Faecal Microbiota of Dogs Offered a Vegetarian Diet with or without the Supplementation of Feather Meal and either Cornmeal, Rye or Fermented Rye: A Preliminary Study

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Diets
2.3. 16S rRNA Gene Analyses
2.4. Statistical Analyses
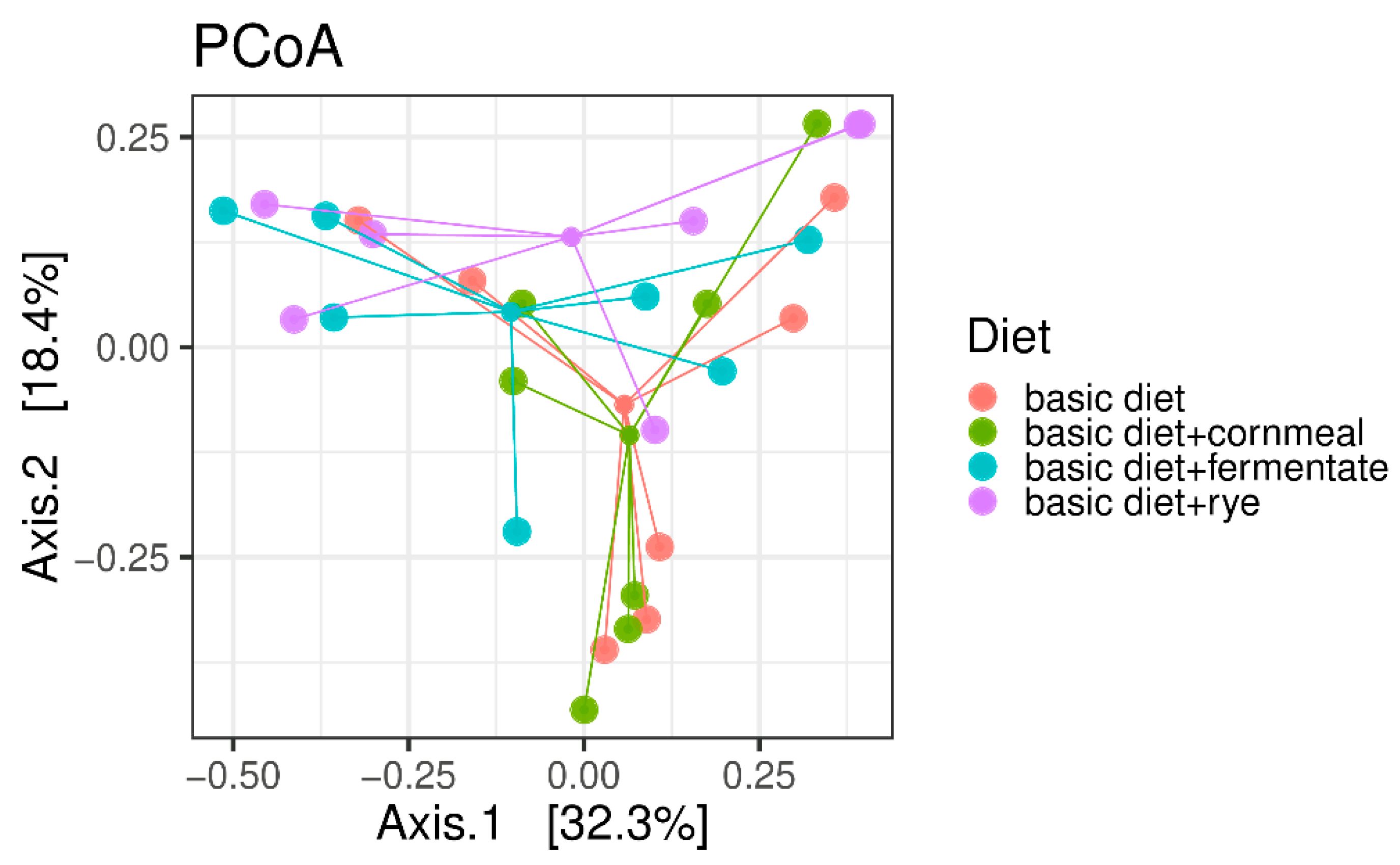
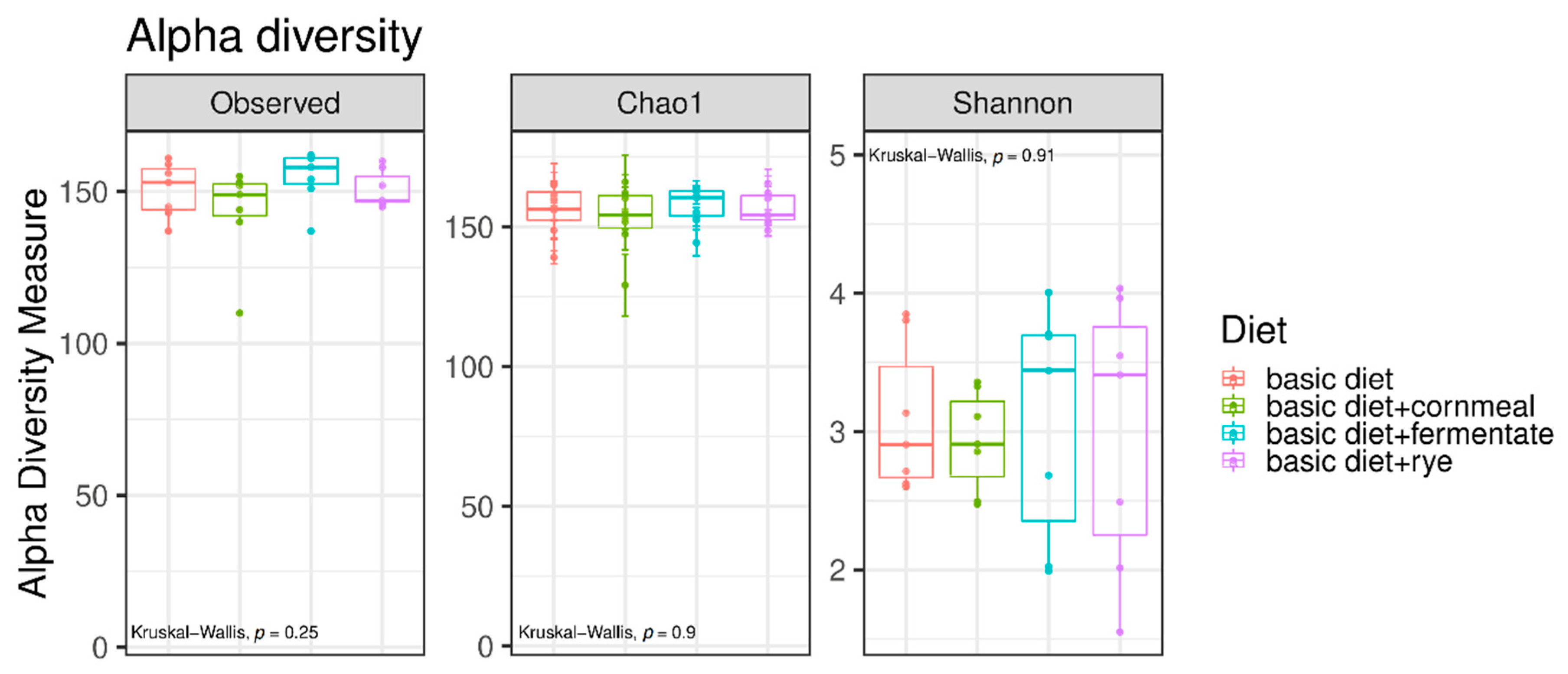
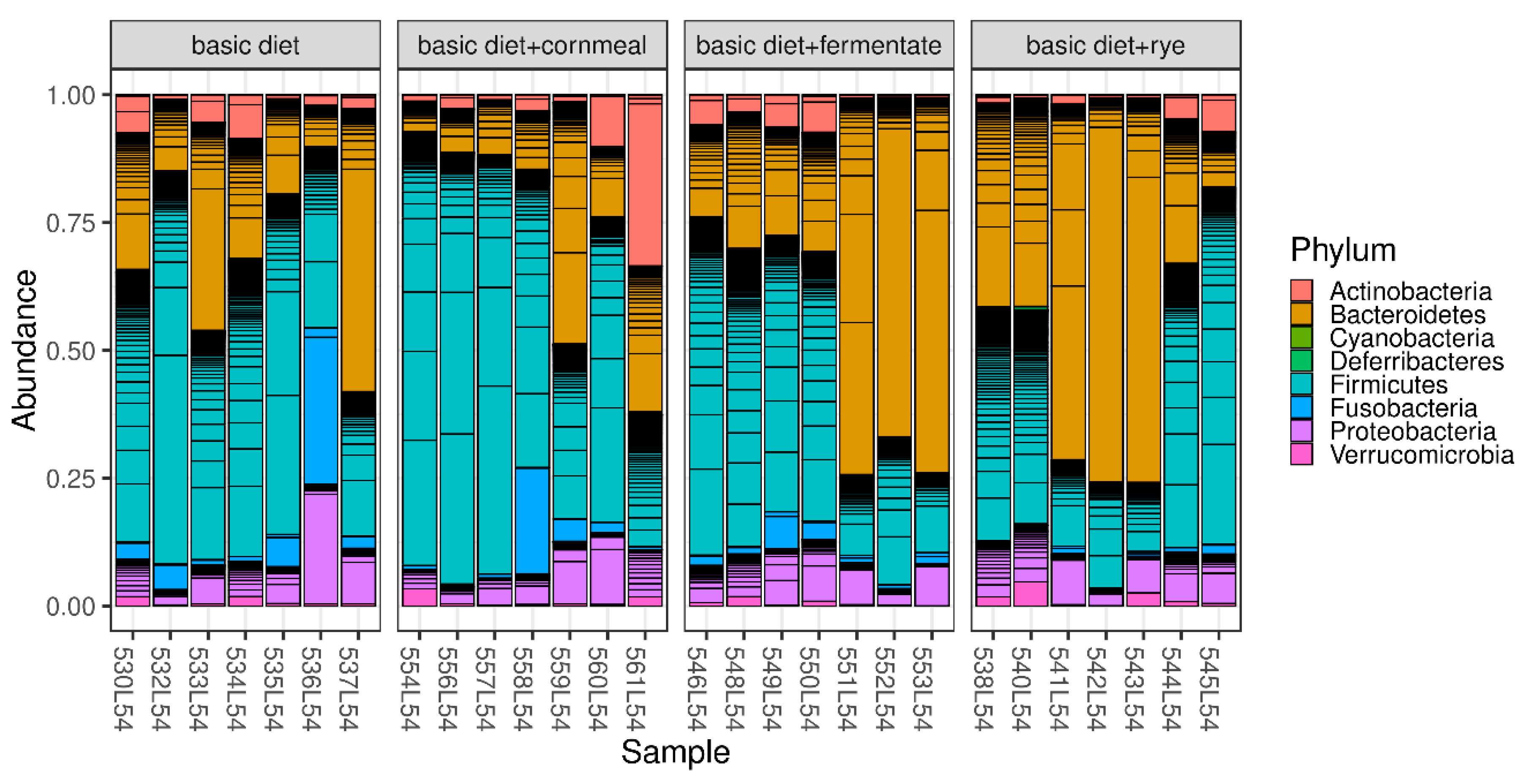
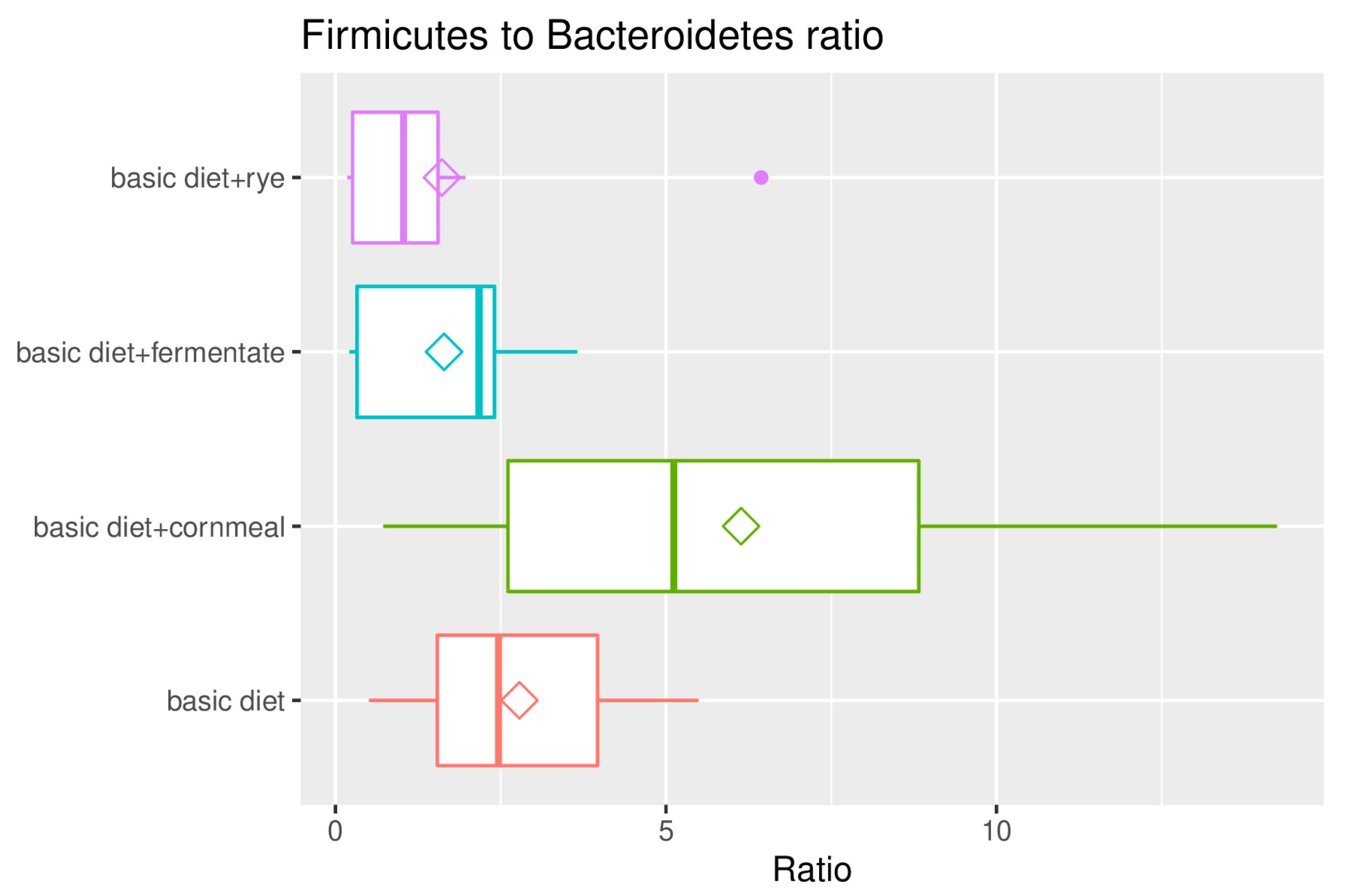
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alessandri, G.; Milani, C.; Mancabelli, L.; Mangifesta, M.; Lugli, G.A.; Viappiani, A.; Duranti, S.; Turroni, F.; Ossiprandi, M.C.; van Sinderen, D. Metagenomic dissection of the canine gut microbiota: Insights into taxonomic, metabolic and nutritional features. Environ. Microbiol. 2019, 21, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Bazolli, R.; Vasconcellos, R.; De-Oliveira, L.; Sá, F.; Pereira, G.; Carciofi, A. Effect of the particle size of maize, rice, and sorghum in extruded diets for dogs on starch gelatinization, digestibility, and the fecal concentration of fermentation products. J. Anim. Sci. 2015, 93, 2956–2966. [Google Scholar] [CrossRef] [PubMed]
- Carciofi, A.; Sakomura, N.; Kawauchi, I.; Vasconcellos, R. Digestibility and metabolizable energy of some carbohydrate sources for dogs. Anim. Feed Sci. Technol. 2010, 156, 121–125. [Google Scholar]
- Buff, P.; Carter, R.; Bauer, J.; Kersey, J. Natural pet food: A review of natural diets and their impact on canine and feline physiology. J. Anim. Sci. 2014, 92, 3781–3791. [Google Scholar] [CrossRef]
- Dodd, S.A.; Cave, N.J.; Adolphe, J.L.; Shoveller, A.K.; Verbrugghe, A. Plant-based (vegan) diets for pets: A survey of pet owner attitudes and feeding practices. PLoS ONE 2019, 14, e0210806. [Google Scholar] [CrossRef]
- Dodd, S.; Cave, N.; Abood, S.; Shoveller, A.-K.; Adolphe, J.; Verbrugghe, A. An observational study of pet feeding practices and how these have changed between 2008 and 2018. Vet. Rec. 2020, 186, 643. [Google Scholar] [CrossRef]
- Freeman, L.M.; Chandler, M.L.; Hamper, B.A.; Weeth, L.P. Current knowledge about the risks and benefits of raw meat–based diets for dogs and cats. J. Am. Vet. Med. Assoc. 2013, 243, 1549–1558. [Google Scholar] [CrossRef]
