Cyclic Di-adenosine Monophosphate Regulates Metabolism and Growth in the Oral Commensal Streptococcus mitis

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth
2.2. Bioinformatics Analyses
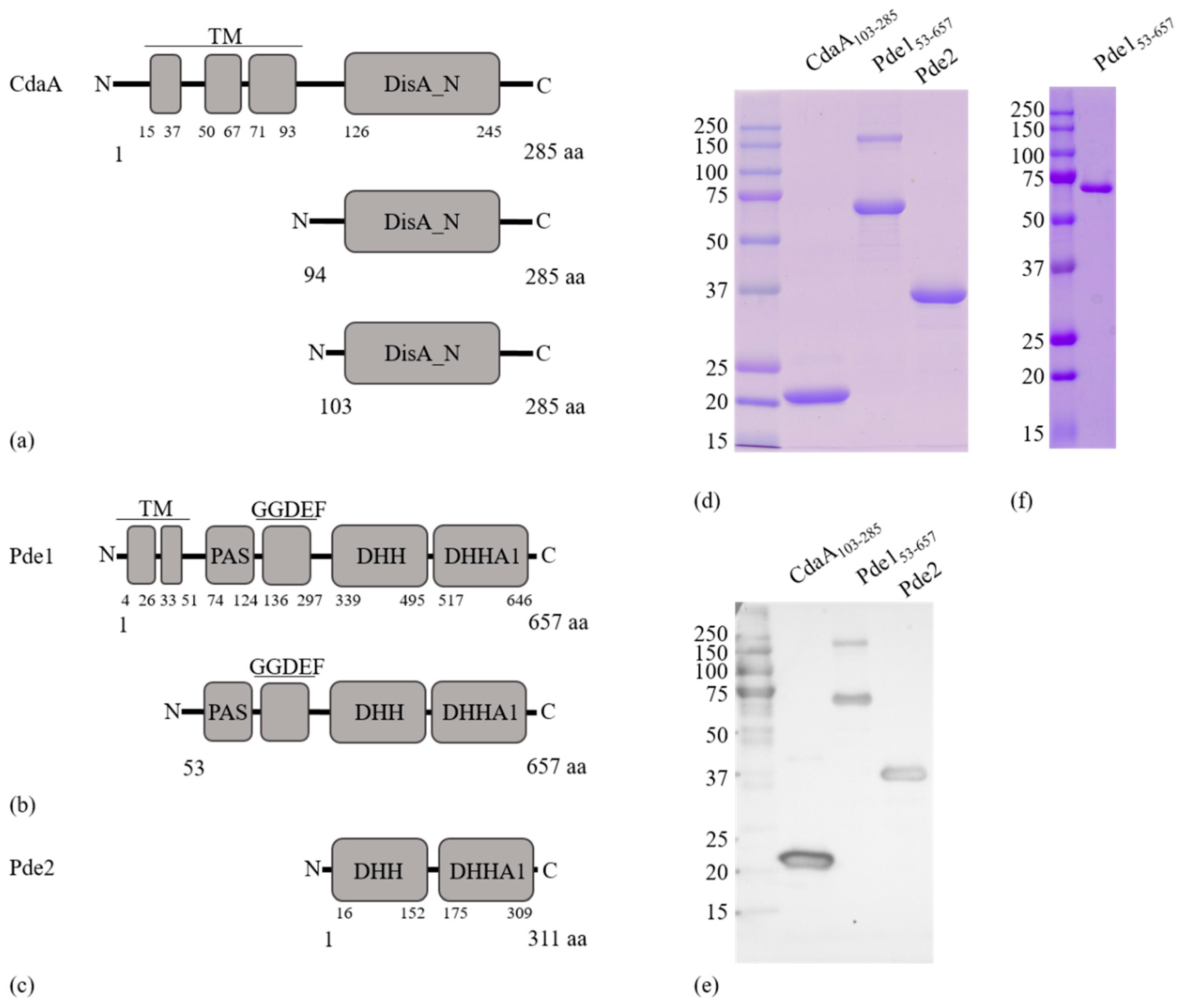
2.3. Cloning and Heterologous Expression of cdaA, pde1 and pde2
2.4. Protein Purification
2.5. Determination of the Quaternary Protein Structure
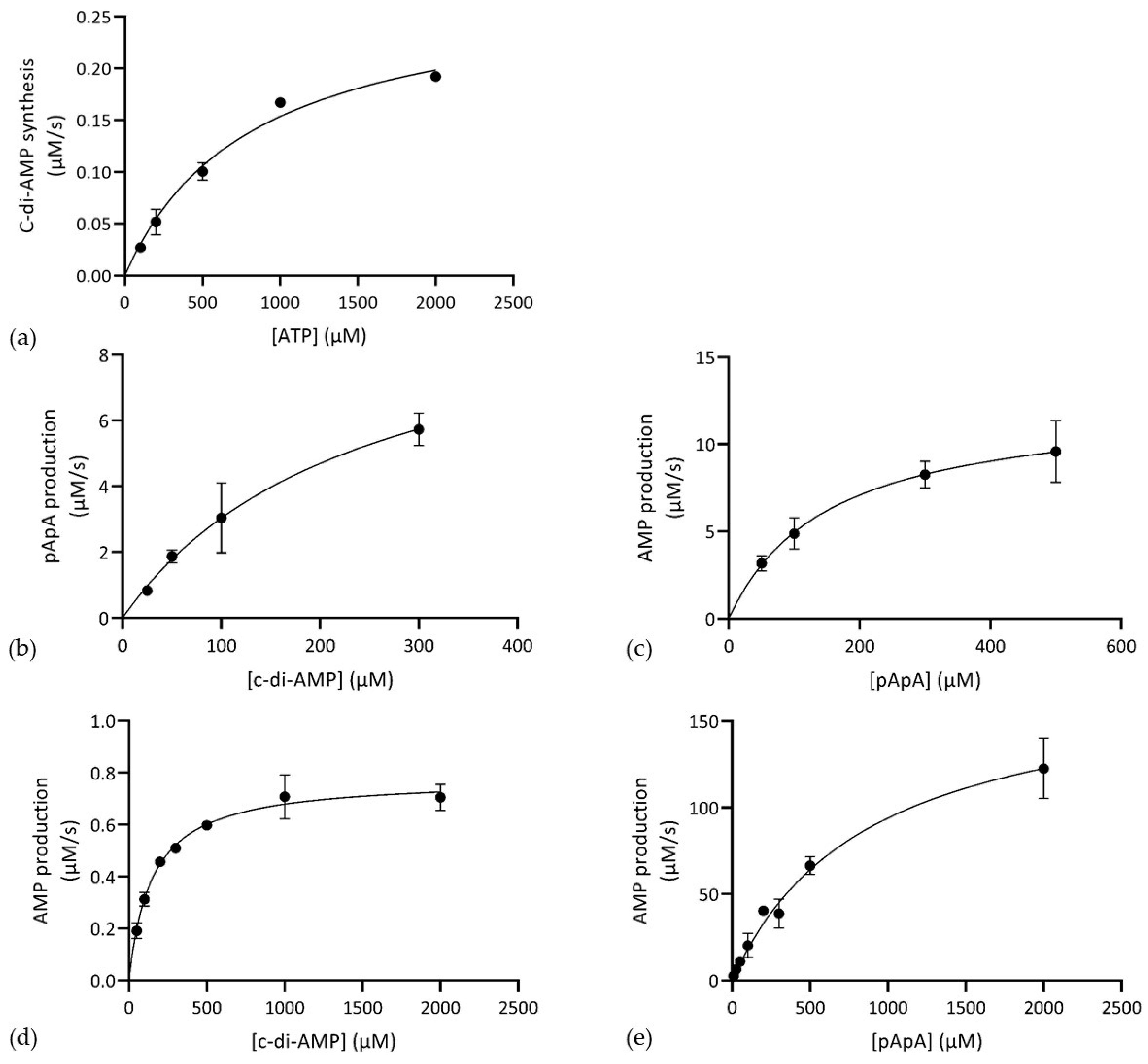
