Characterization of the Bacterial Biofilm Communities Present in Reverse-Osmosis Water Systems for Haemodialysis

 ,
, 
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Scanning Electron Microscopy
2.3. Total and Heterotrophic Bacterial Counts
2.4. Polymerase Chain Reaction (PCR)-Denaturing Gradient Gel Electrophoresis (DGGE) and Analysis of DGGE Profiles
2.5. Genomic DNA Extraction and 16S rRNA Gene Massive Sequencing
2.6. Bacterial Isolation
2.7. Identification of Heterotrophic Aerobic Bacteria
2.8. Antimicrobial Susceptibility Assay
2.9. Statistical Analysis
3. Results
3.1. Scanning Electron Microscopy (SEM) Observation of RO Membranes
3.2. Total Cell and Viable Heterotrophic Bacterial Counts
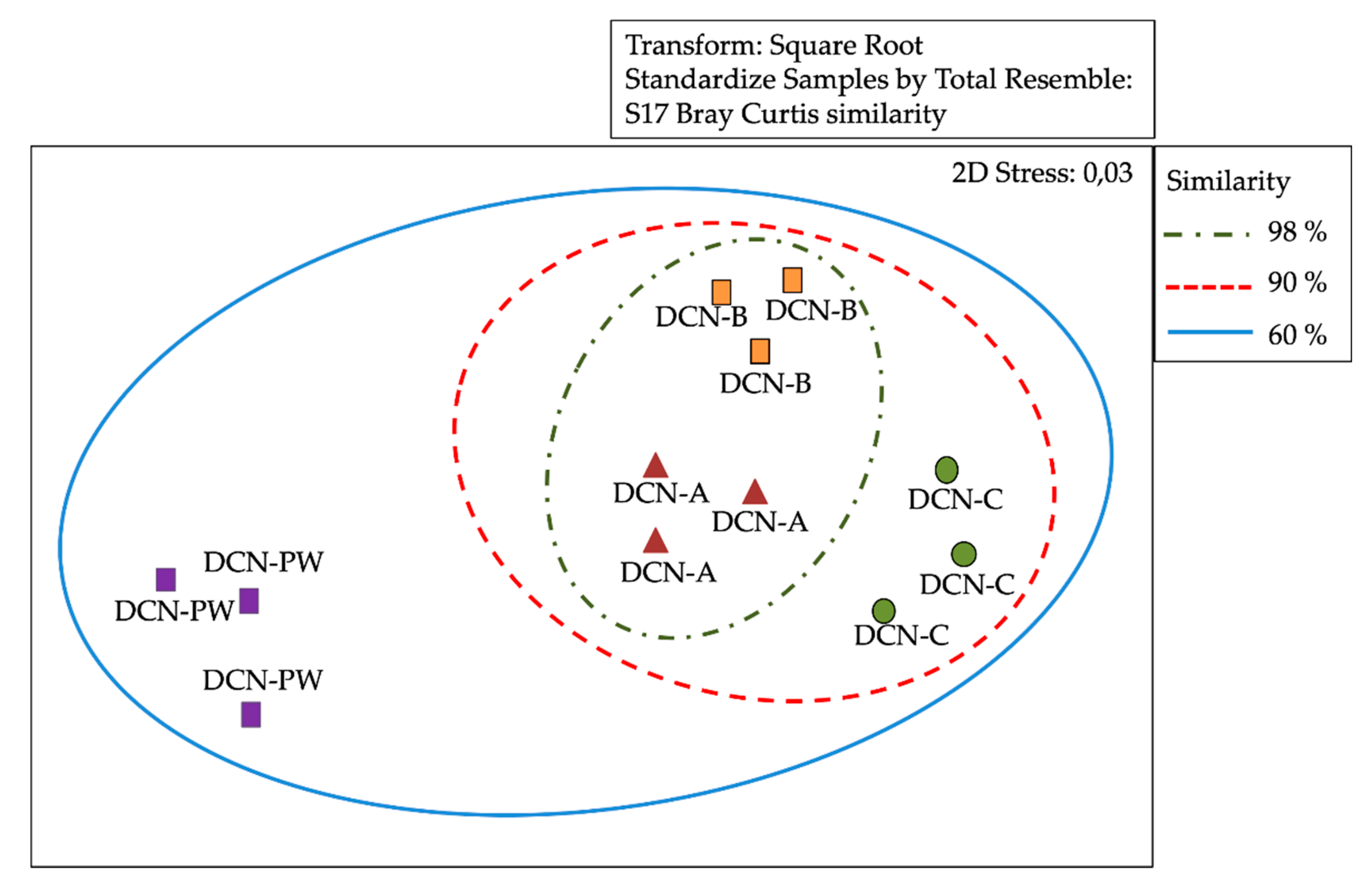
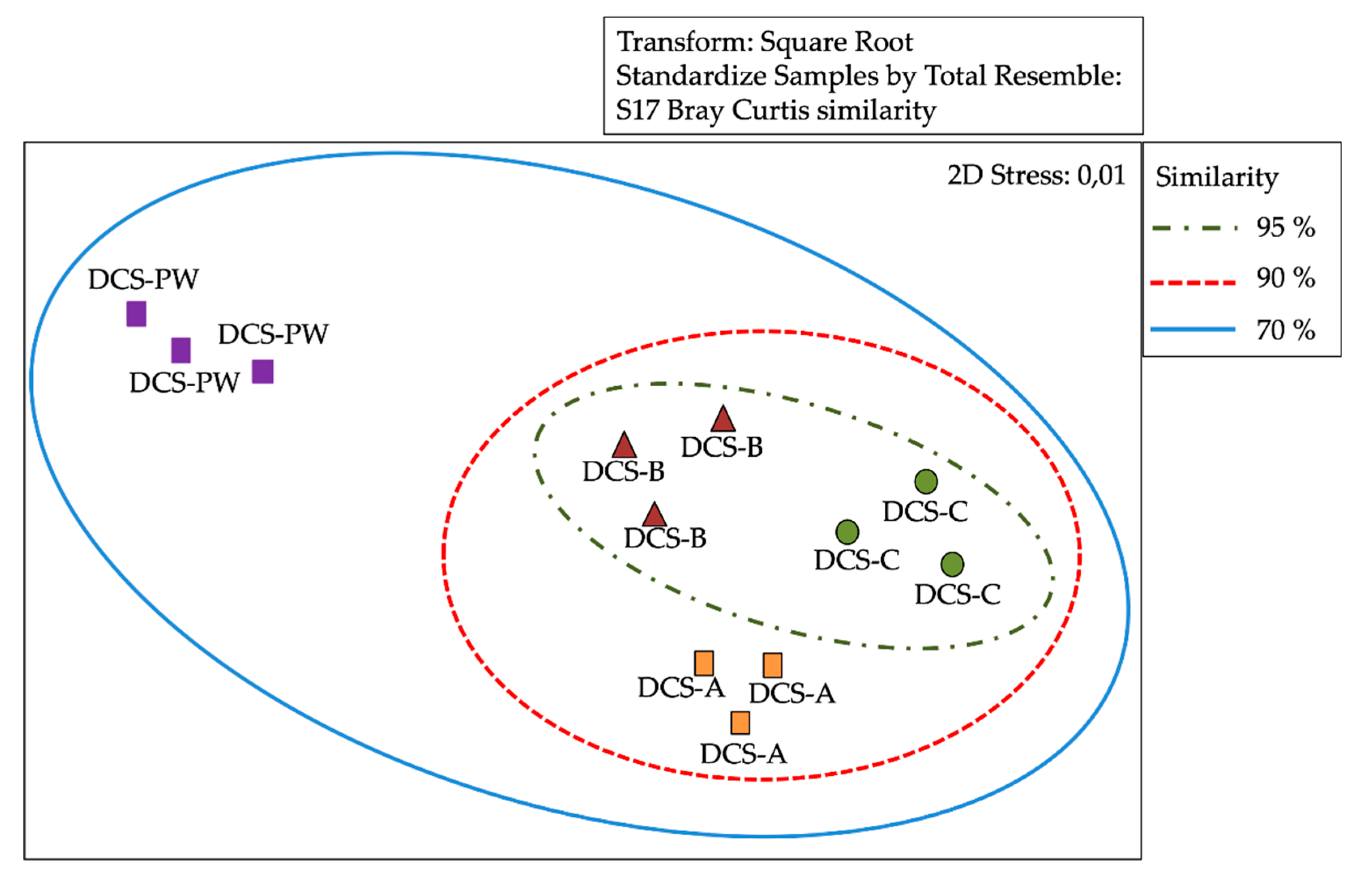
3.3. Analysis of Similarity of Bacterial Communities by DGGE
3.4. Analysis of the Bacterial Communities’ Composition
3.5. Identification of Isolates
3.6. Antimicrobial Susceptibility Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.; Yang, C.W. Chronic kidney disease: Global dimension and perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Sato, K.; Kawakami, S.; Kiyomoto, M.; Takei, H.; Suzuki, T.; Genei, H.; Nakata, H.; Iino, Y.; Katayama, Y. The performance evaluation of endotoxin retentive filters in haemodialysis. J. Nippon Med. Sch. 2011, 78, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Bereschenko, L.A.; Heilig, G.H.; Nederlof, M.M.; van Loosdrecht, M.C.; Stams, A.J.; Euverink, G.J. Molecular characterization of the bacterial communities in the different compartments of a full-scale reverse-osmosis water purification plant. Appl. Environ. Microbiol. 2008, 74, 5297–5304. [Google Scholar] [CrossRef] [PubMed]
- Coulliette, A.D.; Arduino, M.J. Hemodialysis and water quality. Semin. Dial. 2013, 26, 427–438. [Google Scholar] [CrossRef]