- Zimmer, J.; Lange, B.; Frick, J.-S.; Sauer, H.; Zimmermann, K.; Schwiertz, A.; Rusch, K.; Klosterhalfen, S.; Enck, P. A vegan or vegetarian diet substantially alters the human colonic faecal microbiota. Eur. J. Clin. Nutr. 2012, 66, 53–60. [Google Scholar] [CrossRef]
- Losasso, C.; Eckert, E.M.; Mastrorilli, E.; Villiger, J.; Mancin, M.; Patuzzi, I.; Di Cesare, A.; Cibin, V.; Barrucci, F.; Pernthaler, J. Assessing the influence of vegan, vegetarian and omnivore oriented Westernized dietary styles on human gut microbiota: A cross sectional study. Front. Microbiol. 2018, 9, 317. [Google Scholar] [CrossRef]
- Kim, M.S.; Hwang, S.S.; Park, E.J.; Bae, J.W. Strict vegetarian diet improves the risk factors associated with metabolic diseases by modulating gut microbiota and reducing intestinal inflammation. Environ. Microbiol. Rep. 2013, 5, 765–775. [Google Scholar] [CrossRef]
- Zentek, J.; Marquart, B.; Pietrzak, T.; Ballevre, O.; Rochat, F. Dietary effects on bifidobacteria and Clostridium perfringens in the canine intestinal tract. J. Anim. Physiol. Anim. Nutr. 2003, 87, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Nat. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Brown, W. Nutritional and ethical issues regarding vegetarianism in the domestic dog. Recent Adv. Anim. Nutr. Aust. 2009, 17, 137–143. [Google Scholar]
- Brown, R.G. Protein in dog food. Can. Vet. J. 1989, 30, 528. [Google Scholar] [PubMed]
- Meeker, D.; Meisinger, J. Rendered ingredients significantly influence sustainability, quality, and safety of pet food. J. Anim. Sci. 2015, 93, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, I.; Pezzato, L.E.; Barros, M.M. Amino acid availability and protein digestibility of several protein sources for Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2008, 14, 396–404. [Google Scholar] [CrossRef]
- Zeiger, A.L. Untersuchungen zum Einsatz und Futterwert asche- und protein- bzw. keratinreicher Nebenprodukte der Geflügelschlachtung in der Fütterung von Hunden. Ph.D. Thesis, University of Veterinary Medicine Hanover, Foundation, Hannover, Germany, 20 November 2015. [Google Scholar]
