, 18, 276. [Google Scholar] [CrossRef] [PubMed]
- Lalremruata, A.; Magris, M.; Vivas-Martinez, S.; Koehler, M.; Esen, M.; Kempaiah, P.; Jeyaraj, S.; Perkins, D.J.; Mordmüller, B.; Metzger, W.G. Natural infection of Plasmodium brasilianum in humans: Man and monkey share quartan malaria parasites in the Venezuelan Amazon. EBio Med. 2015, 2, 1186–1192. [Google Scholar] [CrossRef]
- Rayner, J.C. Plasmodium malariae malaria: From monkey to man? EBioMedicine 2015, 2, 1023–1024. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.M.; Plenderleith, L.J.; Hahn, B.H. Ape origins of human malaria. Annu. Rev. Microbiol. 2020, 74, 39–63. [Google Scholar] [CrossRef]
- Fonseca, F. Plasmodium of a primate of Brazil. Mem. Inst. Oswaldo Cruz 1951, 49, 543–553. [Google Scholar]
- Bueno, M.G. Pesquisa de Leishmania spp. e Plasmodium spp. em Primatas Neotropicais Provenientes de Regiões de Mata Atlântica eAmazônia Impactadas por Ações Antrópicas: Investigação In Situ e Ex Situ. Ph.D. Thesis, University of São Paulo, Sao Paulo, Brazil, 2012. [Google Scholar]
- de Alvarenga, D.A.M.; Pina-Costa, A.; Sousa, T.N.; Pissinatti, A.; Zalis, M.G.; Suaréz-Mutis, M.C.; Lourenço-de-Oliveira, R.; Brasil, P.; Daniel-Ribeiro, C.T.; Brito, C.F.A. Simian malaria in the Brazilian Atlantic forest: First description of natural infection of capuchin monkeys (Cebinae subfamily) by Plasmodium simium. Malar. J. 2015, 14, 81. [Google Scholar] [CrossRef]
- Deane, L.M.; Ferreira Neto, J.A.; Deane, M.P.; Silveira, I.P.S. Anopheles (Kerteszia) cruzii, a natural vector of the monkey malaria parasites, Plasmodium simium and Plasmodium brasilianum. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 647. [Google Scholar] [CrossRef]
- Costa, D.C.; Assis, G.M.P.; Silva, F.A.S.; Araújo, F.C.; de Souza Junior, J.C.; Braga Hirano, Z.M.; Kano, F.S.; de Sousa, T.N.; Carvalho, L.H.; Brito, C.F.A. Plasmodium simium, a Plasmodium vivax-related malaria parasite: Genetic variability of Duffy binding protein II and the Duffy antigen/receptor for chemokines. PLoS ONE 2015, 10, e0131339. [Google Scholar] [CrossRef]
- Mourier, T.; de Alvarenga, D.A.M.; Kaushik, A.; de Pina-Costa, A.; Douvropoulou, O.; Guan, Q.; Guzmán-Vega, F.J.; Forrester, S.; de Abreu, F.V.S.; Júnior, C.B.; et al. The genome of the zoonotic malaria parasite Plasmodium simium reveals adaptations to host-switching (preprint). bioRxiv 2019. [Google Scholar] [CrossRef]
- Buppan, P.; Putaporntip, C.; Pattanawong, U.; Seethamchai, S.; Jongwutiwes, S. Comparative detection of Plasmodium vivax and Plasmodium falciparum DNA in saliva and urine samples from symptomatic malaria patients in a low endemic area. Malar. J. 2010, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Prugnolle, F.; Durand, P.; Neel, C.; Ollomo, B.; Ayala, F.J.; Arnathau, C.; Etienne, L.; Mpoudi-Ngole, E.; Nkoghe, D.; Leroy, E.; et al. African great apes are natural hosts of multiple related malaria species, including Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2010, 107, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- De Nys, H.M.; Calvignac-Spencer, S.; Thiesen, U.; Boesch, C.; Wittig, R.M.; Mundry, R.; Leendertz, F.H. Age-related effects on malaria parasite infection in wild chimpanzees. Biol. Lett. 2013, 9, 20121160. [Google Scholar] [CrossRef] [PubMed]