- Perez-Garcia, R.; Garcia, M.R.; Gonzalez, P.E.; Solozabal, C.C.; Ramirez, C.R.; Martin-Rabadan, P.; Sobrino, P.P.E.; Gallego, P.O.; Dominguez, J.; de la Cueva, M.E.; et al. Guideline for dialysate quality of Spanish Society of Nephrology (second edition, 2015). Nefrologia 2016, 36, e1–e52. [Google Scholar] [CrossRef][Green Version]
- Flemming, H.-C.; Schaule, G.; Griebe, T.; Schmitt, J.; Tamachkiarowa, A. Biofouling—The achilles heel of membrane processes. Desalination 1997, 113, 215–225. [Google Scholar] [CrossRef]
- Herzberg, M.; Elimelech, M. Biofouling of reverse osmosis membranes: Role of biofilm-enhanced osmotic pressure. J. Membr. Sci. 2007, 295, 11–20. [Google Scholar] [CrossRef]
- Khan, M.M.; Stewart, P.S.; Moll, D.J.; Mickols, W.E.; Nelson, S.E.; Camper, A.K. Characterization and effect of biofouling on polyamide reverse osmosis and nanofiltration membrane surfaces. Biofouling 2011, 27, 173–183. [Google Scholar] [CrossRef]
- Arduino, M.J. Dialysis-Associated Complications and Their Control. In Bennett and Brachman’s Hospital Infections, 5th ed.; Jarvis, W.R., Ed.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007. [Google Scholar]
- Park, S.K.; Hu, J.Y. Assessment of the extent of bacterial growth in reverse osmosis system for improving drinking water quality. J. Environ. Sci. Health Part A 2010, 45, 968–977. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, X.; Zhang, M.; Wang, Y.; Lv, T.; Zhang, S.; Yu, X. Profiling Total Viable Bacteria in a Hemodialysis Water Treatment System. J. Microbiol. Biotechnol. 2017, 27, 995–1004. [Google Scholar] [CrossRef]
- Guzmán-Fierro, V.; Moraga, R.; Leon, C.; Campos, V.; Smith, C.; Mondaca, M. Isolation and characterization of an aerobic bacterial consortium able to degrade roxarsone. Int. J. Environ. Sci. Technol. 2015, 12. [Google Scholar] [CrossRef]
- Mafla, S.; Moraga, R.; Leon, C.G.; Guzman-Fierro, V.G.; Yanez, J.; Smith, C.T.; Mondaca, M.A.; Campos, V.L. Biodegradation of roxarsone by a bacterial community of underground water and its toxic impact. World J. Microbiol. Biotechnol. 