Extension of the Shelf-Life of Fresh Pasta Using Chickpea Flour Fermented with Selected Lactic Acid Bacteria



Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Material, Bacterial Strains, and Fermentation
2.2. Chickpea Sourdough
2.3. Chemical and Microbiological Characterization of Chickpea Sourdough
2.4. Fungi, Culture Media, and Growth Conditions
2.5. Hyphal Radial Growth Rate Assay
2.6. Micro-Titer Plate Assay
2.7. Proteolysis and Heat Stability of Antifungal Compounds
2.8. Purification and Identification of Antifungal Compounds
2.9. Pasta Making and Shelf-Life Investigation
2.10. Nutritional Characterization
2.11. Optimal Cooking Time, Cooking Loss, and Water Absorption
2.12. Microbiological Shelf-Life
2.12.1. Microbiological and Chemical Analysis during Refrigerated Storage
2.12.2. Artificial Fungal Contamination and Antifungal Activity in Pasta
2.13. Statistical Analysis
3. Results
3.1. Chickpea Sourdough Fermentation
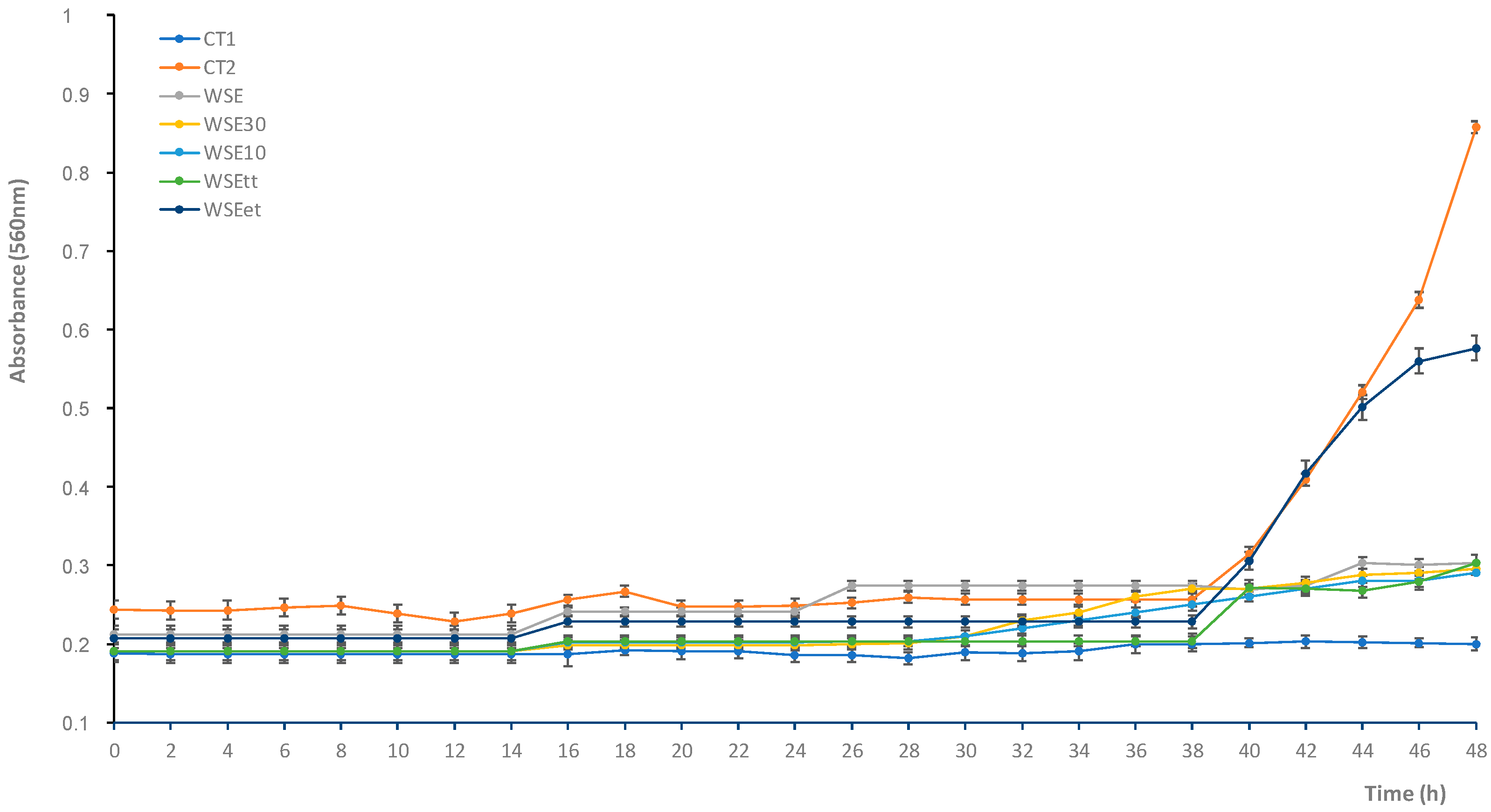
3.2. Antifungal Activity of the Water/Salt-Soluble Extract from Chickpea Sourdough
3.3. Purification and Identification of the Antifungal Compounds
3.4. Assessment of the Pasta Quality and Microbiological Shelf-Life
3.4.1. Nutritional and Technological Features
3.4.2. Microbial Dynamics under Refrigerated Conditions
3.4.3. Mold Growth Monitoring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lucera, A.; Costa, C.; Padalino, L.; Conte, A.; Lacivita, V.; Saccotelli, M.A.; Esposto, D.; Del Nobile, M.A. Combination of process technology and packaging conditions to improve the shelf life of fresh pasta. J. Food Process. Technol. 2014, 5, 12. [Google Scholar]
- Global Fresh Pasta Market Growth 2019–2024. Available online: https://www.fiormarkets.com/report/global-fresh-pasta-market-growth-2019-2024-390768.html (accessed on 26 June 2020).
- The Fresh Pasta Market and Sectoral Developments. Available online: http://www.magazinebbm.com/english/the-fresh-pasta-market-and-sectoral-developments/.html (accessed on 12 March 2018).
- Zardetto, S. Effect of modified atmosphere at abuse temperature on the growth of Penicillium aurantiogriseum isolated from fresh filled pasta. Food Microbiol. 2006, 14, 367–371. [Google Scholar] [CrossRef]