- Kawai, S.; Sato, M.; Kato-Hayashi, N.; Kishi, H.; Huffman, M.A.; Maeno, Y.; Culleton, R.; Nakazawa, S. Detection of Plasmodium knowlesi DNA in the urine and faeces of a Japanese macaque (Macaca fuscata) over the course of an experimentally induced infection. Malar. J. 2014, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Siregar, J.E.; Faust, C.L.; Murdiyarso, L.S.; Rosmanah, L.; Saepuloh, U.; Dobson, A.P.; Iskandriati, D. Non-invasive surveillance for Plasmodium in reservoir macaque species. Malar. J. 2015, 14, 404. [Google Scholar] [CrossRef]
- Mapua, M.I.; Qablan, M.A.; Pomajbíková, K.; Petrželková, K.J.; Hůzová, Z.; Rádrová, J.; Votýpka, J.; Todd, A.; Jirků, M.; Leendertz, F.H.; et al. Ecology of malaria infections in western lowland gorillas inhabiting Dzanga Sangha Protected Areas, Central African Republic. Parasitology 2015, 142, 890–900. [Google Scholar] [CrossRef]
- Liu, W.; Sundararaman, S.A.; Loy, D.E.; Learn, G.H.; Li, Y.; Plenderleith, L.J.; Ndjango, J.-B.N.; Speede, S.; Atencia, R.; Cox, D.; et al. Multigenomic delineation of Plasmodium species of the Laverania subgenus infecting wild-living chimpanzees and gorillas. Genome Biol. Evol. 2016, 8, 1929–1939. [Google Scholar] [CrossRef]
- Dixit, J.; Zachariah, A.; Sajesh, P.K.; Chandramohan, B.; Shanmuganatham, V.; Karanth, K.P. Reinvestigating the status of malaria parasite (Plasmodium sp.) in Indian non-human primates. PLoS Negl. Trop. Dis. 2018, 12, e0006801. [Google Scholar] [CrossRef]
- Jirkú, M.; Pomajbíková, K.; Petrzelková, K.J.; Huzová, Z.; Modrý, D.; Lukes, J. Detection of Plasmodium spp. in human feces. Emerg. Infect. Dis. 2012, 18, 634–636. [Google Scholar] [CrossRef]
- Assis, G.M.P.; Alvarenga, D.A.M.; Costa, D.C.; Souza, J.C., Jr.; Hirano, Z.M.B.; Kano, F.S.; Sousa, T.N.; Brito, C.F.A. Detection of Plasmodium in faeces of the New World primate Alouatta clamitans. Mem. Inst. Oswaldo Cruz 2016, 111, 570–576. [Google Scholar] [CrossRef]
- Abkallo, H.M.; Liu, W.; Hokama, S.; Ferreira, P.E.; Nakazawa, S.; Maeno, Y.; Quang, N.T.; Kobayashi, N.; Kaneko, O.; Huffman, M.A.; et al. DNA from pre-erythrocytic stage malaria parasites is detectable by PCR in the faeces and blood of hosts. Int. J. Parasitol. 2014, 44, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Freitas, M.G.; Lourenço-De-Oliveira, R.; De Carvalho-Pinto, C.J.; Flores-Mendoza, C.; Silva-Do-Nascimento, T.F. Anopheline species complexes in Brazil. current knowledge of those related to malaria transmission. Mem. Inst. Oswaldo Cruz 1998, 93, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Secretaria de Estado da Saúde. Superintendência de Controle de Endemias. Informativo Programa de Malária: Vetores; SESA: São Paulo, Brazil, 2015. Available online: http://www.saude.sp.gov.br/sucen-superintendencia-de-controle-de-endemias/programas/malaria/vetore (accessed on 8 February 2016).
