The Anti-Pseudomonal Peptide D-BMAP18 Is Active in Cystic Fibrosis Sputum and Displays Anti-Inflammatory In Vitro Activity

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Peptides and Antibiotics
2.2. Bacterial Strains and Culture Conditions
2.3. Killing Activity of D-BMAP18 in Cystic-Fibrosis Sputum
2.4. Inhibition of Biofilm Deposition and Eradication
2.5. Cytotoxicity Tests Against Human Cell Lines
2.6. Anti-Inflammatory Property of D-BMP18 on THP-1 Cells
2.7. Isolation and Differentiation of Human Peripheral Monocytes into Macrophages and Their In Vitro Polarization
2.8. Anti-Inflammatory Property of D-BMAP18 on Macrophages Differentiated Cells
2.9. Statistical Analysis
3. Results
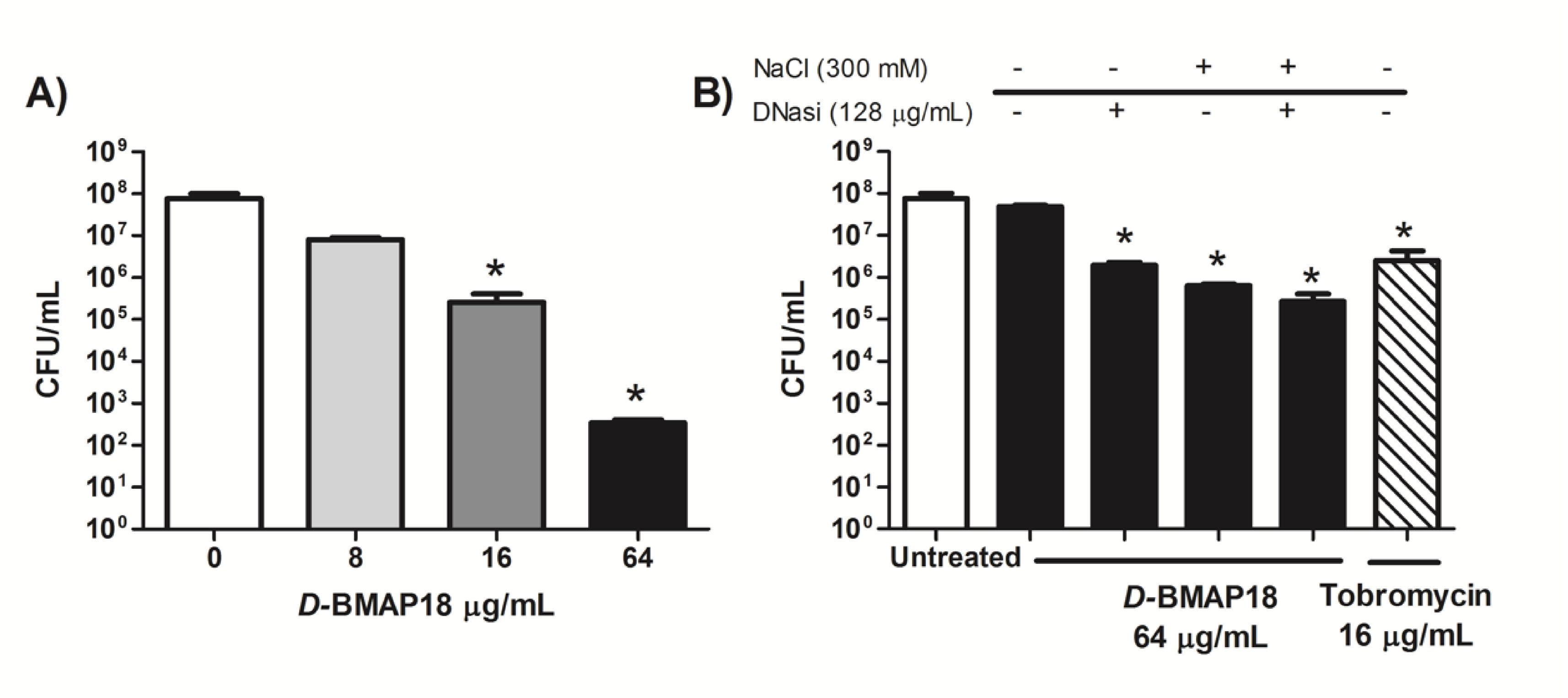
3.1. Antipseudomonal Activity of D-BMAP18 in CF Sputum
3.2. Activity of D-BMAP18 Against Biofilm Deposition
3.3. Activity of D-BMAP18 Against Preformed Biofilms
3.4. In vitro Cytotoxicity of D-BMAP18 Against Human Cell Lines
3.5. D-BMAP18 Reduces the Bacterial Pro-Inflammatory Stimulus In Vitro
3.6. The Overall Anti-Inflammatory and Anti-Fibrotic Activity of D-BMAP18
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bruscia, E.M.; Bonfield, T.L. Cystic Fibrosis Lung Immunity: The Role of the Macrophage. J. Innate Immun. 2016, 8, 550–563. [Google Scholar] [CrossRef]
- Heijerman, H. Infection and inflammation in cystic fibrosis: A short review. J. Cyst. Fibros. 2005, 4, 3–5. [Google Scholar] [CrossRef]
- Martin, C.; Hamard, C.; Kanaan, R.; Boussaud, V.; Grenet, D.; Abély, M.; Hubert, D.; Munck, A.; Lemonnier, S.L.; Burgel, P.-R. Causes of death in French cystic fibrosis patients: The need for improvement in transplantation referral strategies! J. Cyst. Fibros. 2016, 15, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Zolin, A.; Bossi, A.; Cirilli, N.; Kashirskaya, N.; Padoan, R. Cystic Fibrosis Mortality in Childhood. Data from European Cystic Fibrosis Society Patient Registry. Int. J. Environ. Res. Public Heal. 2018, 15, 2020. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Dasenbrook, E.C.; Checkley, W.; Merlo, C.A.; Konstan, M.W.; Lechtzin, N.; Boyle, M. Association Between Respiratory Tract Methicillin-Resistant Staphylococcus aureus and Survival in Cystic Fibrosis. JAMA 2010, 303, 2386. [Google Scholar] [CrossRef]
- Flume, P.; Mogayzel, P.J.; Robinson, K.A.; Goss, C.; Rosenblatt, R.L.; Kuhn, R.J.; Marshall, B.C. Clinical Practice Guidelines for Pulmonary Therapies Committee* Cystic Fibrosis Pulmonary Guidelines. Am. J. Respir. Crit. Care Med. 2009, 180, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Burrell, A.; Fanous, H.; Chaney, H.; Sami, I.; Perez, G.F.; Koumbourlis, A.C.; Freishtat, R.J.; Crandall, K.A. Antibiotic multidrug resistance in the cystic fibrosis airway microbiome is associated with decreased diversity. Heliyon 2018, 4, e00795. [Google Scholar] [CrossRef]
