Effect of Lactic Acid Fermentation on Color, Phenolic Compounds and Antioxidant Activity in African Nightshade


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Starter Cultures and Fermentation of Nightshade Leaves
2.4. Physicochemical Properties and Ascorbic Acid Content
2.5. Quantification of Targeted Phenolic Compounds Using HPLC-DAD and Total Polyphenolic Content
2.6. Ferric Reducing Antioxidant Power (FRAP) Assay
2.7. Proximate Analysis of the Final Product
2.8. Statistical Analysis
3. Results and Discussion
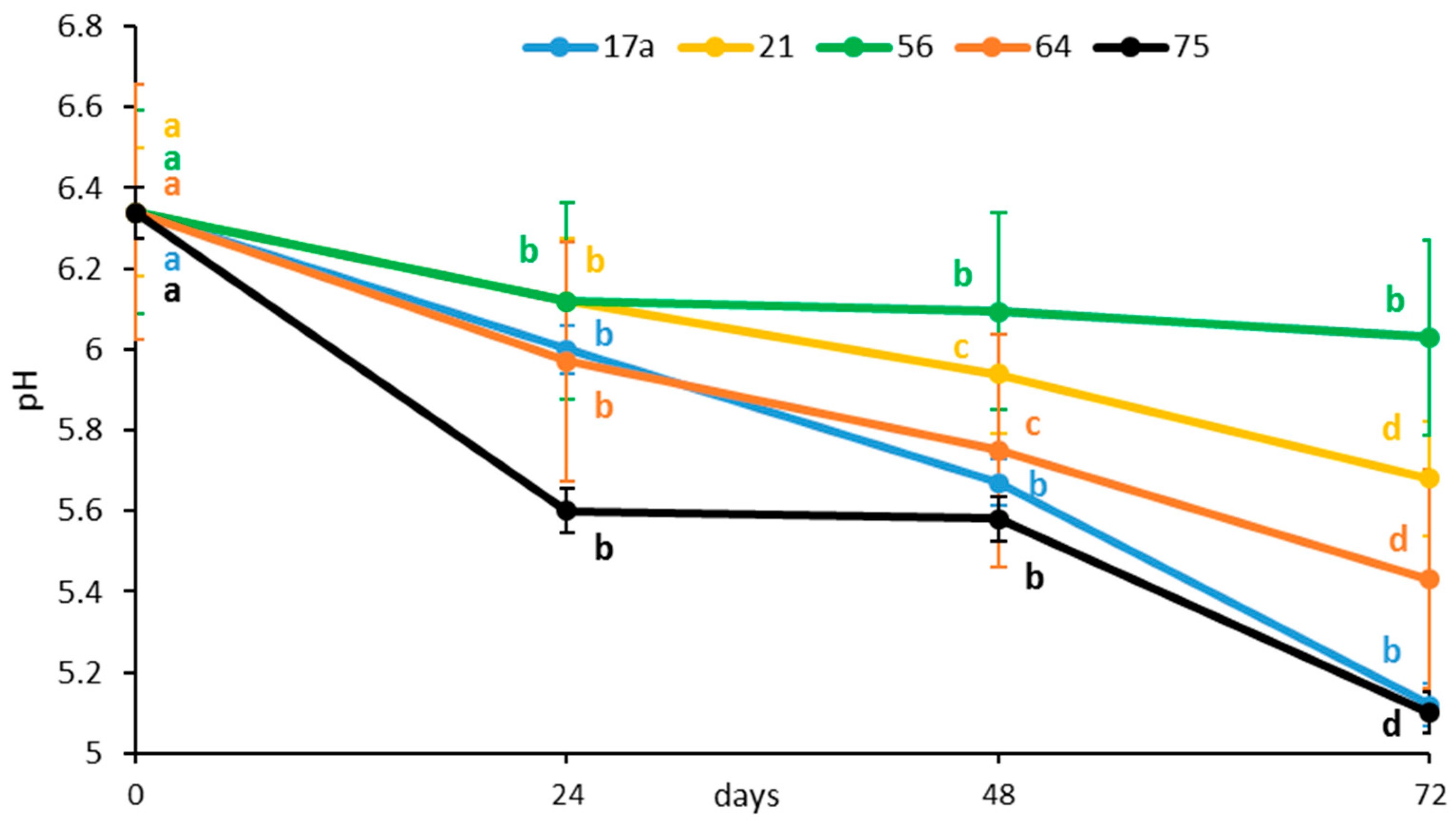
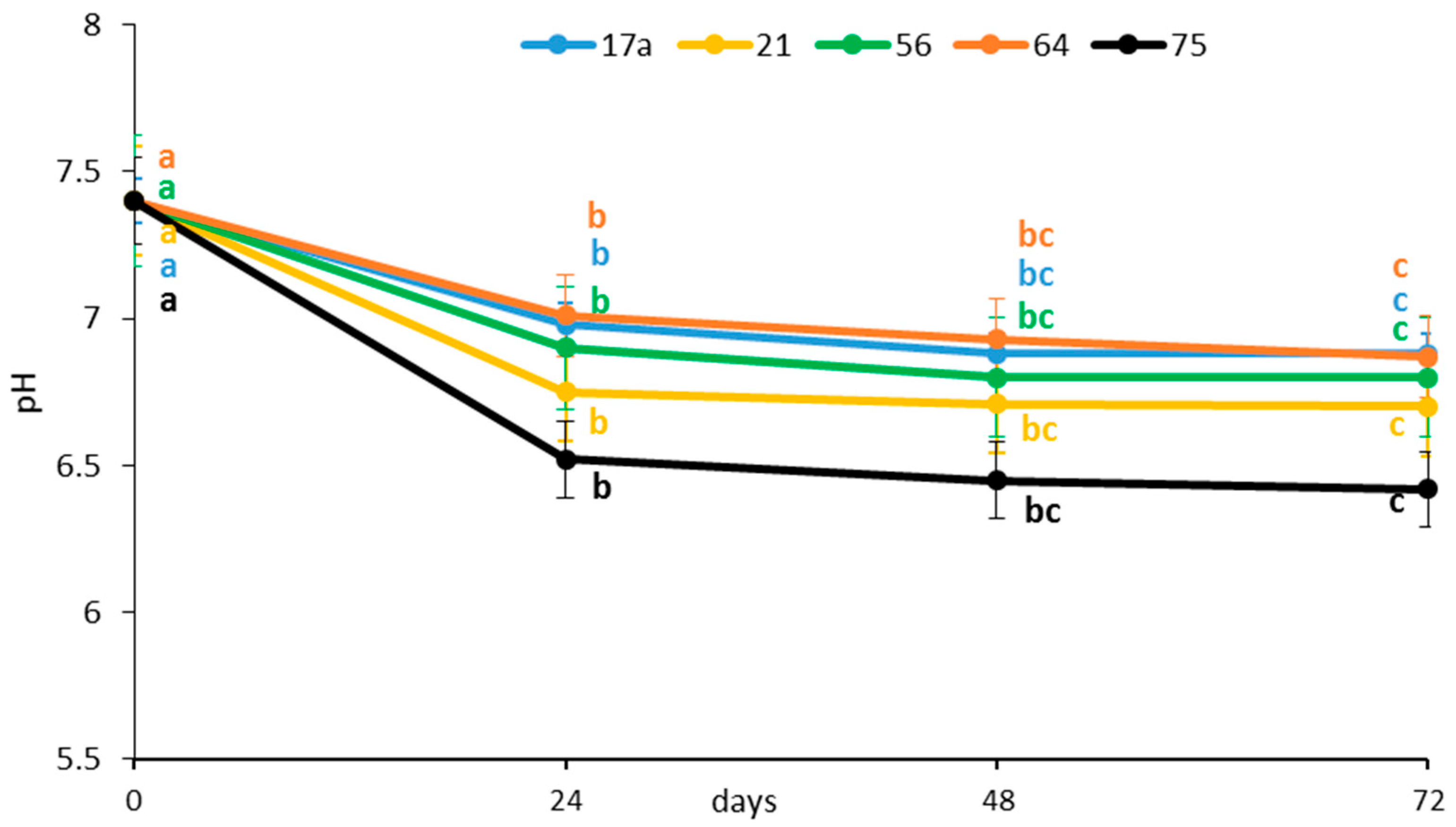
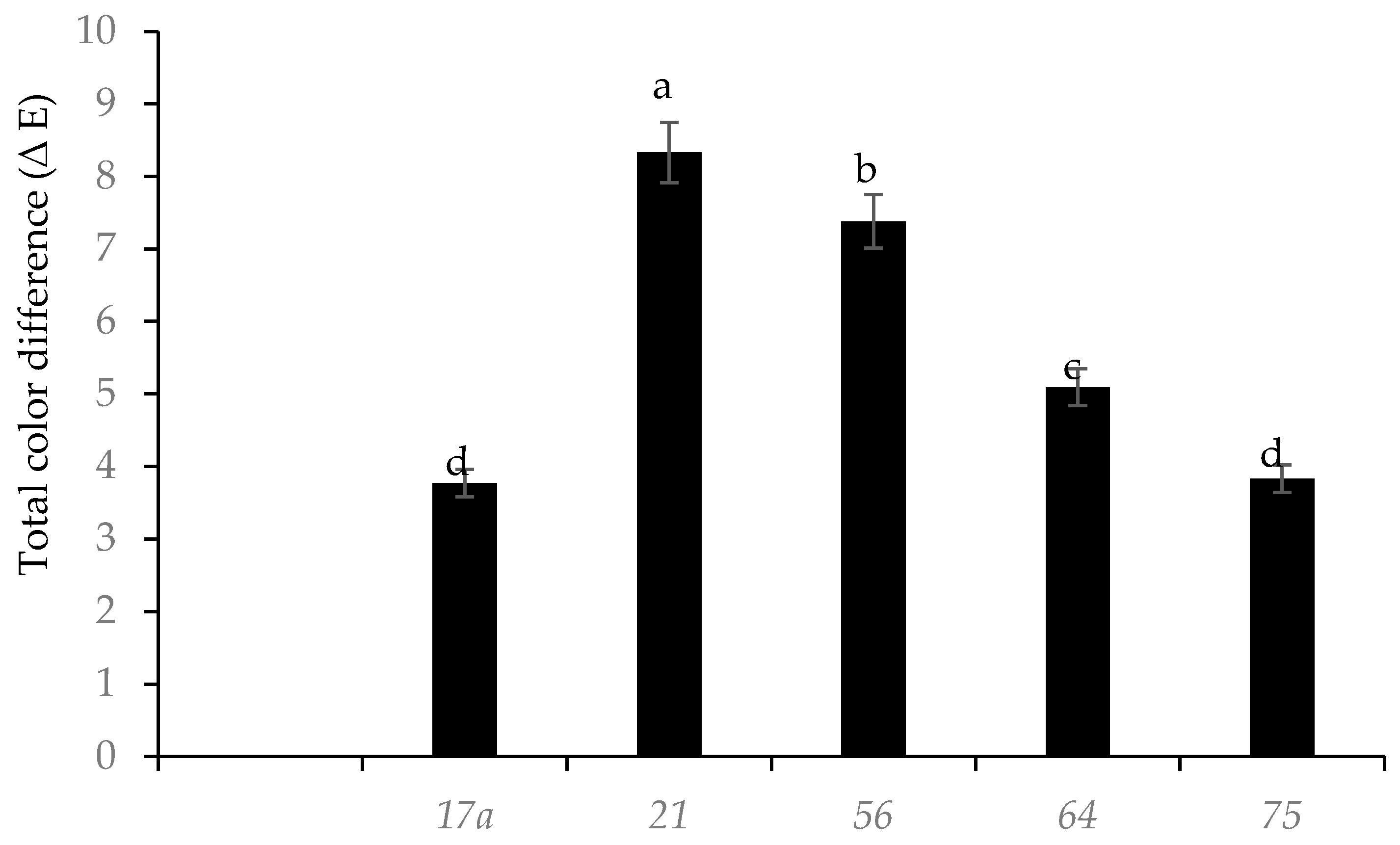
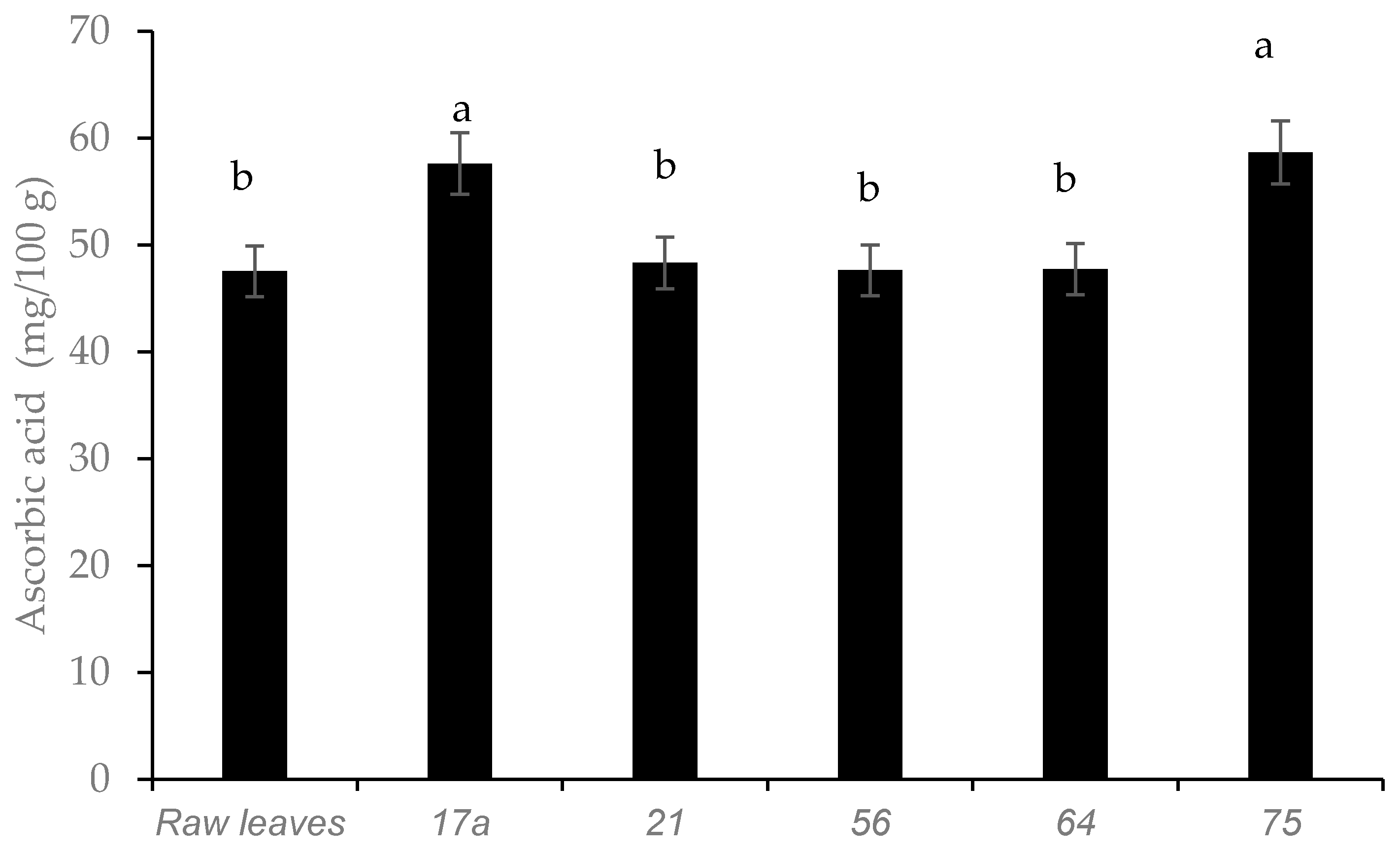
3.1. Effect of Fermentation on Physicochemical Parameters and Ascorbic Acid Content
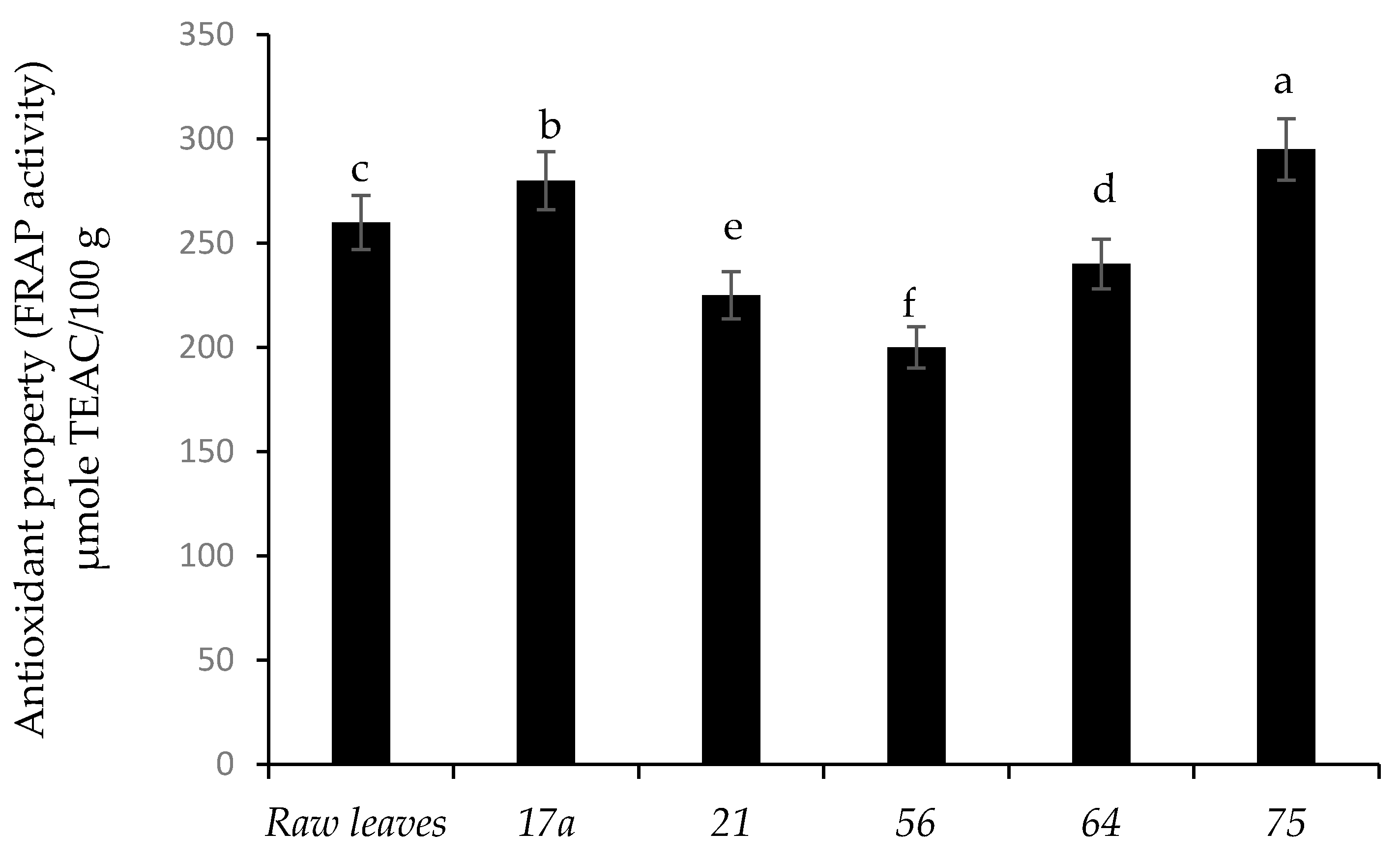
3.2. Phenolic Components and Antioxidant Activity
3.3. Proximate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| DAD | Diode array detector |
| FRAP | Ferric Reducing Antioxidant Power |
| GRAS | Generally regarded as safe |
| HPLC | High-performance liquid chromatography |
| LAB | Lactic acid bacterium |
| LSD | Least significant difference |
| TEAC | Trolox equivalent antioxidant capacity |
| TPC | Total polyphenolic content |
| TSS | Total soluble solids |
| MRS | Man Rogosa Sharpe |
| USDA | United States Department of Agriculture |
References
- Managa, M.; Sultanbawa, Y.; Sivakumar, D. Effects of different drying methods on untargeted phenolic metabolites, and antioxidant activity in Chinese cabbage (Brassica rapa L. subsp. chinensis) and nightshade (Solanum retroflexum Dun.). Molecules 2020, 25, 1326. [Google Scholar] [CrossRef] [PubMed]