- Goñi, I.; Garcia-Diz, L.; Mañas, E.; Saura-Calixto, F. Analysis of resistant starch: A method for foods and food products. Food Chem. 1996, 56, 445–449. [Google Scholar] [CrossRef]
- Mann, J.; Cummings, J. Possible implications for health of the different definitions of dietary fibre. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 226–229. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Nordlund, E.; Aura, A.-M.; Mattila, I.; Kössö, T.; Rouau, X.; Poutanen, K. Formation of phenolic microbial metabolites and short-chain fatty acids from rye, wheat, and oat bran and their fractions in the metabolical in vitro colon model. J. Agric. Food Chem. 2012, 60, 8134–8145. [Google Scholar] [CrossRef]
- Bunte, S.; Grone, R.; Keller, B.; Keller, C.; Galvez, E.; Strowig, T.; Kamphues, J.; Hankel, J. Intestinal Microbiota of Fattening Pigs Offered Non-Fermented and Fermented Liquid Feed with and without the Supplementation of Non-Fermented Coarse Cereals. Microorganisms 2020, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Naumann, C.; Bassler, R. Band III Die chemische Untersuchung von Futtermitteln. In Methoden der landwirtschaftlichen Forschungs- und Untersuchungsanstalt; VDLUFA-Verlag: Darmstadt, Germany, 2012. [Google Scholar]
- Rodehutscord, M.; Rückert, C.; Maurer, H.P.; Schenkel, H.; Schipprack, W.; Bach Knudsen, K.E.; Schollenberger, M.; Laux, M.; Eklund, M.; Siegert, W. Variation in chemical composition and physical characteristics of cereal grains from different genotypes. Arch. Anim. Nutr. 2016, 70, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Schulten, L. Vergleichende Untersuchungen mit ileocaecal-fistulierten Miniaturschweinen sowie mit Hunden zur Verdaulichkeit ausgewählter Einzel- und Mischfuttermittel für Hunde. Ph.D. Thesis, University of Veterinary Medicine Hanover, Foundation, Hannover, Germany, 9 November 2018. [Google Scholar]
- Adejumo, O.I.; Adetunji, O.C.; Ogundipe, K.; Osademe, N.S. Chemical composition and amino acid profile of differently processed feather meal. J. Agric. Sci. 2016, 61, 237–246. [Google Scholar] [CrossRef]