- Sousa, A.M.; Pereira, M.O. Pseudomonas aeruginosa Diversification during Infection Development in Cystic Fibrosis Lungs—A Review. Pathogens 2014, 3, 680–703. [Google Scholar] [CrossRef]
- Geitani, R.; Moubareck, C.A.; Touqui, L.; Sarkis, D.K. Cationic antimicrobial peptides: Alternatives and/or adjuvants to antibiotics active against methicillin-resistant Staphylococcus aureus and multidrug-resistant Pseudomonas aeruginosa. BMC Microbiol. 2019, 19, 54. [Google Scholar] [CrossRef]
- Lee, J.-K.; Luchian, T.; Park, Y. New antimicrobial peptide kills drug-resistant pathogens without detectable resistance. Oncotarget 2018, 9, 15616–15634. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-J.; Gallo, R.L. Antimicrobial peptides. Curr. Boil. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Wadman, M.W.; Dohm, M.T.; Czyzewski, A.M.; Spormann, A.M.; Barron, A.E. Antimicrobial Peptoids Are Effective against Pseudomonas aeruginosa Biofilms. Antimicrob. Agents Chemother. 2011, 55, 3054–3057. [Google Scholar] [CrossRef] [PubMed]
- Saiman, L.; Tabibi, S.; Starner, T.D.; Gabriel, P.S.; Winokur, P.L.; Jia, H.P.; McCray, P.B.; Tack, B.F. Cathelicidin Peptides Inhibit Multiply Antibiotic-Resistant Pathogens from Patients with Cystic Fibrosis. Antimicrob. Agents Chemother. 2001, 45, 2838–2844. [Google Scholar] [CrossRef]
- Zhang, L.; Parente, J.; Harris, S.M.; Woods, D.E.; Hancock, R.E.W.; Falla, T.J. Antimicrobial Peptide Therapeutics for Cystic Fibrosis. Antimicrob. Agents Chemother. 2005, 49, 2921–2927. [Google Scholar] [CrossRef]
- Benincasa, M.; Skerlavaj, B.; Gennaro, R.; Pellegrini, A.; Zanetti, M. In vitro and in vivo antimicrobial activity of two α-helical cathelicidin peptides and of their synthetic analogs. Pept. 2003, 24, 1723–1731. [Google Scholar] [CrossRef]
- Mardirossian, M.; Pompilio, A.; Crocetta, V.; De Nicola, S.; Guida, F.; Degasperi, M.; Gennaro, R.; Di Bonaventura, G.; Scocchi, M. In vitro and in vivo evaluation of BMAP-derived peptides for the treatment of cystic fibrosis-related pulmonary infections. Amino Acids 2016, 48, 2253–2260. [Google Scholar] [CrossRef]
- Pompilio, A.; Crocetta, V.; Scocchi, M.; Pomponio, S.; Di Vincenzo, V.; Mardirossian, M.; Gherardi, G.; Fiscarelli, E.V.; Dicuonzo, G.; Gennaro, R.; et al. Potential novel therapeutic strategies in cystic fibrosis: Antimicrobial and anti-biofilm activity of natural and designed α-helical peptides against Staphylococcus aureus, Pseudomonas aeruginosa, and Stenotrophomonas maltophilia. BMC Microbiol. 2012, 12, 145. [Google Scholar] [CrossRef]
- Pompilio, A.; Scocchi, M.; Pomponio, S.; Guida, F.; Di Primio, A.; Fiscarelli, E.V.; Gennaro, R.; Di Bonaventura, G. Antibacterial and anti-biofilm effects of cathelicidin peptides against pathogens isolated from cystic fibrosis patients. Pept. 2011, 32, 1807–1814. [Google Scholar] [CrossRef]