2015, 31, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.L.; Valenzuela, C.; Yarza, P.; Kampfer, P.; Vidal, R.; Zaror, C.; Mondaca, M.A.; Lopez-Lopez, A.; Rossello-Mora, R. Pseudomonas arsenicoxydans sp. nov., an arsenite-oxidizing strain isolated from the Atacama desert. Syst. Appl. Microbiol. 2010, 33, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Leon, C.G.; Moraga, R.; Valenzuela, C.; Gugliandolo, C.; Lo Giudice, A.; Papale, M.; Vilo, C.; Dong, Q.; Smith, C.T.; Rossello-Mora, R.; et al. Effect of the natural arsenic gradient on the diversity and arsenic resistance of bacterial communities of the sediments of Camarones River (Atacama Desert, Chile). PLoS ONE 2018, 13, e0195080. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Satoh, N.; Nakamura, M.; Horita, S.; Seki, G.; Moriya, K. Bacteremia in hemodialysis patients. World J. Nephrol. 2016, 5, 489–496. [Google Scholar] [CrossRef]
- Fisher, M.; Golestaneh, L.; Allon, M.; Abreo, K.; Mokrzycki, M.H. Prevention of Bloodstream Infections in Patients Undergoing Hemodialysis. Clin. J. Am. Soc. Nephrol. 2020, 15, 132–151. [Google Scholar] [CrossRef]
- Suzuki, M.N.; Fregonesi, B.M.; Machado, C.S.; Zagui, G.S.; Kusumota, L.; Suzuki, A.K.; da Costa, J.A.C.; Llopart, J.S.; Nadal, M.; Domingo, J.L.; et al. Hemodialysis water parameters as predisposing factors for anemia in patients in dialytic treatment: Application of mixed regression models. Biol. Trace Elem. Res. 2019, 190, 30–37. [Google Scholar] [CrossRef]
- Al Ashhab, A.; Sweity, A.; Bayramoglu, B.; Herzberg, M.; Gillor, O. Biofouling of reverse osmosis membranes: Effects of cleaning on biofilm microbial communities, membrane performance, and adherence of extracellular polymeric substances. Biofouling 2017, 33, 397–409. [Google Scholar] [CrossRef]
- Nagaraj, V.; Skillman, L.; Ho, G.; Li, D.; Gofton, A. Characterisation and comparison of bacterial communities on reverse osmosis membranes of a full-scale desalination plant by bacterial 16S rRNA gene metabarcoding. NPJ Biofilms Microbiomes 2017, 3, 13. [Google Scholar] [CrossRef]