2.6. In Vitro Analysis of Enzyme Activity and Enzyme Kinetics Activity Assays
2.7. Kinetics Assays
2.8. Quantification of Nucleotides by Reversed Phase Chromatography
2.9. Mutant Construction
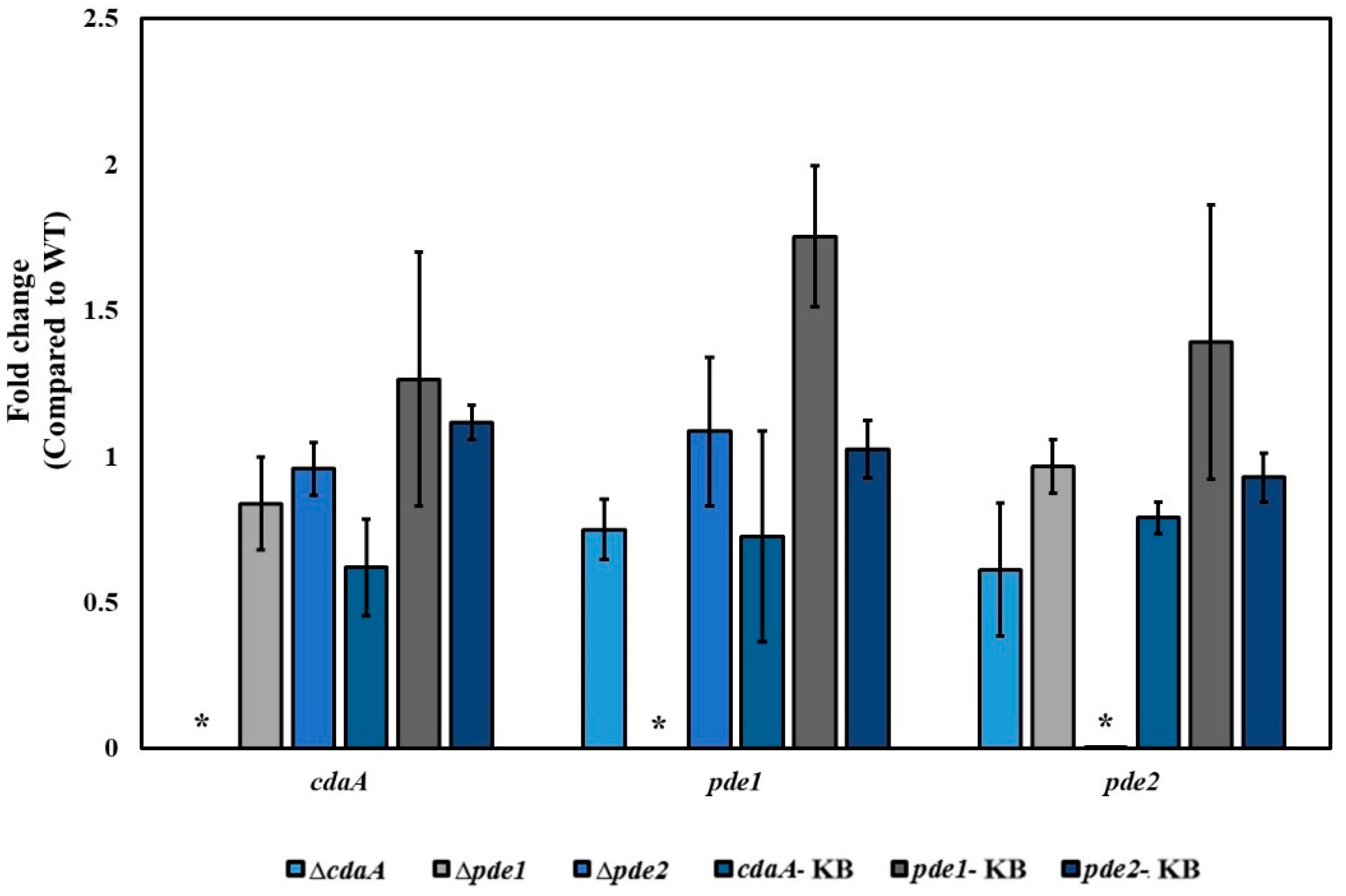
2.10. RNA Isolation and Real Time Quantitative PCR (RT- qPCR)
2.11. Colony Size
2.12. Growth Assays
2.13. Glucose Metabolism
2.14. C-di-AMP and pApA Profiling by HRMS/MS