- Oguntoyinbo, F.A.; Fusco, V.; Cho, G.-S.; Kabisch, J.; Neve, H.; Bockelmann, W.; Huch, M.; Frommherz, L.; Trierweiler, B.; Becker, B.; et al. Produce from Africa’s gardens: Potential for leafy vegetable and fruit fermentations. Front. Microbiol. 2016, 7, 981. [Google Scholar] [CrossRef] [PubMed]
- Fessard, A.; Remize, F. Genetic and technological characterization of lactic acid bacteria isolated from tropically grown fruits and vegetables. Int. J. Food Microbiol. 2019, 301, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Wen, J.; Hu, J.; Nie, Q.X.; Chen, H.H.; Nie, S.P.; Xiong, T.; Xie, M.Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–67. [Google Scholar] [CrossRef]
- Fujita, A.; Sarkar, D.; Genovese, M.L.; Shetty, K. Improving anti-hyperglycemic and anti-hypertensive bioactive properties of camu-camu (Myriciaria dubia Mc. Vaugh) using lactic acid bacterial fermentation. Process Biochem. 2017, 59, 133–140. [Google Scholar] [CrossRef]
- Wafula, E.N.; Franz, C.M.A.P.; Rohn, S.; Huch, M.; Mathara, J.M.; Trierweiler, B.; Becker, B. Fermentation of African Leafy Vegetables to Lower Postharvest Losses, Maintain Quality and Increase Product Safety. Afr. J. Hortic. Sci. 2016, 9, 13. [Google Scholar]
- Karovičová, J.; Kohajdová, Z. Lactic acid fermented vegetable juices. Hortic. Sci. 2011, 30, 152–158. [Google Scholar] [CrossRef]
- Quattrini, M.; Korcari, D.; Ricci, G.; Fortina, M.G. A polyphasic approach to characterize Weissella cibaria and Weissella confusa strains. J. Appl. Microbiol. 2020, 128, 500–512. [Google Scholar] [CrossRef]