- Bunte, S. Die Fermentation von Flüssigfutter als Fütterungskonzept im Schweinebestand-Potentiale, aber auch Risiken aus Sicht der Tierernährung und Tiermedizin. Ph.D. Thesis, Tierärztliche Hochschule Hannover, Hannover, Germany, 7 November 2018. [Google Scholar]
- NRC. Nutrient Requirements of Dogs and Cats; The National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Hankel, J.; Jung, K.; Kuder, H.; Keller, B.; Keller, C.; Galvez, E.; Strowig, T.; Visscher, C. Caecal Microbiota of Experimentally Camyplobacter jejuni-Infected Chickens at Different Ages. Front. Microbiol. 2019, 10, 2303. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Deng, P.; Swanson, K.S. Gut microbiota of humans, dogs and cats: Current knowledge and future opportunities and challenges. Br. J. Nutr. 2015, 113, S6–S17. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Li, Q.; Lauber, C.L.; Czarnecki-Maulden, G.; Pan, Y.; Hannah, S.S. Effects of the dietary protein and carbohydrate ratio on gut microbiomes in dogs of different body conditions. MBio 2017, 8, e01703–e01716. [Google Scholar] [CrossRef] [PubMed]
- Böswald, L.; Dobenecker, B.; Clauss, M.; Kienzle, E. A comparative meta-analysis on the relationship of faecal calcium and phosphorus excretion in mammals. J. Anim. Physiol. Anim. Nutr. 2018, 102, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Metzler, B.; Mosenthin, R. A review of interactions between dietary fiber and the gastrointestinal microbiota and their consequences on intestinal phosphorus metabolism in growing pigs. Asian Austral. J. Anim. 2008, 21, 603–615. [Google Scholar] [CrossRef]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the bacterial butyrate synthesis pathways by analyzing (meta) genomic data. MBio 2014, 5, e00889-14. [Google Scholar] [CrossRef]
- Kuhn, K.; Regner, E.H.; Mehta, G.; Hendrickson, J.D.; Whitney, A.K.; Campbell, E.L.; Ohri, N.; Colgan, S.P. Colonic intraepithelial lymphocytes produce IL-6 in response to resident bacteria to modulate epithelial barrier function. J. Immunol. 2016, 196 (Suppl. 1), 208.5. [Google Scholar]
- Mazmanian, S.K. Capsular polysaccharides of symbiotic bacteria modulate immune responses during experimental colitis. J. Pediatr. Gastroenterol. Nutr. 2008, 46, E11–E12. [Google Scholar]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef]