- Al Juboori, R.Y.T. Biofouling in RO System: Mechanisms, monitoring and controlling. Desalination 2012, 302, 1–23. [Google Scholar] [CrossRef]
- Qureshi, B.; Zubair, S.; Sheikh, A.K.; Bhujle, A.; Dubowsky, S. Design and performance evaluation of reverse osmosis desalination systems: An emphasis on fouling modeling. Appl. Therm. Eng. 2013, 60, 208–217. [Google Scholar] [CrossRef]
- Vitzilaiou, E.; Stoica, I.M.; Knøchel, S. Microbial biofilm communities on Reverse Osmosis membranes in whey water processing before and after cleaning. J. Membr. Sci. 2019, 587, 117174. [Google Scholar] [CrossRef]
- Nakano, M.; Deguchi, T.; Kawamura, T.; Yasuda, M.; Kimura, M.; Okano, Y.; Kawada, Y. Mutations in the gyrA and parC genes in fluoroquinolone-resistant clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1997, 41, 2289–2291. [Google Scholar] [CrossRef] [PubMed]
- Shahryari, A.; Nikaeen, M.; Hatamzadeh, M.; Vahid Dastjerdi, M.; Hassanzadeh, A. Evaluation of bacteriological and chemical quality of dialysis water and fluid in Isfahan, central Iran. Iran J. Public Health 2016, 45, 650–656. [Google Scholar] [PubMed]
- Heidarieh, P.; Hashemi Shahraki, A.; Yaghoubfar, R.; Hajehasani, A.; Mirsaeidi, M. Microbiological Analysis of Hemodialysis Water in a Developing Country. ASAIO J. 2016, 62, 332–339. [Google Scholar] [CrossRef]
- Novosad, S.A.; Lake, J.; Nguyen, D.; Soda, E.; Moulton-Meissner, H.; Pho, M.T.; Gualandi, N.; Bepo, L.; Stanton, R.A.; Daniels, J.B.; et al. Multicenter Outbreak of Gram-Negative Bloodstream Infections in Hemodialysis Patients. Am. J. Kidney Dis. 2019, 74, 610–619. [Google Scholar] [CrossRef]
- Otaíza, F. Informe de Vigilancia de Infecciones Asociadas a la Atención en Salud. In Ministerio de Salud, Departamento de Calidad y Formación Control de IAAS; Minsal: Santiago, Chile, 2017. Available online: www.minsal.cl/wp-content/uploads/2015/09/informe-vigilancia-2017.pdf (accessed on 15 July 2020).
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa Biofilm, a Programmed Bacterial Life for Fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef]