- Srionnual, S.; Yanagida, F.; Lin, L.H.; Hsiao, K.N.; Chen, Y.S. Weissellicin 110, a newly discovered bacteriocin from Weissella cibaria 110, isolated from plaa-som, a fermented fish product from Thailand. Appl. Environ. Microbiol. 2007, 73, 2247–2250. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of AOAC International, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Mpai, S.; du Preez, R.; Sultanbawa, Y.; Sivakumar, D. Phytochemicals and nutritional composition in accessions of Kei-apple (Dovyalis caffra): Southern African indigenous fruit. Food Chem. 2018, 253, 37–45. [Google Scholar] [CrossRef]
- Zeb, A. A reversed phase HPLC-DAD method for the determination of phenolic compounds in plant leaves. Anal. Methods 2015, 18, 7753–7757. [Google Scholar] [CrossRef]
- Ruiz Rodríguez, L.G.; Mohamed, F.; Bleckwedel, J.; Medina, R.B.; De Vuyst, L.; Hebert, E.M.; Mozzi, F. Diversity and functional properties of lactic acid bacteria isolated from wild fruits and flowers present in Northern Argentina. Front. Microbiol. 2019, 10, 1019. [Google Scholar] [CrossRef] [PubMed]
- Fessard, A.; Kapoor, A.; Patche, J.; Assemat, S.; Hoarau, M.; Bourdon, E.; Bahorun, T.; Remize, F. Lactic fermentation as an efficient tool to enhance the antioxidant activity of tropical fruit juices and teas. Microorganisms 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Lübeck, M.; Stephensen Lübeck, P. Application of lactic acid bacteria in green biorefineries. FEMS Microbiol. Lett. 2019, 366, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Guan, Q.; Song, S.; Hao, M.; Xie, M. Dynamic change of lactic acid bacteria flora during Chinese sauerkraut fermentation. Food Control 2012, 26, 178–181. [Google Scholar] [CrossRef]