- Mardirossian, M.; Pompilio, A.; Degasperi, M.; Runti, G.; Pacor, S.; Di Bonaventura, G.; Scocchi, M. D-BMAP18 Antimicrobial Peptide Is Active In vitro, Resists to Pulmonary Proteases but Loses Its Activity in a Murine Model of Pseudomonas aeruginosa Lung Infection. Front. Chem. 2017, 5, 40. [Google Scholar] [CrossRef]
- Palmer, K.L.; Aye, L.M.; Whiteley, M. Nutritional Cues Control Pseudomonas aeruginosa Multicellular Behavior in Cystic Fibrosis Sputum. J. Bacteriol. 2007, 189, 8079–8087. [Google Scholar] [CrossRef] [PubMed]
- Masat, E.; Gasparini, C.; Agostinis, C.; Bossi, F.; Radillo, O.; De Seta, F.; Tamassia, N.; Cassatella, M.A.; Bulla, R. RelB activation in anti-inflammatory decidual endothelial cells: A master plan to avoid pregnancy failure? Sci. Rep. 2015, 5, 14847. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Pfaffl, M.W.; Zhao, S.; Spiess, A.N.; Boggy, G.; Blom, J.; Rutledge, R.G.; Sisti, D.; Lievens, A.; De Preter, K.; et al. Evaluation of qPCR curve analysis methods for reliable biomarker discovery: Bias, resolution, precision, and implications. Methods 2013, 59, 32–46. [Google Scholar] [CrossRef]
- Namas, R.A.; AlMahmoud, K.; Mi, Q.; Ghuma, A.; Namas, R.; Zaaqoq, A.; Zhu, X.; Abdul-Malak, O.; Sperry, J.; Zamora, R.; et al. Individual-specific principal component analysis of circulating inflammatory mediators predicts early organ dysfunction in trauma patients. J. Crit. Care 2016, 36, 146–153. [Google Scholar] [CrossRef]
- Klodzinska, S.N.; Priemel, P.A.; Rades, T.; Nielsen, H.M. Inhalable Antimicrobials for Treatment of Bacterial Biofilm-Associated Sinusitis in Cystic Fibrosis Patients: Challenges and Drug Delivery Approaches. Int. J. Mol. Sci. 2016, 17, 1688. [Google Scholar] [CrossRef] [PubMed]
- Leeuwen, P.B.V.D.W.-V.; Arets, B.; Van Der Ent, C.K.; Beekman, J.M. Infection, inflammation and exercise in cystic fibrosis. Respir. Res. 2013, 14, 32. [Google Scholar] [CrossRef]
- Mulcahy, L.R.; Isabella, V.M.; Lewis, K. Pseudomonas aeruginosa Biofilms in Disease. Microb. Ecol. 2013, 68, 1–12. [Google Scholar] [CrossRef]
- Dosler, S.; Karaaslan, E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides 2014, 62, 32–37. [Google Scholar] [CrossRef]
- Galdiero, E.; Lombardi, L.; Falanga, A.; Libralato, G.; Guida, M.; Carotenuto, R. Biofilms: Novel Strategies Based on Antimicrobial Peptides. Pharmaceutics 2019, 11, 322. [Google Scholar] [CrossRef]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.W.; Rehm, B.H.A.; Hancock, R.E.W. Human Host Defense Peptide LL-37 Prevents Bacterial Biofilm Formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef]