- Pinotti, M. The biological basis for the campaign against the malaria vectors of Brazil. Trans. R. Soc. Trop. Med. Hyg. 1951, 44, 663–682. [Google Scholar] [CrossRef]
- Rachou, R.G. Da infectibilidade dos anofelinos do subgênero Kerteszia pelos parasitas da malária humana. A Folha Médica 1946, 27, 181–183. [Google Scholar]
- Corrêa, R.R.; Forattini, O.P.; Guarita, O.F.; Rabello, E.X. Observações sobre o vôo do Anopheles (Kerteszia) cruzii e do Anopheles (Kerteszia) bellator, vetores de malária (Diptera, Culicidae). Arquivos de Higiene e Saúde Pública 1961, 26, 333–342. [Google Scholar]
- Ferreira, E.; Corrêa, R.R.; Tomich, A.; Sá, F.T. Estudo sobre o raio de vôo do Anopheles (Kerteszia) cruzii e do Anopheles (Kerteszia) bellator em Guaratuba, litoral do estado do Paraná, Brasil. Revista Brasileira de Malariologia e Doenças Tropicais 1969, 21, 819–822. [Google Scholar]
- Deane, L.M.; Deane, M.P.; Ferreira Neto, J.A. Studies on transmission of simian malaria and on a natural infection of man with Plasmodium simium in Brazil. Bull. World Health Organ. 1966, 35, 805–808. [Google Scholar]
- Carvalho-Pinto, C.J.; Lourenço-De-Oliveira, R. Isoenzimatic analysis of four Anopheles (Kerteszia) cruzii (Diptera: Culicidae) populations of Brazil. Mem. Inst. Oswaldo Cruz 2004, 99, 471–475. [Google Scholar] [CrossRef]
- Rona, L.D.P.; Carvalho-Pinto, C.J.; Gentile, C.; Grisard, E.C.; Peixoto, A.A. Assessing the molecular divergence between Anopheles (Kerteszia) cruzii populations from Brazil using the timeless gene: Further evidence of a species complex. Malar. J. 2009, 8, 60. [Google Scholar] [CrossRef]
- Rona, L.D.P.; Carvalho-Pinto, C.J.; Peixoto, A.A. Molecular evidence for the occurrence of a new sibling species within the Anopheles (Kerteszia) cruzii complex in south-east Brazil. Malar. J. 2010, 9, 33. [Google Scholar] [CrossRef]
- Dias, G.R.; Fujii, T.T.S.; Fogel, B.F.; Lourenço-de-Oliveira, R.; Silva-do-Nascimento, T.F.; Pitaluga, A.N.; Carvalho-Pinto, C.J.; Carvalho, A.B.; Peixoto, A.A.; Rona, L.D.P. Cryptic diversity in an Atlantic Forest malaria vector from the mountains of South-East Brazil. Parasites Vectors 2018, 11, 36. [Google Scholar] [CrossRef]
- Branquinho, M.S.; Marrelli, M.T.; Curado, I.; Natal, D.; Barata, J.M.S.; Tubaki, R.; Carréri-Bruno, G.C.; Menezes, R.T.; Kloetzel, J.K. Infecção do Anopheles (Kerteszia) cruzii por Plasmodium vivax e Plasmodium vivax variante VK247 nos municípios de São Vicente e Juquitiba, São Paulo. Rev. Panam Salud Publica/Pan Am. J. Public Health 1997, 2, 189–193. [Google Scholar]
- Duarte, A.M.R.C.; Pereira, D.M.; de Paula, M.B.; Fernandes, A.; Urbinatti, P.R.; Ribeiro, A.F.; Mello, M.H.S.H.; Matos, M.O.; Mucci, L.F.; Fernandes, L.N.; et al. Natural infection in anopheline species and its implications for autochthonous malaria in the Atlantic Forest in Brazil. Parasites Vectors 2013, 6, 58. [Google Scholar] [CrossRef]