- Chen, L.; Opara, U.L. Texture measurement approaches in fresh and processed foods—A review. Food Res. Int. 2013, 51, 823–835. [Google Scholar] [CrossRef]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Sarkar, D.; Shetty, K. Fermentation-based biotransformation of bioactive phenolics and volatile compounds from cashew apple juice by select lactic acid bacteria. Process Biochem. 2017, 59, 141–149. [Google Scholar] [CrossRef]
- Zubaidah, E.; Ifadah, R.A.; Afgani, C.A. Changes in chemical characteristics of kombucha from various cultivars of snake fruit during fermentation. In International Conference on Green Agro-Industry and Bioeconomy, IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; Volume 230, p. 012098. [Google Scholar]
- Abou-Zaid, F.O.F.; Ibraheem, A.A. Using of some different acids in de-bittering OF green olives. J. Food. Dairy Sci. 2015, 6, 393–404. [Google Scholar] [CrossRef][Green Version]
- Ramírez, E.; Gandul-Rojas, B.; Romero, C.; Brenes, M.; Gallardo-Guerrero, L. Composition of pigments and color changes in green table olives related to processing type. Food Chem. 2015, 166, 115–124. [Google Scholar] [CrossRef]
- Filannino, P.; Cavoski, I.; Thlien, N.; Vincentini, O.; DeAngelis, M.; Silano, M.; Gobbetti, M.; Di Cagno, R. Lactic acid fermentation of cactus cladodes (Opuntia ficus-indica L.) generates flavonoid derivatives with antioxidant and anti-inflammatory properties. PLoS ONE 2016, 11, e0152575. [Google Scholar]
- Jagannath, A.; Raju, P.S.; Bawa, A.S. Controlled lactic fermentative stabilization of ascorbic acid in amaranthus paste. LWT-Food Sci. Technol. 2012, 48, 297–301. [Google Scholar] [CrossRef]
- Tayo, B.A.; Akpeji, S. Probiotic viability. Physicochemical and sensory properties of probiotic pineapple juice. Fermentation 2016, 2, 20. [Google Scholar]