- Del Nobile, M.A.; Di Benedetto, N.; Suriano, N.; Conte, A.; Lamacchia, C.; Corbo, M.R.; Sinigaglia, M. Use of natural compounds to improve the microbial stability of Amaranth based homemade fresh pasta. Food Microbiol. 2009, 26, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Del Nobile, M.A.; Di Benedetto, N.; Suriano, N.; Conte, A.; Corbo, M.R.; Sinigaglia, M. Combined effects of chitosan and MAP to improve the microbial quality of amaranth homemade fresh pasta. Food Microbiol. 2009, 26, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Lucera, A.; Mastromatteo, M.; Conte, A.; Del Nobile, M.A. Shelf life extension of durum semolina-based fresh pasta. Int. J. Food Sci. Technol. 2010, 45, 1545–1551. [Google Scholar] [CrossRef]
- Oliveira, P.M.; Zannini, E.; Arendt, E.K. Cereal fungal infection, mycotoxins, and lactic acid bacteria mediated bioprotection: From crop farming to cereal products. Food Microbiol. 2014, 37, 78–95. [Google Scholar] [CrossRef]
- Zardetto, S.; Dalla Rosa, M. Effect of heat treatment of the microbiology and quality of Fresh Filled pasta. In New Issues in Food Policy, Control and Research; Arthur, P., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2007. [Google Scholar]
- Ahmed, J.; Ramaswamy, H.S. Microwave Pasteurization and sterilization of foods. In Handbook of Food Preservation, 2nd ed.; Tayler and Francis Group: Abingdon, UK, 2007. [Google Scholar]
- Chaudrasekaran, S.; Ramanthan, S.; Basak, T. Microwave food processing-A review. Food Res. Int. 2013, 52, 243–261. [Google Scholar] [CrossRef]
- Sanguinetti, A.M.; Del Caro, A.; Mangia, N.P.; Secchi, N.; Catzeddu, P.; Piga, A. Quality changes of fresh filled pasta during storage: Influence of modified atmosphere packaging on microbial growth and sensory properties. Food Sci. Technol. 2011, 17, 23–29. [Google Scholar] [CrossRef]
- Cruz, R.S.; Soares, N.F.F.; Andrade, N.J. Evaluation of oxygen absorber on antimicrobial preservation of lasagna -type fresh pasta under vacuum packed. Cienc. Agrotec. 2006, 30, 1135–1138. [Google Scholar] [CrossRef]
- Cassares, M.; Sakotani, N.L.; Kunigk, L.; Vasquez, P.; Jurkiewicz, C. Effect of gamma irradiation on shelf life extension of fresh pasta. Radiat. Phys. Chem. 2020, 174, 108940. [Google Scholar] [CrossRef]
- Heydaryinia, A.; Veissi, M.; Sodadi, A. A comparative study of the effects of the two preservatives, sodium benzoate and potassium sorbate on Aspergillus niger and Penicillium notatum. Jundishapur. J. Microbiol. 2011, 4, 301–307. [Google Scholar]
- Lucera, A.; Mastromatteo, M.; Conte, A.; Zambrini, A.V.; Faccia, M.; Del Nobile, M.A. Effect of active coating on microbiological and sensory properties of fresh mozzarella cheese. Food Packag. Shelf Life 2014, 1, 25–29. [Google Scholar] [CrossRef]
- Li, M.; Zhu, K.; Guo, X.; Peng, W.; Zhou, H. Effect of water activity (aw) and irradiation on the shelf-life of fresh noodles. Innov. Food Sci. Emerg. Technol. 2011, 12, 526–530. [Google Scholar] [CrossRef]