- Deane, L.M.; Ferreira Neto, J.A.; Lima, M.M. The vertical dispersion of Anopheles (Kerteszia) cruzii in a forest in southern Brazil suggests that human cases of malaria of simian origin might be expected. Mem. Inst. Oswaldo Cruz 1984, 79, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.E.; Gentile, C.; Lopes, C.M.; Mello, R.P. Ecology of mosquitoes (Diptera: Culicidae) in areas of Serra do Mar State Park, State of São Paulo, Brazil: II—Habitat distribution. Mem. Inst. Oswaldo Cruz 2000, 95, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Forattini, O.P.; Kakitani, I.; Massad, E.; Gomes, A.C. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment: 1—Parity of blood seeking Anopheles (Kerteszia) in South-Eastern Brazil. Revista de Saúde Pública 1993, 27, 1–8. [Google Scholar]
- Forattini, O.P.; Lopes, O.S.; Rabello, E.X. Investigações sobre o comportamento de formas adultas de mosquitos silvestres. Rev. Saúde Pública 1968, 2, 111–173. [Google Scholar] [PubMed]
- Deane, L.M. Malaria vectors in Brazil. Mem. Inst. Oswaldo Cruz 1986, 81, 5–14. [Google Scholar] [CrossRef][Green Version]
- Ueno, H.M.; Forattini, O.P.; Kakitani, I. Distribuição vertical e sazonal de Anopheles (Kerteszia) em Ilha Comprida, SP. Rev. Saúde Pública 2007, 41, 269–275. [Google Scholar]
- Medeiros-Sousa, A.R.; de Oliveira Christe, R.; de Castro Duarte, A.M.R.; Mucci, L.F.; Ceretti-Junior, W.; Marrelli, M.T. Effects of anthropogenic landscape changes on the abundance and acrodendrophily of Anopheles (Kerteszia) cruzii, the main vector of malaria parasites in the Atlantic Forest in Brazil. Malar. J. 2019, 18, 110. [Google Scholar] [CrossRef]
- Maselli, L.M.F.; Levy, D.; Laporta, G.Z.; Monteiro, A.M.; Fukuya, L.A.; Ferreira-da-Cruz, M.F.; Daniel-Ribeiro, C.T.; Dorlhiac-Llacer, P.E.; Sallum, M.A.M.; Bydlowski, S.P. Detection of Plasmodium falciparum and Plasmodium vivax subclinical infection in non-endemic region: Implications for blood transfusion and malaria epidemiology. Malar. J. 2014, 13, 224. [Google Scholar] [CrossRef] [PubMed]
- Curado, I.; Malafronte, R.S.; Duarte, A.M.R.C.; Kirchgatter, K.; Branquinho, M.S.; Galati, E.A.B. Malaria epidemiology in low-endemicity areas of the Atlantic Forest in the Vale do Ribeira, São Paulo, Brazil. Acta Trop. 2006, 100, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Multini, L.C.; Souza, A.L.S.; Marrelli, M.T.; Wilke, A.B.B. The influence of anthropogenic habitat fragmentation on the genetic structure and diversity of the malaria vector Anopheles cruzii (Diptera: Culicidae). Sci. Rep. 2020, 10, 18018. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo, T.S.; Lorenz, C.; Chiaravalloti-Neto, F.; Sallum, M.A.M. Kerteszia cruzii and extra-Amazonian malaria in Brazil: Challenges due to climate change in the Atlantic Forest. Infect. Genet. Evol. 2020, 85, 104456. [Google Scholar] [CrossRef] [PubMed]
- Benchimol, J.L.; Sá, M.R. Adolpho Lutz—Febre Amarela, Malária e Protozoologia—v.2, Livro 1; Fiocruz: Rio de Janeiro, Brazil, 2005. [Google Scholar]