- Apramita, D.; Appaiaha, K.A. Diverse physiological and metabolic adaptations by Lactobacillus plantarum and Oenococcus oeni in response to the phenolic stress during wine fermentation. Food Chem. 2018, 268, 101–109. [Google Scholar]
- Okcu, G.; Ayhan, K.; Altuntas, E.G.; Vural, N.; Poyrazoglu, E.S. Determination of phenolic acid decarboxylase produced by lactic acid bacteria isolated from shalgam (şalgam) juice using green analytical chemistry method. LWT-Food Sci Technol. 2016, 66, 615–621. [Google Scholar] [CrossRef]
- Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef]
- Silva, I.; Campos, F.M.; Hogg, T.; Couto, J.A. Wine phenolic compounds influence the production of volatile phenols by wine-related lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 360–370. [Google Scholar] [CrossRef]
- Jiménez-López, J.; Ruiz-Medina, A.; Ortega-Barrales, P.; Llorent-Martínez, E.J. Phytochemical profile and antioxidant activity of caper berries (Capparis spinosa L.): Evaluation of the influence of the fermentation process. Food Chem. 2018, 250, 54–59. [Google Scholar] [CrossRef]
- Gan, R.U.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Fermentation alters antioxidant capacity and polyphenol distribution in selected edible legumes. Int. J. Food Sci. Technol. 2016, 51, 875–884. [Google Scholar] [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Aryl glycosidases from Lactobacillus plantarum increase antioxidant activity of phenolic compounds. J. Funct. Foods 2014, 7, 322–329. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Sun, X.H.; Zhang, Q.; Yang, N. Biotransformation of phenolics and metabolites and the change in antioxidant activity in kiwifruit induced by Lactobacillus plantarum fermentation. J. Sci. Food Agric. 2020, 100, 3283–3290. [Google Scholar] [CrossRef]