- Nionelli, L.; Wang, Y.; Pontonio, E.; Immonen, M.; Rizzello, C.G.; Maina, H.N.; Katina, K.; Coda, R. Antifungal effect of bioprocessed surplus bread as ingredient for bread-making: Identification of active compounds and impact on shelf-life. Food Control 2020, 118, 107437. [Google Scholar] [CrossRef]
- Manthey, F.A.; Sinha, S.; Wolf-Hall, C.E.; Hall, C.A., III. Effect of flaxseed flour and packaging on shelf life of refrigerated pasta. J. Food Process. Preserv. 2008, 32, 75–87. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Valdramidis, V.P.; O’Donnell, C.P.; Muthukumarappan, K.; Bourke, P.; Cullen, P.J. Application of natural antimicrobials for food preservation. J. Agric. Food Chem. 2009, 57, 5987–6000. [Google Scholar] [CrossRef]
- Miceli, A.; Francesca, N.; Moschetti, G.; Settanni, L. The influence of addition of Borago officinalis with antibacterial activity on the sensory quality of fresh pasta. Int. J. Gastron. Food Sci. 2015, 2, 93–97. [Google Scholar] [CrossRef][Green Version]
- Nielsen, P.V.; Rios, R. Inhibition of fungal growth on bread by volatile components from spices and herbs, and the possible application in active packaging, with special emphasis on mustard essential oil. Int. J. Food Microbiol. 2000, 60, 219–229. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Rutella, G.S.; Saa, D.L.T.; Gianotti, A. Bioactive peptides from vegetable food matrices: Research trends and novel biotechnologies for synthesis and recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Montemurro, M.; Coda, R.; Rizzello, C.G. Recent advances in the use of sourdough biotechnology in pasta making. Foods 2019, 8, 129. [Google Scholar] [CrossRef]
- Lorusso, A.; Verni, M.; Montemurro, M.; Coda, R.; Gobbetti, M.; Rizzello, C.G. Use of fermented quinoa flour for pasta making and evaluation of the technological and nutritional features. LWT Food Sci. Technol. 2017, 78, 215–221. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Verni, M.; Koivula, H.; Montemurro, M.; Seppa, L.; Kemell, M.; Katina, K.; Coda, R.; Gobbetti, M. Influence of fermented faba bean flour on the nutritional, technological and sensory quality of fortified pasta. Food Funct. 2017, 8, 860–871. [Google Scholar] [CrossRef] [PubMed]
- Schettino, R.; Pontonio, E.; Rizzello, C.G. Use of fermented hemp, chickpea and milling by-products to improve the nutritional value of semolina pasta. Foods 2019, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- AACC. Approved Methods of Analysis. 2010. Available online: http://methods.aaccnet.org/ (accessed on 18 December 2015).