2.15. Determination of Chain Length
2.16. Flow Cytometry Analysis
2.17. Transformation Efficiency
3. Results
3.1. Identification of c-di-AMP Turnover Proteins in S. mitis
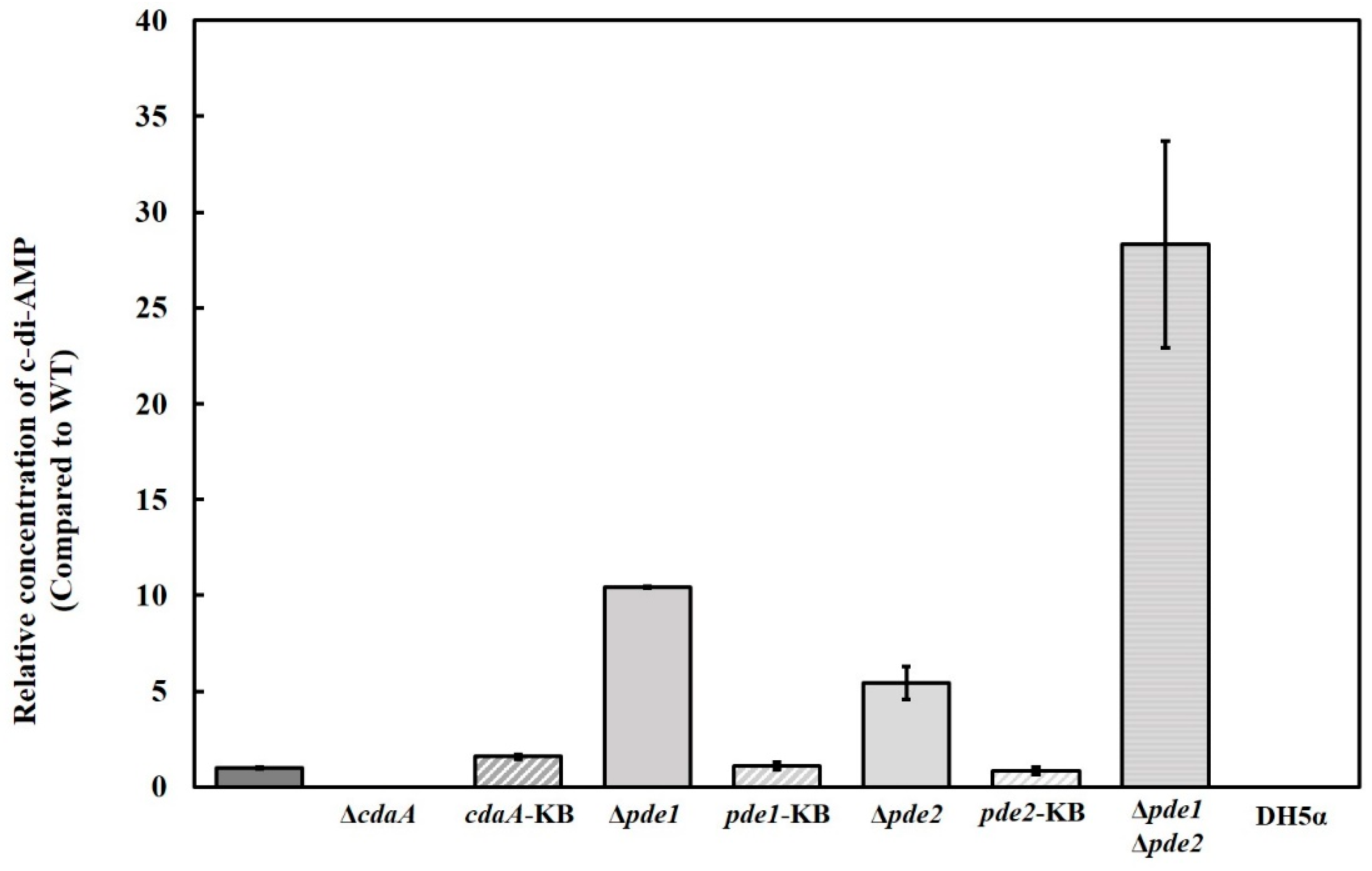