- Kaltsa, A.; Papaliaga, D.; Papaioannou, E.; Kotzekidou, P. Characteristics of oleuropeinolytic strains of Lactobacillus plantarum group and influence on phenolic compounds in table olives elaborated under reduced salt conditions. Food Microbiol. 2015, 48, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G. Dietary Antioxidants and Prevention of Non-Communicable Diseases. Antioxidants 2018, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- USDA Dietary Guidelines. Dietary Guidelines for Americans 2015–2020. Available online: https://www.dietaryguidelines.gov/sites/default/files/2019-05/2015-2020_Dietary_Guidelines.pdf (accessed on 13 April 2020).
- Tamang, J.P.; Tamang, B.; Schillinger, U.; Guigas, C.; Holzapfel, W.H. Functional properties of lactic acid bacteria isolated from ethnic fermented vegetables of the Himalayas. Int. J. Food Microbiol. 2009, 135, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Alan, Y.; Topalcengiz, Z.; Dığrak, M. Biogenic amine and fermentation metabolite production assessments of Lactobacillus plantarum isolates for naturally fermented pickles. LWT-Food Sci. Technol. 2018, 98, 322–328. [Google Scholar] [CrossRef]
- Xu, X.; Luo, D.; Bao, Y.; Liao, X.; Wu, J. Characterization of diversity and probiotic efficiency of the autochthonous lactic acid bacteria in the fermentation of selected raw fruit and vegetable juices. Front. Microbiol. 2018, 9, 2539. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef]