- Trifonova, A.; Strateva, T. Stenotrophomonas maltophilia—A low-grade pathogen with numerous virulence factors. Infect. Dis. 2019, 51, 168–178. [Google Scholar] [CrossRef]
- Fang, Q.; Feng, Y.; Feng, P.; Wang, X.; Zong, Z. Nosocomial bloodstream infection and the emerging carbapenem-resistant pathogen Ralstonia insidiosa. BMC Infect. Dis. 2019, 19, 334. [Google Scholar] [CrossRef]
- Kendirli, T.; Ciftci, E.; Ince, E.; Incesoy, S.; Guriz, H.; Aysev, A.D.; Tutar, E.; Yavuz, G.; Dogru, U. Ralstonia pickettii outbreak associated with contaminated distilled water used for respiratory care in a paediatric intensive care unit. J. Hosp. Infect. 2004, 56, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Tejera, D.; Limongi, G.; Bertullo, M.; Cancela, M. Ralstonia pickettii bacteremia in hemodialysis patients: A report of two cases. Rev. Bras. Ter. Intensiva 2016, 28, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Bridger, N.; Drews, S.; Burdz, T.; Wiebe, D.; Pacheco, A.L.; Ng, B.; Bernard, K. Isolation and characterization of Pigmentiphaga-like isolates from human clinical material. J. Med. Microbiol. 2013, 62, 708–711. [Google Scholar] [CrossRef] [PubMed]
- Belila, A.; El-Chakhtoura, J.; Otaibia, M.; Muyzerc, G.; Gonzalez-Gila, G.; Saikalya, P.E.; van Loosdrechtb, M.C.M.; Vrouwenvelde, J.S. Bacterial community structure and variation in a full-scale seawater desalination plant for drinking water production. Water Res. 2016, 94, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Chanasit, W.; Gonzaga, Z.J.C.; Rehm, B.H.A. Analysis of the alginate O-acetylation machinery in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2020, 104, 2179–2191. [Google Scholar] [CrossRef] [PubMed]
- Loo, L.W.; Liew, Y.X.; Choong, H.L.; Tan, A.L.; Chlebicki, P. Microbiology and audit of vascular access-associated bloodstream infections in multi-ethnic Asian hemodialysis patients in a tertiary hospital. Infect. Dis. 2015, 47, 225–230. [Google Scholar] [CrossRef]