3.2. Deletion Mutants Displayed Altered Intracellular Concentration of c-di-AMP and pApA
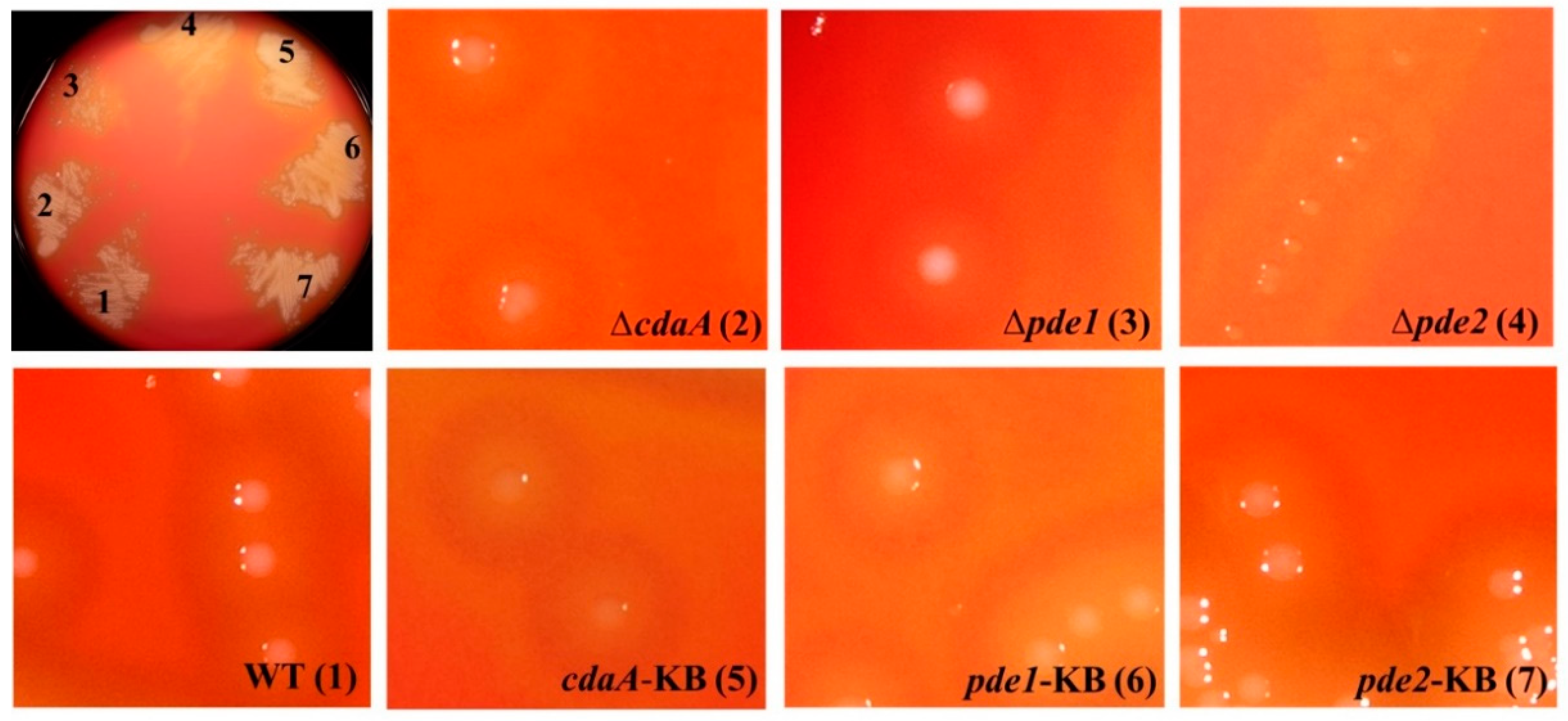
3.3. Deletion of pde2 Has a Pronounced Effect on S. mitis Colony Morphology