- Smith, C.J.; Rocha, E.R.; Paster, B.J. The Medically Important Bacteroides spp. in Health and Disease. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Knudsen, K.E.B.; Jensen, B.B.; Hansen, I. Digestion of polysaccharides and other major components in the small and large intestine of pigs fed on diets consisting of oat fractions rich in β-D-glucan. Br. J. Nutr. 1993, 70, 537–556. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.E.B.; Serena, A.; Kjær, A.K.B.; Jørgensen, H.; Engberg, R. Rye bread enhances the production and plasma concentration of butyrate but not the plasma concentrations of glucose and insulin in pigs. J. Nutr. 2005, 135, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.E.B.; Lærke, H.N. Rye arabinoxylans: Molecular structure, physicochemical properties and physiological effects in the gastrointestinal tract. Cereal Chem. 2010, 87, 353–362. [Google Scholar] [CrossRef]
- Cummings, J.; Hill, M.; Bone, E.; Branch, W.; Jenkins, D. The effect of meat protein and dietary fiber on colonic function and metabolism II. Bacterial metabolites in feces and urine. Am. J. Clin. Nutr. 1979, 32, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Van Dokkum, W.; Pikaar, N.; Thissen, J. Physiological effects of fibre-rich types of bread: 2. Dietary fibre from bread: Digestibility by the intestinal microflora and water-holding capacity in the colon of human subjects. Br. J. Nutr. 1983, 50, 61–74. [Google Scholar] [PubMed]
- McBurney, M.I.; van Soest, P.J.; Jeraci, J.L. Colonic carcinogenesis: The microbial feast or famine mechanism. Nutr. Cancer 1987, 10, 23–28. [Google Scholar] [CrossRef]
- Bingham, S.A. Mechanisms and experimental and epidemiological evidence relating dietary fibre (non-starch polysaccharides) and starch to protection against large bowel cancer. Proc. Nutr. Soc. 1990, 49, 153–171. [Google Scholar] [CrossRef]
- Hauner, H. Glykämischer Index und glykämische Last: Relevant für die Gesundheit? Akt. Ernähr. Med. 2006, 31, 42–48. [Google Scholar] [CrossRef]