- Karanika, S.; Zervou, F.N.; Zacharioudakis, I.M.; Paudel, S.; Mylonakis, E. Risk factors for meticillin-resistant Staphylococcus aureus colonization in dialysis patients: A meta-analysis. J. Hosp. Infect. 2015, 91, 257–263. [Google Scholar] [CrossRef]
- Imaizumi, T.; Hasegawa, T.; Nomura, A.; Sasaki, S.; Nishiwaki, H.; Ozeki, T.; Shimizu, H.; Minatoguchi, S.; Yamakawa, T.; Yazawa, M.; et al. Association between staphylococcus aureus bacteremia and hospital mortality in hemodialysis patients with bloodstream infection: A multicenter cohort from Japanese tertiary care centers. Ther. Apher. Dial. 2017, 21, 354–360. [Google Scholar] [CrossRef]
- Gil, C.; Solano, C.; Burgui, S.; Latasa, C.; Garcia, B.; Toledo-Arana, A.; Lasa, I.; Valle, J. Biofilm matrix exoproteins induce a protective immune response against Staphylococcus aureus biofilm infection. Infect. Immun. 2014, 82, 1017–1029. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Beta-Lactams and beta-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6. [Google Scholar] [CrossRef]
- Vasoo, S.; Barreto, J.N.; Tosh, P.K. Emerging Issues in Gram-negative Bacterial Resistance: An Update for the Practicing Clinician. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2015; Volume 90, pp. 395–403. [Google Scholar] [CrossRef]
- Doi, Y.; Wachino, J.I.; Arakawa, Y. Aminoglycoside resistance: The emergence of acquired 16S ribosomal RNA methyltransferases. Infect. Dis. Clin. 2016, 30, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.; Brooks, B.D.; Brooks, A.E. The Complex Relationship between Virulence and Antibiotic Resistance. Genes 2017, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Nagy, S.S.; Kjelleberg, S.; Gillings, M.R.; Paulsen, I.T. Rapid microevolution of biofilm cells in response to antibiotics. NPJ Biofilms Microbiomes 2019, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Allison, K.R.; Brynildsen, M.P.; Collins, J.J. Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature 2011, 473, 216–220. [Google Scholar] [CrossRef]