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; De Angelis, M.; Gobbetti, M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901 and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Osborne, T.B. The Proteins of the Wheat Kernel, Carnegie Inst.; Carnegie Institution of Washington: Washington, DC, USA, 1907; p. 84. [Google Scholar] [CrossRef]
- Weiss, W.; Vogelmeier, C.; Görg, A. Electrophoretic characterization of wheat grain allergens from different cultivars involved in bakers’ asthma. Electrophoresis 1993, 14, 805–816. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; Di Cagno, R.; Gobbetti, M. Use of sourdough fermented wheat germ forenhancing the nutritional, texture and sensory characteristics of the white bread. Eur. Food Res. Technol. 2009, 230, 645–654. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Hagerman, A.E. Acid Buthanol Assay for Proanthocyanidins. In The Tannin Handbook 2002; Hagerman, A.E., Ed.; Miami University: Oxford, UK, 2002. [Google Scholar]
- Coda, R.; Rizzello, C.G.; Nigro, F.; De Angelis, M.; Arnault, P.; Gobbetti, M. Long-term fungal inhibitory activity of water-soluble extracts of Phaseolus vulgaris cv. pinto and sourdough lactic acid bacteria during bread storage. Appl. Environ. Microbiol. 2008, 74, 7391–7398. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Coda, R.; De Angelis, M.; Di Cagno, R.; Carnevali, P.; Gobbetti, M. Long-term fungal inhibitory activity of water-soluble extract from Amaranthus spp. seeds during storage of gluten-free and wheat flour breads. Int. J. Food Microbiol. 2009, 131, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, E.N.; Sampietro, A.R.; Vattuone, M.A. Screening antifungal activities of selected medicinal plants. J. Ethnopharmacol. 2001, 74, 89–96. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Hassan, Z.; Abu Bakar, F.; Algboory, H.L.; Saari, N. Novel antifungal peptides produced by Leuconostoc mesenteroides DU15 effectively inhibit growth of Aspergillus niger. J. Food Sci. 2015, 80, M1026–M1030. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, M.; Choiset, Y.; Dalgalarrondo, M.; Chobert, J.M.; Dousset, X.; Ivanova, I.; Haertlé, T. Isolation and partial biochemical characterization of a proteinaceous anti-bacteria and anti-yeast compound produced by Lactobacillus paracasei subsp. paracasei strain M3. Int. J. Food Microbiol. 2003, 87, 63–73. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Akeson, W.R.; Stahmann, M.A. A Pepsin Pancreatin Digest Index of Protein Quality Evaluation. J. Nutr. 1964, 83, 257–261. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Calasso, M.; Campanella, D.; De Angelis, M.; Gobbetti, M. Use of sourdough fermentation and mixture of wheat, chickpea, lentil and bean flours for enhancing the nutritional, texture and sensory characteristics of white bread. Int. J. Food Microbiol. 2014, 180, 78–87. [Google Scholar] [CrossRef]
- De Angelis, M.; Damiano, N.; Rizzello, C.G.; Cassone, A.; Di Cagno, R.; Gobbetti, M. Sourdough fermentation as a tool for the manufacture of low-glycemic index white wheat bread enriched in dietary fibre. Eur. Food Res. Technol. 2009, 229, 593–601. [Google Scholar] [CrossRef]