- Isaksson, H.; Rakha, A.; Andersson, R.; Fredriksson, H.; Olsson, J.; Åman, P. Rye kernel breakfast increases satiety in the afternoon-an effect of food structure. Nutr. J. 2011, 10, 31. [Google Scholar] [CrossRef]
- Place, R.F.; Noonan, E.J.; Giardina, C. HDAC inhibition prevents NF-κB activation by suppressing proteasome activity: Down-regulation of proteasome subunit expression stabilizes IκBα. Biochem. Pharmacol. 2005, 70, 394–406. [Google Scholar] [CrossRef]
- Smith, J.G.; Yokoyama, W.H.; German, J.B. Butyric acid from the diet: Actions at the level of gene expression. Crit. Rev. Food Sci. 1998, 38, 259–297. [Google Scholar] [CrossRef]
- Tedelind, S.; Westberg, F.; Kjerrulf, M.; Vidal, A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826. [Google Scholar] [CrossRef] [PubMed]
- Theil, P.K.; Jørgensen, H.; Serena, A.; Hendrickson, J.; Knudsen, K.E.B. Products deriving from microbial fermentation are linked to insulinaemic response in pigs fed breads prepared from whole-wheat grain and wheat and rye ingredients. Br. J. Nutr. 2011, 105, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Tsubaki, J.; Hwa, V.; Twigg, S.M.; Rosenfeld, R.G. Differential activation of the IGF binding protein-3 promoter by butyrate in prostate cancer cells. Endocrinology 2002, 143, 1778–1788. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsubaki, J.; Choi, W.; Ingermann, A.; Twigg, S.; Kim, H.; Rosenfeld, R.; Oh, Y. Effects of sodium butyrate on expression of members of the IGF-binding protein superfamily in human mammary epithelial cells. J. Endocrinol. 2001, 169, 97–110. [Google Scholar] [CrossRef][Green Version]
- Williams, E.A.; Coxhead, J.M.; Mathers, J.C. Anti-cancer effects of butyrate: Use of micro-array technology to investigate mechanisms. Proc. Nutr. Soc. 2003, 62, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Musco, N.; Calabrò, S.; Tudisco, R.; Grossi, M.; Addi, L.; Moniello, G.; Lombardi, P.; Cutrignelli, M.I. Diet effect on short-and long-term glycaemic response in adult healthy cats. Vet. Ital. 2017, 53, 141–145. [Google Scholar]