- Szutowska, J. Functional properties of lactic acid bacteria in fermented fruit and vegetable juices: A systematic literature review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAB Strain | ||||||||
|---|---|---|---|---|---|---|---|---|
| Phenolic acids (mg/kg) | RT (min) | Raw leaves | 17a | 56 | 64 | 21 | 75 | |
| Gallic acid | C7H6OH5 | 6.78 | 162.8 ± 1.1 d | 178.0 ± 1.3 c | 150.5 ± 1.09 e | 171.5 ± 1.4 cd | 210.0 ± 1.7 b | 281.0 ± 1.2 a |
| Caffeic acid | C9H8O4 | 20.55 | 360.5 ± 1.4 b | 294 ± 0.98 c | 77.5 ± 0.21 d | 2145 ± 1.4 a | 395 ± 1.9 b | nd |
| Vanillic acid | C8H8O4 | 18.70 | 403.0 ± 0.5 b | 343 ± 2.3 c | 21.0 ± 1.7 e | 383.5 ± 0.67 b | 245 ± 0.98 d | 1352.0 ± 1.23 a |
| 2,5 Dihydroxybenzoic acid | C7H6O4 | 14.50 | 1593 ± 1.5 c | 2406 ± 0.7 b | 489 ± 1.8f | 1003.3 ± 2.2 e | 1267 ± 1.2 d | 2827.5 ± 1.4 a |
| p-Coumaric acid | C9H8O3 | 49.37 | 1115 ± 0.76 d | 1269.5 ± 1.8 c | 929 ± 1.8 e | 1387.0 ± 1.5 b | 980.5 ± 2.1 e | 1577 ± 0.56 a |
| Ferulic acid | C10H10O4 | 20.61 | 1321 ± 9.8 d | 2343.0 ± 1.2 a | 1030 ± 1.6 e | 2000 ± 1.9 b | 1487 ± 1.6 c | nd |
| Ellagic acid | C14H6O8 | 12.3 | nd | nd | 113.0 ± 1.4 c | nd | 191.0 ± 1.6 b | 453.5 ± 1.8 a |
| Total phenolic acids | 4955.3 ± 1.23 d | 6833.5 ± 1.41 b | 2810 ± 1.12 e | 7090.3 ± 1.70 a | 4775.5 ± 1.63 d | 6491 ± 1.16 c | ||
| Flavonoids (mg/kg) | ||||||||
| Catechin | C15H14O6 | 12.75 | 726.5 ± 2.5 b | 658.5 ± 1.3 cd | 574.5 ± 0.78 d | 669 ± 1.3 bc | 602.0 ± 0.45 c | 1322.0 ± 1.7 a |
| Quercetin | C15H10O7 | 62.34 | 326.0 ± 1.4 d | 498.5 ± 1.8 b | 438.0 ± 1.7 c | 116.0 ± 1.4f | 304.0 ± 1.1 e | 582.5 ± 1.7 a |
| Luteolin | C15H10O6 | 20.00 | nd | 256 ± 1.5 a | nd | 141.5 ± 2.1 c | nd | 242.5 ± 2.4 b |