- Pittendrigh, C.S. The bromeliad—Anopheles—malaria complex in Trinidad. I—The bromeliad flora. Evolution 1948, 1, 58–89. [Google Scholar] [CrossRef]
- Ministério da Saúde. Superintendência de Campanhas de Saúde Pública. Departamento de Erradicação e Controle de Endemias. O controle de Endemias no Brasil: De 1979 a 1984; SUCAM: Brasília, Brazil, 1985. [Google Scholar]
- Barata, R.B. Malária e Seu Controle, 1st ed.; Hucitec: São Paulo, Brazil, 1998. [Google Scholar]
- De Alencar, F.E.C.; Malafronte, R.S.; Cerutti, C., Jr.; Fernandes, L.N.; Buery, J.C.; Fux, B.; Rezende, H.R.; Miranda, A.E. Reassessment of asymptomatic carriers of Plasmodium spp. in an endemic area with a very low incidence of malaria in extra-Amazonian Brazil. Malar. J. 2017, 6, 452. [Google Scholar] [CrossRef]
- De Alencar, F.E.C.; Malafronte, R.S.; Cerutti, C., Jr.; Fernandes, L.N.; Buery, J.C.; Fux, B.; Rezende, H.R.; Duarte, A.M.R.C.; Medeiros-Sousa, A.R.; Miranda, A.E. Assessment of asymptomatic Plasmodium spp. infection by detection of parasite DNA in residents of an extra-Amazonian region of Brazil. Malar. J. 2018, 17, 113. [Google Scholar] [CrossRef]
- Miguel, R.M.; Peiter, P.C.; Albuquerque, H.; Coura, J.R.; Moza, P.G.; Costa, A.P.; Brasil, P.; Suárez-Mutis, M.C. Malaria in the state of Rio de Janeiro, Brazil, an Atlantic Forest area: An assessment using the health surveillance service. Mem. Inst. Oswaldo Cruz 2014, 109, 634–640. [Google Scholar] [CrossRef]
- Amir, A.; Cheong, F.W.; De Silva, J.R.; Lau, Y.L. Diagnostic tools in childhood malaria. Parasites Vectors 2018, 11, 53. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buery, J.C.; de Alencar, F.E.C.; Duarte, A.M.R.d.C.; Loss, A.C.; Vicente, C.R.; Ferreira, L.M.; Fux, B.; Medeiros, M.M.; Cravo, P.; Arez, A.P.; et al. Atlantic Forest Malaria: A Review of More than 20 Years of Epidemiological Investigation. Microorganisms 2021, 9, 132. https://doi.org/10.3390/microorganisms9010132
Buery JC, de Alencar FEC, Duarte AMRdC, Loss AC, Vicente CR, Ferreira LM, Fux B, Medeiros MM, Cravo P, Arez AP, et al. Atlantic Forest Malaria: A Review of More than 20 Years of Epidemiological Investigation. Microorganisms. 2021; 9(1):132. https://doi.org/10.3390/microorganisms9010132
Chicago/Turabian StyleBuery, Julyana Cerqueira, Filomena Euridice Carvalho de Alencar, Ana Maria Ribeiro de Castro Duarte, Ana Carolina Loss, Creuza Rachel Vicente, Lucas Mendes Ferreira, Blima Fux, Márcia Melo Medeiros, Pedro Cravo, Ana Paula Arez, and et al. 2021. "Atlantic Forest Malaria: A Review of More than 20 Years of Epidemiological Investigation" Microorganisms 9, no. 1: 132. https://doi.org/10.3390/microorganisms9010132
APA StyleBuery, J. C., de Alencar, F. E. C., Duarte, A. M. R. d. C., Loss, A. C., Vicente, C. R., Ferreira, L. M., Fux, B., Medeiros, M. M., Cravo, P., Arez, A. P., & Cerutti Junior, C. (2021). Atlantic Forest Malaria: A Review of More than 20 Years of Epidemiological Investigation. Microorganisms, 9(1), 132. https://doi.org/10.3390/microorganisms9010132