- Capriles, V.D.; Arêas, J.A.G. Effects of prebiotic inulin-type fructans on structure, quality, sensory acceptance and glycemic response of gluten-free breads. Food Funct. 2013, 4, 104–110. [Google Scholar] [CrossRef]
- Schoenlechner, R.; Drausinger, J.; Ottenschlaeger, V.; Jurackova, K.; Berghofer, E. Functional Properties of Gluten-Free Pasta Produced from Amaranth, Quinoa and Buckwheat. Plant Foods Hum. Nutr. 2010, 65, 339–349. [Google Scholar] [CrossRef]
- D’egidio, M.G.; Mariani, B.M.; Nardi, S.; Novaro, P.; Cubadda, R. Chemical and technological variables and their relationships: A predictive equation for pasta cooking quality. Cereal Chem. 1990, 67, 275–281. [Google Scholar]
- Suwaibah, G.; Abdulamir, A.S.; Fatimah, A.B.; Roselina, K.; Nazamid, S. Microbial growth, sensory characteristic and pH as potential spoilage indicators of Chinese yellow wet noodles from commercial processing plants. Am. J. Appl. Sci. 2009, 6, 1059–1066. [Google Scholar]
- Asioli, D.; Aschemann-Witzel, J.; Caputo, V.; Vecchio, R.; Annunziata, A.; Næs, T.; Varela, P. Making sense of the “clean label” trends: A review of consumer food choice behavior and discussion of industry implications. Food Res. Int. 2017, 99, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Axel, C.; Zannini, E.; Arendt, E.K. Mold spoilage of bread and its biopreservation: A review of current strategies for bread shelf life extension. Crit. Rev. Food Sci. Nutr. 2017, 57, 3528–3542. [Google Scholar] [CrossRef]
- De Pasquale, I.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Nutritional and functional effects of the lactic acid bacteria fermentation on gelatinized legume flours. Int. J. Food Microbiol. 2020, 316, 108426. [Google Scholar] [CrossRef]
- Curiel, J.A.; Coda, R.; Centomani, I.; Summo, C.; Gobbetti, M.; Rizzello, C.G. Exploitation of the nutritional and functional characteristics of traditional Italian legumes: The potential of sourdough fermentation. Int. J. Food Microbiol. 2015, 196, 51–61. [Google Scholar] [CrossRef]
- Coda, R.; Varis, J.; Verni, M.; Rizzello, C.G.; Katina, K. Improvement of the protein quality of wheat bread through faba bean sourdough addition. LWT Food Sci. Technol. 2017, 82, 296–302. [Google Scholar] [CrossRef]
- Coda, R.; Kianjam, M.; Pontonio, E.; Verni, M.; Di Cagno, R.; Katina, K.; Rizzello, C.G.; Gobbetti, M. Sourdough-type propagation of faba bean flour: Dynamics of microbial consortia and biochemical implications. Int. J. Food Microbiol. 2017, 248, 10–21. [Google Scholar] [CrossRef]
- Lomonaco, S.; Grassi, M.A.; Vallone, L.; Pistone, V.; Civera, T. Presence of fungal contamination in the production chain of fresh filled pasta, with particular regard to the Penicillium genus. Ital. J. Food Saf. 2011, 1, 33–37. [Google Scholar] [CrossRef]