| Total flavonoids | 1052.5 ± 1.21 c | 1413.0 ± 1.83 b | 1012.5 ± 1.62 c | 926.5 ± 0.95 d | 906.0 ± 1.86 d | 2147.0 ± 1.40 a | ||
| Total polyphenol content | 6007.8 ± 1.34 c | 8246.5 ± 1.50 b | 3822.5 ± 1.75 d | 8016.8 b | 5681.5 c | 8638 ± 1.69 a | ||
| Component | Per Fermented Vegetable (100 g Fresh Weight) |
|---|---|
| Energy (kJ) | 136.96 |
| Moisture (g) | 81.51 ± 0.22 * |
| Fat (g) | 0.23 ± 0.03 |
| Protein (g) | 3.86 ± 0.19 |
| Carbohydrate (g) | 2.51 ± 0.05 |
| Fibre (g) | 2.52 ± 0.30 |
| Ash (g) | 9.91 ± 0.50 |
| Na (sodium) (mg) | 231.00 ± 2.81 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degrain, A.; Manhivi, V.; Remize, F.; Garcia, C.; Sivakumar, D. Effect of Lactic Acid Fermentation on Color, Phenolic Compounds and Antioxidant Activity in African Nightshade. Microorganisms 2020, 8, 1324. https://doi.org/10.3390/microorganisms8091324
Degrain A, Manhivi V, Remize F, Garcia C, Sivakumar D. Effect of Lactic Acid Fermentation on Color, Phenolic Compounds and Antioxidant Activity in African Nightshade. Microorganisms. 2020; 8(9):1324. https://doi.org/10.3390/microorganisms8091324
Chicago/Turabian StyleDegrain, Alexandre, Vimbainashe Manhivi, Fabienne Remize, Cyrielle Garcia, and Dharini Sivakumar. 2020. "Effect of Lactic Acid Fermentation on Color, Phenolic Compounds and Antioxidant Activity in African Nightshade" Microorganisms 8, no. 9: 1324. https://doi.org/10.3390/microorganisms8091324
APA StyleDegrain, A., Manhivi, V., Remize, F., Garcia, C., & Sivakumar, D. (2020). Effect of Lactic Acid Fermentation on Color, Phenolic Compounds and Antioxidant Activity in African Nightshade. Microorganisms, 8(9), 1324. https://doi.org/10.3390/microorganisms8091324