- Lombardi, P.; Musco, N.; Calabrò, S.; Tudisco, R.; Mastellone, V.; Vastolo, A.; Infascelli, F.; Cutrignelli, M.I. Different carbohydrate sources affect swine performance and post-prandial glycaemic response. Ital. J. Anim. Sci. 2020, 19, 421–430. [Google Scholar] [CrossRef]
- Bruni, N.; Martello, E.; Fusi, E.; Meineri, G.; Giardini, A. Study of faecal parameters and body condition in dogs with a diet supplemented with Lactobacillus acidophilus D2/CSL (CECT 4529). Ital. J. Anim. Sci. 2020, 19, 704–711. [Google Scholar] [CrossRef]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial community evenness favours functionality under selective stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components (% as Fed) | Basic Diet (Vegetarian) | Basic Diet + Cornmeal (non-Vegetarian) | Basic Diet + Fermentate (non-Vegetarian) | Basic Diet + Rye (non-Vegetarian) |
|---|---|---|---|---|
| Wheat | 17.3 | 10.7 | 10.7 | 10.7 |
| Broken rice | 17.3 | 10.7 | 10.7 | 10.7 |
| Wheat gluten | 5.2 | 3.3 | 3.3 | 3.3 |
| Rice protein | 5.2 | 3.3 | 3.3 | 3.3 |
| Sunflower oil | 4.1 | 2.5 | 2.5 | 2.5 |
| Dried beet pulp | 1.8 | 1.1 | 1.1 | 1.1 |
| Brewer’s yeast | 1.2 | 0.7 | 0.7 | 0.7 |
| Linseed | 1.2 | 0.7 | 0.7 | 0.7 |
| Lignocellulose | 0.5 | 0.3 | 0.3 | 0.3 |
| Sum of additives 1 | 5.7 | 3.5 | 3.5 | 3.5 |
| Feather meal | - | 2.7 | 2.7 | 2.7 |
| Cornmeal | - | 20.1 | - | - |
| Rye | - | - | - | 20.1 |
| Fermented rye | - | - | 60.4 2 | |
| Water | 40.5 | 40.3 | - | 40.3 |
| Basic Diet (Vegetarian) | Basic Diet + Cornmeal (non-Vegetarian) | Basic Diet + Fermentate (non-Vegetarian) | Basic Diet + Rye (non-Vegetarian) | |
|---|---|---|---|---|
| Moisture (% as fed) | 44.9 | 39.7 | 41.3 | 42.7 |
| Crude ash | 53.8 | 42.6 | 41.8 | 41.3 |
| Crude fat | 67.5 | 72.7 | 71.2 | 62.9 |
| Crude protein | 220 | 218 | 214 | 214 |
| Crude fibre | 18.3 | 21.6 | 18.0 | 17.0 |
| N-free Extracts | 640 | 645 | 655 | 665 |
| Starch | 468 | 435 | 483 | 482 |
| Sugar | 26.3 | 42.2 | 39.9 | 48.0 |
| ME (MJ/100 g as fed) 1 | 0.89 | 0.98 | 0.96 | 0.93 |
| Calcium | 9.92 | 6.85 | 7.00 | 6.86 |
| Phosphorus | 3.86 | 3.01 | 3.09 | 2.99 |
| Lysine | 10.0 | 7.41 | 8.02 | 8.22 |
| Methionine | 6.13 | 4.29 | 4.35 | 4.45 |
| Threonine | 6.33 | 6.34 | 6.60 | 6.76 |
| Tryptophan | 2.25 | 1.74 | 1.93 | 1.98 |