- Corsetti, A.; Gobbetti, M.; Rossi, J.; Damiani, P. Antimould activity of sourdough lactic acid bacteria: Identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1. Appl. Microbiol. Biotechnol. 1998, 50, 253–256. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Ng, T.B. An antifungal protein from flageolet beans. Peptides 2005, 26, 2397–2403. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Lavecchia, A.; Gramaglia, V.; Gobbetti, M. Long-term fungal inhibition by Pisum sativum flour hydrolysate during storage of wheat flour bread. Appl. Environ. Microbiol. 2015, 81, 4195–4206. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Verni, M.; Bordignon, S.; Gramaglia, V.; Gobbetti, M. Hydrolysate from a mixture of legume flours with antifungal activity as an ingredient for prolonging the shelf-life of wheat bread. Food Microbiol. 2017, 64, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Y.; Ng, T.B. Isolation of a new cyclophilin-like protein from chickpeas with mitogenic, antifungal and anti-HIV-1 reverse transcriptase activities. Life Sci. 2002, 70, 1129–1138. [Google Scholar] [CrossRef]
- Chu, K.T.; Liu, K.H.; Ng, T.B. Cicerarin, a novel antifungal peptide from the green chickpea. Peptides 2003, 24, 659–663. [Google Scholar] [CrossRef]
- Thery, T.; Lynch, K.M.; Arendt, E.K. Natural antifungal peptides/proteins as model for novel food preservatives. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1327–1360. [Google Scholar] [CrossRef]
- Wang, G. Improved methods for classification, prediction and design of antimicrobial peptides. In Computational Peptidology; Zhou, P., Huang, J., Eds.; Humana Press: New York, NY, USA, 2015; pp. 43–66. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Hassan, Z.; Abu Bakar, F.; Saari, N. Identification of antifungal peptides produced by Lactobacillus plantarum IS10 grown in the MRS broth. Food Control 2016, 59, 27–30. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Chilosi, G.; Caruso, C.; Caporale, C.; Leonardi, L.; Bertini, L.; Buzi, A.; Nobile, M.; Magro, P.; Buonocore, V. Antifungal activity of a Bowman–Birk type trypsin inhibitor from wheat kernel. J. Phytopathol. 2000, 148, 477–481. [Google Scholar] [CrossRef]
- Ye, X.Y.; Ng, T.B.; Rao, P.F. A Bowman–Birk-type trypsin–chymotrypsin inhibitor from broad beans. Biochem. Biophys. Res. Commun. 2001, 289, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B. Isolation of cucurmoschin, a novel antifungal peptide abundant in arginine, glutamate and glycine residues from black pumpkin seeds. Peptides 2003, 24, 969–972. [Google Scholar] [CrossRef]
- Rogozhin, E.; Ryazantsev, D.; Smirnov, A.; Zavriev, S. Primary structure analysis of antifungal peptides from cultivated and wild cereals. Plants 2018, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Osorio-Díaz, P.; Agama-Acevedo, E.; Mendoza-Vinalay, M.; Tovar, J.; Bello-Pérez, L.A. Pasta added with chickpea flour: Chemical composition, in vitro starch digestibility and predicted glycemic index. Cienc. Tecnol. Aliment. 2008, 6, 6–12. [Google Scholar] [CrossRef]
- Regulation (EC) No 1924/2006 of the European Parliament and of the Council of 20 December 2006 on nutrition and health claims made on foods. Off. J. Eur. Union L 1924, 404, 9–25.