- Jung, S.H.; Ryu, C.M.; Kim, J.S. Bacterial persistence: Fundamentals and clinical importance. J. Microbiol. 2019, 57, 829–835. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
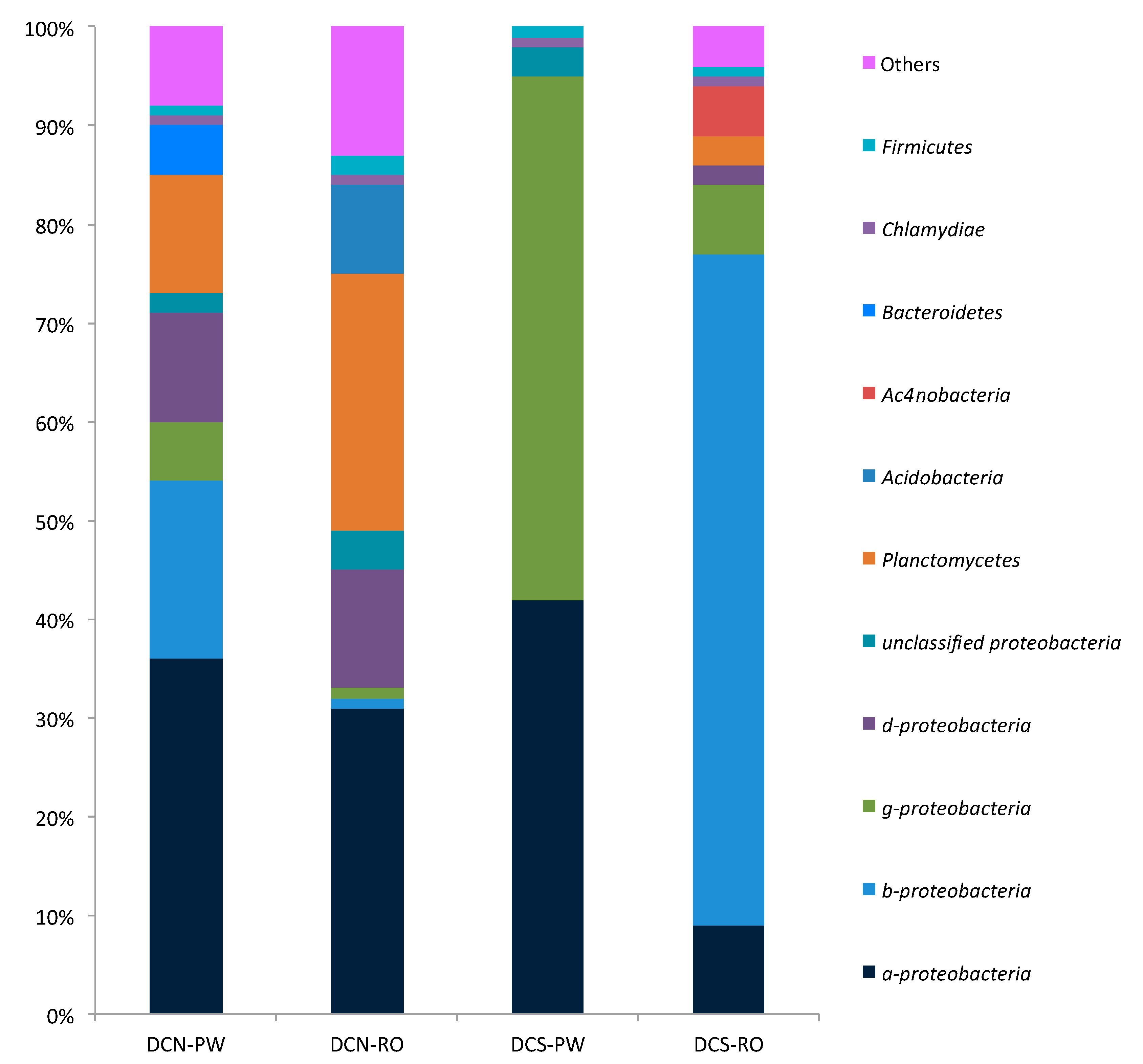{kind=link}
| Strain | Closest Relative Sequence | Phylum | Identity (%) | Gen Bank Access Number |
|---|---|---|---|---|
| DCN RO-3 | Pseudomonas sp. | Proteobacteria | 98 | MH114031.1 |
| DCN RO-5 | Pseudomonas veronii | Proteobacteria | 95 | MH482963.1 |
| DCN RO-6 | Stenotrophomonas maltophilia | Proteobacteria | 96 | FN395264.1 |
| DCN RO-7 | Agrobacterium tumefaciens | Proteobacteria | 98 | MG575921.1 |
| DCN RO-12 | Pigmentiphaga kullae | Proteobacteria | 98 | KR856352.1 |
| DCN PW-4 | Ralstonia sp. | Proteobacteria | 97 | MH844635.1 |
| DCN PW-10 | Arthrobacter sp. | Actinobacteria | 95 | JQ316230.1 |
| DCN OW-7 | Bacteroides dorei | Bacteroidetes | 96 | CP008741.1 |
| DCS RO-1 | Pseudomonas sp. | Proteobacteria | 98 | MK883238.1 |
| DCS RO9 | Staphylococcus sp. | Firmicutes | 97 | LC435711.1 |
| DCS RO-11 | Staphylococcus pasteuri | Firmicutes | 98 | MG996881.1 |
| DCS RO-17 | Pseudomonas fluorescens | Proteobacteria | 98 | EU434450.1 |
| DCS PW-7 | Pseudomonas poae | Proteobacteria | 97 | MG835964.1 |
| DCS OW-3 | Stenotrophomonas sp. | Proteobacteria | 97 | KP663381.1 |
| DCS OW-7 | Stenotrophomonas sp. | Proteobacteria | 98 | KY357351.1 |
| Antibiotics | % Resistant Strains |
|---|---|
| Ampicillin | 69% |
| Chloramphenicol | 55% |
| Tetracycline | 24% |
| Cefotaxime | 53% |
| Ciprofloxacin | 15% |
| Trimethoprim/sulfamethoxazole | 38% |
| Gentamicin | 31% |
| Bacterial Genus | AM 10 | CHL 30 | Te 30 | CTX 30 | CIP 5 | TSX 25 | GE 10 |
|---|---|---|---|---|---|---|---|
| Pseudomonas | 67% (7/11) | 55% (6/11) | 27% (3/11) | 67% (7/11) | 18% (2/11) | 36% (4/11) | 18% (2/11) |
| Stenotrophomonas | 100% (4/4) | 50% (2/4) | 50% (2/4) | 50% (2/4) | 0% (0/4) | 25% (1/4) | 50% (2/4) |
| Agrobacterium | 100% (0/2) | 0% (0/2) | 0% (0/2) | 100% (2/2) | 0% (0/2) | 0% (0/2) | 50% (1/2) |