| Phylum | Basic Diet (Vegetarian) | Basic Diet + Cornmeal (non-Vegetarian) | Basic Diet + Fermentate (non-Vegetarian) | Basic Diet + Rye (non-Vegetarian) | adj. p-Value 1 |
|---|---|---|---|---|---|
| Actinobacteria | 3.94 ± 3.16 | 7.56 ± 11.8 | 3.49 ± 2.97 | 2.48 ± 2.54 | 0.771 |
| Bacteroidetes | 26.7 ± 16.4 | 18.0 ± 14.8 | 43.2 ± 26.3 | 48.5 ± 25.2 | 0.378 |
| Firmicutes | 51.7 ± 17.1 | 61.3 ± 24.2 | 41.5 ± 21.0 | 37.7 ± 21.3 | 0.698 |
| Fusobacteria | 7.08 ± 10.5 | 4.24 ± 7.44 | 2.64 ± 2.20 | 0.48 ± 0.66 | 0.771 |
| Proteobacteria | 9.50 ± 6.58 | 7.76 ± 4.18 | 8.37 ± 2.86 | 8.81 ± 2.82 | 0.922 |
| Verrucomicrobia | 0.82 ± 0.72 | 0.98 ± 1.19 | 0.62 ± 0.63 | 1.57 ± 1.68 | 0.771 |
| F/B ratio | 2.47 | 5.12 | 2.17 | 1.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hankel, J.; Abd El-Wahab, A.; Grone, R.; Keller, B.; Galvez, E.; Strowig, T.; Visscher, C. Faecal Microbiota of Dogs Offered a Vegetarian Diet with or without the Supplementation of Feather Meal and either Cornmeal, Rye or Fermented Rye: A Preliminary Study. Microorganisms 2020, 8, 1363. https://doi.org/10.3390/microorganisms8091363
Hankel J, Abd El-Wahab A, Grone R, Keller B, Galvez E, Strowig T, Visscher C. Faecal Microbiota of Dogs Offered a Vegetarian Diet with or without the Supplementation of Feather Meal and either Cornmeal, Rye or Fermented Rye: A Preliminary Study. Microorganisms. 2020; 8(9):1363. https://doi.org/10.3390/microorganisms8091363
Chicago/Turabian StyleHankel, Julia, Amr Abd El-Wahab, Richard Grone, Birgit Keller, Eric Galvez, Till Strowig, and Christian Visscher. 2020. "Faecal Microbiota of Dogs Offered a Vegetarian Diet with or without the Supplementation of Feather Meal and either Cornmeal, Rye or Fermented Rye: A Preliminary Study" Microorganisms 8, no. 9: 1363. https://doi.org/10.3390/microorganisms8091363
APA StyleHankel, J., Abd El-Wahab, A., Grone, R., Keller, B., Galvez, E., Strowig, T., & Visscher, C. (2020). Faecal Microbiota of Dogs Offered a Vegetarian Diet with or without the Supplementation of Feather Meal and either Cornmeal, Rye or Fermented Rye: A Preliminary Study. Microorganisms, 8(9), 1363. https://doi.org/10.3390/microorganisms8091363