- Wozniak, D.J.; Keyser, R. Effects of subinhibitory concentrations of macrolide antibiotics on Pseudomonas aeruginosa. Chest 2004, 125, 62S–69S. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Spiegelman, G.B.; Yim, G. The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 2006, 9, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Elborn, J.S.; Vataire, A.-L.; Fukushima, A.; Aballéa, S.; Khemiri, A.; Moore, C.; Medic, G.; Hemels, M.E. Comparison of Inhaled Antibiotics for the Treatment of Chronic Pseudomonas aeruginosa Lung Infection in Patients With Cystic Fibrosis: Systematic Literature Review and Network Meta-analysis. Clin. Ther. 2016, 38, 2204–2226. [Google Scholar] [CrossRef]
- Rosenecker, J.; Naundorf, S.; Gersting, S.W.; Hauck, R.W.; Gessner, A.; Nicklaus, P.; Muller, R.H.; Rudolph, C. Interaction of bronchoalveolar lavage fluid with polyplexes and lipoplexes: Analysing the role of proteins and glycoproteins. J. Gene Med. 2002, 5, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Manzenreiter, R.; Kienberger, F.; Marcos, V.; Schilcher, K.; Krautgartner, W.D.; Obermayer, A.; Huml, M.; Stoiber, W.; Hector, A.; Griese, M.; et al. Ultrastructural characterization of cystic fibrosis sputum using atomic force and scanning electron microscopy. J. Cyst. Fibros. 2012, 11, 84–92. [Google Scholar] [CrossRef]
- Ratjen, F.; Paul, K.; Van Koningsbruggen, S.; Breitenstein, S.; Rietschel, E.; Nikolaizik, W. DNA concentrations in BAL fluid of cystic fibrosis patients with early lung disease: Influence of treatment with dornase alpha. Pediatr. Pulmonol. 2004, 39, 1–4. [Google Scholar] [CrossRef]
- Aoki, W.; Ueda, M. Characterization of Antimicrobial Peptides toward the Development of Novel Antibiotics. Pharmaceuticals 2013, 6, 1055–1081. [Google Scholar] [CrossRef]
- Huang, L.C.; Petkova, T.D.; Reins, R.Y.; Proske, R.J.; McDermott, A.M. Multifunctional Roles of Human Cathelicidin (LL-37) at the Ocular Surface. Investig. Opthalmology Vis. Sci. 2006, 47, 2369–2380. [Google Scholar] [CrossRef]
- Reeves, E.P.; Molloy, K.; Pohl, K.; McElvaney, N.G. Hypertonic Saline in Treatment of Pulmonary Disease in Cystic Fibrosis. Sci. World J. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Dong, W.; Mao, X.; Guan, Y.; Kang, Y.; Shang, D. Antimicrobial and anti-inflammatory activities of three chensinin-1 peptides containing mutation of glycine and histidine residues. Sci. Rep. 2017, 7, 40228. [Google Scholar] [CrossRef]
- Popovic, S.; Urbán, E.; Lukic, M.L.; Conlon, J.M. Peptides with antimicrobial and anti-inflammatory activities that have therapeutic potential for treatment of acne vulgaris. Pept 2012, 34, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Arribillaga, L.; Dotor, J.; Basagoiti, M.; Riezu-Boj, J.I.; Borrás-Cuesta, F.; Lasarte, J.J.; Sarobe, P.; Cornet, M.E.; Feijoo, E. Therapeutic effect of a peptide inhibitor of TGF-β on pulmonary fibrosis. Cytokine 2011, 53, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Kramer, E.L.; Clancy, J.P. TGFβ as a therapeutic target in cystic fibrosis. Expert Opin. Ther. Targets 2017, 22, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.E.; Eickelberg, O. The Impact of TGF-? on Lung Fibrosis. Proc. Am. Thorac. Soc. 2012, 9, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Eckert, R. Road to clinical efficacy: Challenges and novel strategies for antimicrobial peptide development. Futur. Microbiol. 2011, 6, 635–651. [Google Scholar] [CrossRef]
- Barańska-Rybak, W.; Pikuła, M.; Dawgul, M.; Kamysz, W.; Trzonkowski, P.; Roszkiewicz, J. Safety profile of antimicrobial peptides: Camel, citropin, protegrin, temporin a and lipopeptide on HaCaT keratinocytes. Acta Pol. Pharm. Drug Res. 2013, 70, 795–801. [Google Scholar]
- Paiva, A.D.; De Oliveira, M.D.; De Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiol. 2012, 158, 2851–2858. [Google Scholar] [CrossRef]
- Brender, J.R.; McHenry, A.J.; Ramamoorthy, A. Does Cholesterol Play a Role in the Bacterial Selectivity of Antimicrobial Peptides? Front. Immunol. 2012, 3, 3. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Tm | Sense | Sequence | Accession Number |
|---|---|---|---|---|
| 18S | 60 | Forward | ATCCCTGAAAAGTTCCAGCA | NM_022551.2 |
| Reverse | CCCTCTTGGTGAGGTCAATG | |||
| TNF-α | 66 | Forward | GGCCCAGGCAGTCAGATCAT | NM_000594.3 |
| Reverse | GGGGCTCTTGATGGCAGAGA | |||
| IL-1β | 60 | Forward | GTACATCCTCGACGGCATC | NM_000600.3 |
| Reverse | CCAGGCAAGTCTCCTCATTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degasperi, M.; Agostinis, C.; Mardirossian, M.; Maschio, M.; Taddio, A.; Bulla, R.; Scocchi, M. The Anti-Pseudomonal Peptide D-BMAP18 Is Active in Cystic Fibrosis Sputum and Displays Anti-Inflammatory In Vitro Activity. Microorganisms 2020, 8, 1407. https://doi.org/10.3390/microorganisms8091407
Degasperi M, Agostinis C, Mardirossian M, Maschio M, Taddio A, Bulla R, Scocchi M. The Anti-Pseudomonal Peptide D-BMAP18 Is Active in Cystic Fibrosis Sputum and Displays Anti-Inflammatory In Vitro Activity. Microorganisms. 2020; 8(9):1407. https://doi.org/10.3390/microorganisms8091407
Chicago/Turabian StyleDegasperi, Margherita, Chiara Agostinis, Mario Mardirossian, Massimo Maschio, Andrea Taddio, Roberta Bulla, and Marco Scocchi. 2020. "The Anti-Pseudomonal Peptide D-BMAP18 Is Active in Cystic Fibrosis Sputum and Displays Anti-Inflammatory In Vitro Activity" Microorganisms 8, no. 9: 1407. https://doi.org/10.3390/microorganisms8091407
APA StyleDegasperi, M., Agostinis, C., Mardirossian, M., Maschio, M., Taddio, A., Bulla, R., & Scocchi, M. (2020). The Anti-Pseudomonal Peptide D-BMAP18 Is Active in Cystic Fibrosis Sputum and Displays Anti-Inflammatory In Vitro Activity. Microorganisms, 8(9), 1407. https://doi.org/10.3390/microorganisms8091407