- Handa, V.; Sharma, D.; Kaur, A.; Arya, S.K. Biotechnological applications of microbial phytase and phytic acid in food and feed industries. Biocatal. Agric. Biotechnol. 2020, 25, 101600. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Semolina (%) | Water (%) | Chickpea Sourdough | Calcium Propionate (%) | ||
|---|---|---|---|---|---|
| Chickpea Flour (%) | Water (%) | ||||
| CS-p | 63.9 | 24.7 | 7.1 | 4.3 | |
| CT-p | 71.0 | 29.0 | |||
| cP-p | 70.7 | 29.0 | 0.3 | ||
| CS-T0 | CS-T24 | |
|---|---|---|
| Lactic acid bacteria (log10 cfu/g) | 7.28 ± 0.24 b | 9.78 ± 0.11 a |
| pH | 6.2 ± 0.2 a | 4.3 ± 0.1 b |
| TTA | 4.6 ± 0.3 b | 28.1 ± 1.2 a |
| Lactic acid (mmol/kg) | n.d | 180.7 ± 4.3 |
| Acetic acid (mmol/kg) | n.d. | 42.7 ± 0.7 |
| Fermentation quotient (FQ) | - | 4.3 ± 0.2 |
| Peptides (mg/kg) | 750 ± 16 b | 2600 ± 50 a |
| TFAA (mg/kg) | 965 ± 10 b | 2049 ± 15 a |
| Phytic acid g/100g | 2.72 ± 0.05 a | 0.13 ± 0.05 b |
| Raffinose g/kg | 1.88 ± 0.09 a | 0.60 ± 0.06 b |
| Condensed tannins g/kg | 0.90 ± 0.02 a | 0.45 ± 0.05 b |
| Fraction | Sequence | Experimental Mass (Da) | Calculated Mass (Da) | Δmass (Da) | Length | Net Charge | Hydrophobic Ratio (%) | Accession Number (Protein) | Fragment |
|---|---|---|---|---|---|---|---|---|---|
| B | ISSATSAIADKA | 1133.5914 | 1133.5928 | 0.0014 | 12 | 0 | 50 | XP_004515723.2 (LIM domain-containing protein A-like isoform X1) | 645–656 |
| C | NLESTEQGKGGSDVLGAVKE | 2016.9928 | 2016.9964 | 0.0036 | 20 | −2 | 25 | XP_004487170.1 (seed biotin-containing protein SBP65) | 559–578 |
| TGQYADDAAEDTRDFAGE | 1930.7765 | 1930.7817 | 0.0052 | 18 | −5 | 27 | XP_004508082.1 (embryonic protein DC-8-like) | 177–194 | |
| KIQDVATGAGEYTAEK | 1679.8348 | 1679.8366 | 0.0018 | 16 | −1 | 31 | 373–388 | ||
| ASNIADAGKDTFEAAK | 1607.7778 | 1607.7791 | 0.0013 | 16 | −1 | 43 | 389–404 | ||
| D | AGESIKETAANVGASAK | 1602.8208 | 1602.8213 | 0.0005 | 17 | 0 | 41 | NP_001266059.1 (18 kDa seed maturation protein-like) | 7–23 |
| ESAGFVGHETATNIAR | 1658.8004 | 1658.8012 | 0.0008 | 16 | −0.75 | 37 | XP_012567985.1 (late embryogenesis abundant protein D-34-like isoform X1) | 67–82 | |
| E | NVVSSIGETVGK | 1188.6354 | 1188.6350 | 0.0004 | 12 | 0 | 33 | XP_004487170.1 (seed biotin-containing protein SBP65) | 491–502 |
| VKSTTTACCDSCVCTK | 1876.7769 | 1876.7788 | 0.0019 | 16 | 1 | 43 | XP_004492724.1 (Bowman-Birk type proteinase inhibitor-like) | 49–64 | |
| SVADVAGYVGQK | 1192.6079 | 1192.6088 | 0.0009 | 12 | 0 | 41 | XP_004489219.1 (P24 oleosin isoform A-like) | 156–167 |
| CS-p | CT-p | cP-p | |
|---|---|---|---|
| Moisture (%) | 28.1 ± 0.27 a | 28.5 ± 0.15 a | 28.2 ± 0.18 a |
| pH | 5.1 ± 0.2 b | 6.5 ± 0.2 a | 6.5 ± 0.2 a |
| TTA | 4.8 ± 0.35 a | 2.10 ± 0.22 b | 2.15 ± 0.17 b |