| Pigmentiphaga | 100% (2/2) | 0% (0/2) | 50% (1/2) | 100% (2/2) | 0% (0/2) | 0% (0/2) | 100% (2/2) |
| Ralstonia | 100% (2/2) | 50% (1/2) | 0% (0/2) | 0% (0/2) | 0% (0/2) | 50% (1/2) | 0% (0/2) |
| Archobacter | 0% (0/1) | 0% (0/1) | 0% (0/1) | 0% (0/1) | 0% (0/1) | 0% (0/1) | 0% (0/1) |
| Bacteroides | 50% (1/2) | 0% (0/2) | 0% (0/2) | 0% (0/2) | 0% (0/2) | 0% (0/2) | 50% (1/2) |
| Staphylococcus | 80% (4/5) | 40% (2/5) | 20% (1/5) | 40% (2/5) | 0% (0/5) | 100% (5/5) | 20% (1/5) |
| * E. coli ATCC 25922 | R | S | S | S | S | S | S |
| * P. aeruginosa ATCC 27853 | R | R | R | R | S | R | S |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuevas, J.-P.; Moraga, R.; Sánchez-Alonzo, K.; Valenzuela, C.; Aguayo, P.; Smith, C.T.; García, A.; Fernandez, Í.; Campos, V.L. Characterization of the Bacterial Biofilm Communities Present in Reverse-Osmosis Water Systems for Haemodialysis. Microorganisms 2020, 8, 1418. https://doi.org/10.3390/microorganisms8091418
Cuevas J-P, Moraga R, Sánchez-Alonzo K, Valenzuela C, Aguayo P, Smith CT, García A, Fernandez Í, Campos VL. Characterization of the Bacterial Biofilm Communities Present in Reverse-Osmosis Water Systems for Haemodialysis. Microorganisms. 2020; 8(9):1418. https://doi.org/10.3390/microorganisms8091418
Chicago/Turabian StyleCuevas, Juan-Pablo, Ruben Moraga, Kimberly Sánchez-Alonzo, Cristian Valenzuela, Paulina Aguayo, Carlos T. Smith, Apolinaria García, Ítalo Fernandez, and Víctor L Campos. 2020. "Characterization of the Bacterial Biofilm Communities Present in Reverse-Osmosis Water Systems for Haemodialysis" Microorganisms 8, no. 9: 1418. https://doi.org/10.3390/microorganisms8091418
APA StyleCuevas, J.-P., Moraga, R., Sánchez-Alonzo, K., Valenzuela, C., Aguayo, P., Smith, C. T., García, A., Fernandez, Í., & Campos, V. L. (2020). Characterization of the Bacterial Biofilm Communities Present in Reverse-Osmosis Water Systems for Haemodialysis. Microorganisms, 8(9), 1418. https://doi.org/10.3390/microorganisms8091418