| Water activity | 0.971 ± 0.002 b | 0.975 ± 0.002 a | 0.975 ± 0.002 a |
| Protein (%) | 11.2 ± 0.27 a | 10.25 ± 0.09 b | 10.12 ± 0.08 b |
| Fat (%) | 1.40 ± 0.02 a | 1.35 ± 0.10 a | 1.33 ± 0.04 a |
| Available carbohydrates (%) | 53.15 ± 0.21 b | 55.32 ± 0.17 a | 55.33 ± 0.10 a |
| Total dietary fibers (%) | 4.37 ± 0.31 a | 2.98 ± 0.08 b | 2.88 ± 0.18 b |
| Ash (%) | 1.05 ± 0.05 a | 0.92 ± 0.05 b | 0.94 ± 0.03 b |
| HI (%) | 50.61 ± 1.23 b | 67.5 ± 1.1 a | 67.0 ± 1.8 a |
| pGI | 67.49 ± 1.04 b | 76.76 ± 0.98 a | 74.35 ± 1.98 a |
| IVPD (%) | 88.80 ± 0.50 a | 65.1 ± 1.42 b | 63.1 ± 2.02 b |
| CS-p | CT-p | cP-p | |
|---|---|---|---|
| OCT (min) | 3.10 ± 0.20 b | 4.25 ± 0.20 a | 4.30 ± 0.10 a |
| Water Absorption (%) | 85 ± 2 a | 80 ± 2 b | 80 ± 2 b |
| Cooking loss (% of d.m.) | 4.57 ± 0.12 a | 3.91 ± 0.10 b | 3.80 ± 0.15 b |
| pH (at the OCT) | 6.0 ± 0.1 b | 6.7 ± 0.2 a | 6.7 ± 0.1 a |
| CS-p | CT-p | cP-p | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Storage time (days) | 0 | 10 | 20 | 30 | 40 | 0 | 10 | 20 | 30 | 40 | 0 | 10 | 20 | 30 | 40 |
| Moisture | 28.1 ± 0.27 a | 27.5 ± 0.07 b | 27.1 ± 0.11 c | 26.5 ± 0.12 d | 26.2 ± 0.07 e | 28.5 ± 0.15 a | 27.4 ± 0.10 b | 27.1 ± 0.08 c | 26.6 ± 0.05 d | 26.3 ± 0.05 e | 28.2 ± 0.18 a | 27.5 ± 0.13 b | 27.2 ± 0.09 c | 26.5 ± 0.10 d | 26.2 ± 0.13 e |
| pH | 5.1 ± 0.2 c | 4.8 ± 0.1 d | 4.5 ± 0.1 e | 4.4 ± 0.2 e | 4.4 ± 0.2 e | 6.5 ± 0.2 a | 6.0 ± 0.1 b | 5.8 ± 0.2 b | 5.8 ± 0.2 b | 5.7 ± 0.1 b | 6.5 ± 0.2 a | 6.3 ± 0.2 a | 6.2 ± 0.1 a | 5.9 ± 0.2 b | 5.8 ± 0.2 b |
| FC | − | − | − | + | + | − | ++ | +++ | ++++ | ++++ | − | − | − | ++ | +++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schettino, R.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Extension of the Shelf-Life of Fresh Pasta Using Chickpea Flour Fermented with Selected Lactic Acid Bacteria. Microorganisms 2020, 8, 1322. https://doi.org/10.3390/microorganisms8091322
Schettino R, Pontonio E, Gobbetti M, Rizzello CG. Extension of the Shelf-Life of Fresh Pasta Using Chickpea Flour Fermented with Selected Lactic Acid Bacteria. Microorganisms. 2020; 8(9):1322. https://doi.org/10.3390/microorganisms8091322
Chicago/Turabian StyleSchettino, Rosa, Erica Pontonio, Marco Gobbetti, and Carlo Giuseppe Rizzello. 2020. "Extension of the Shelf-Life of Fresh Pasta Using Chickpea Flour Fermented with Selected Lactic Acid Bacteria" Microorganisms 8, no. 9: 1322. https://doi.org/10.3390/microorganisms8091322
APA StyleSchettino, R., Pontonio, E., Gobbetti, M., & Rizzello, C. G. (2020). Extension of the Shelf-Life of Fresh Pasta Using Chickpea Flour Fermented with Selected Lactic Acid Bacteria. Microorganisms, 8(9), 1322. https://doi.org/10.3390/microorganisms8091322