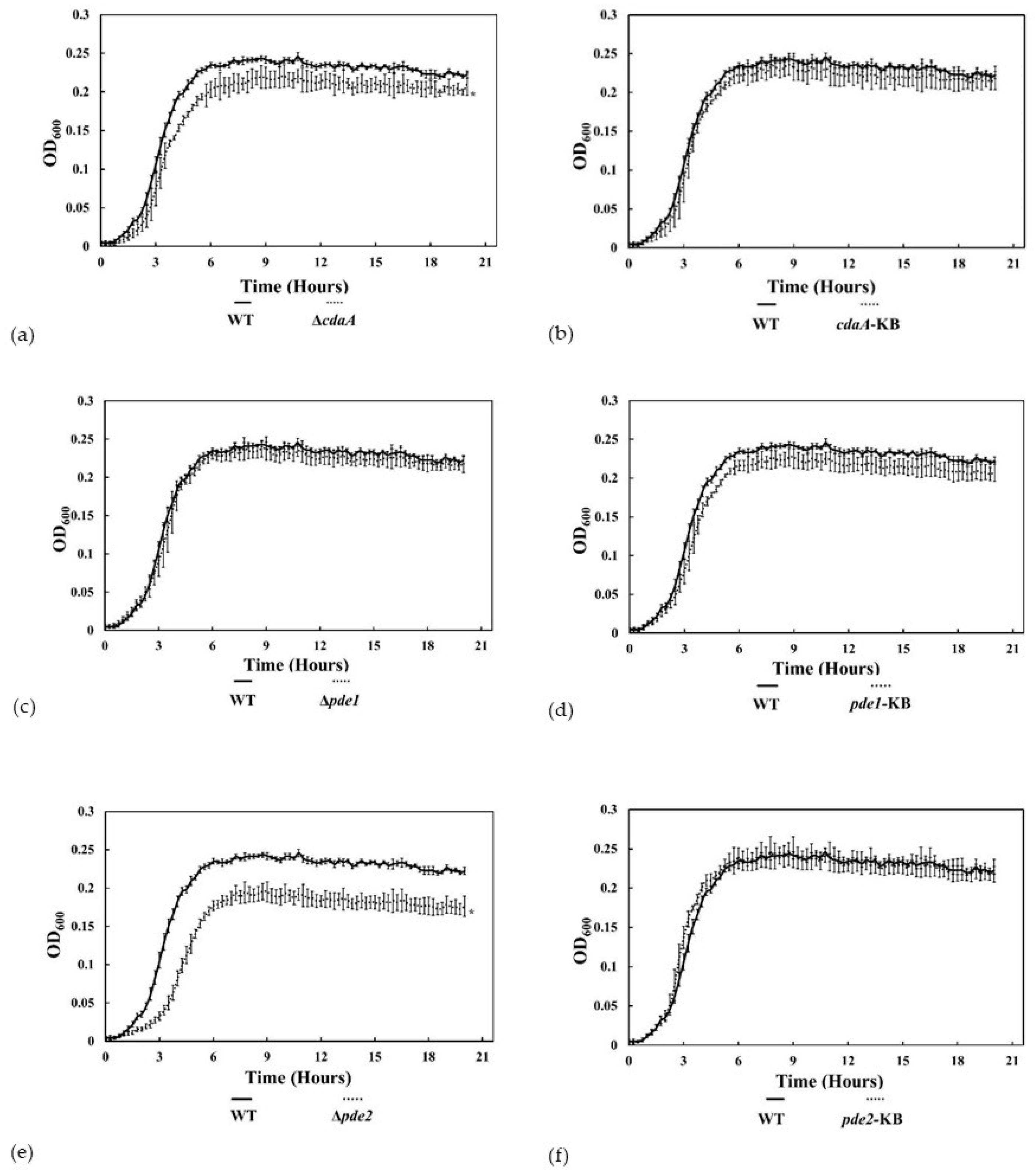
3.4. The ∆pde2-Mutant Displayed Reduced Growth Rate
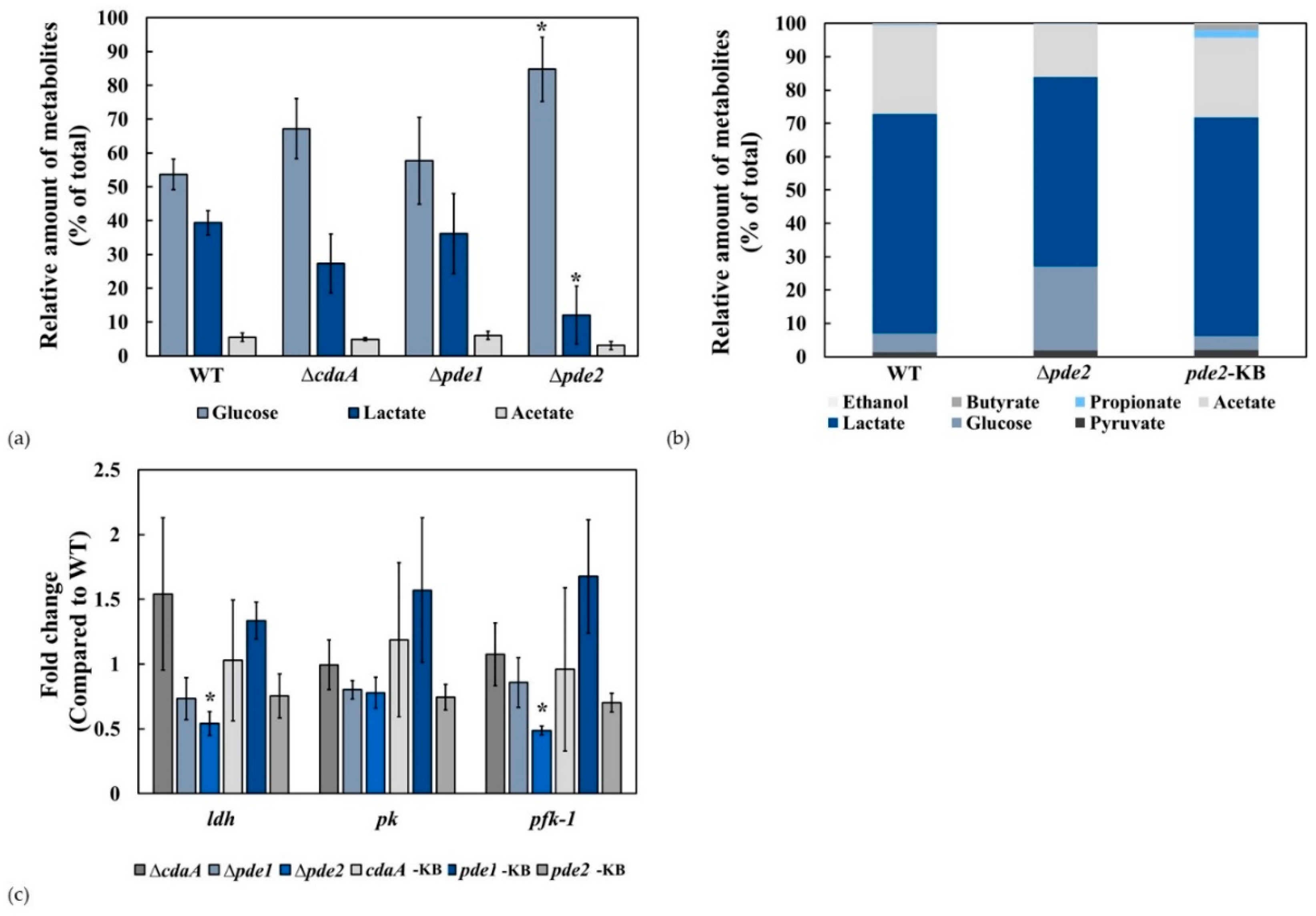
3.5. The ∆pde2 Mutant Displayed Reduced Rate of Glucose Metabolism
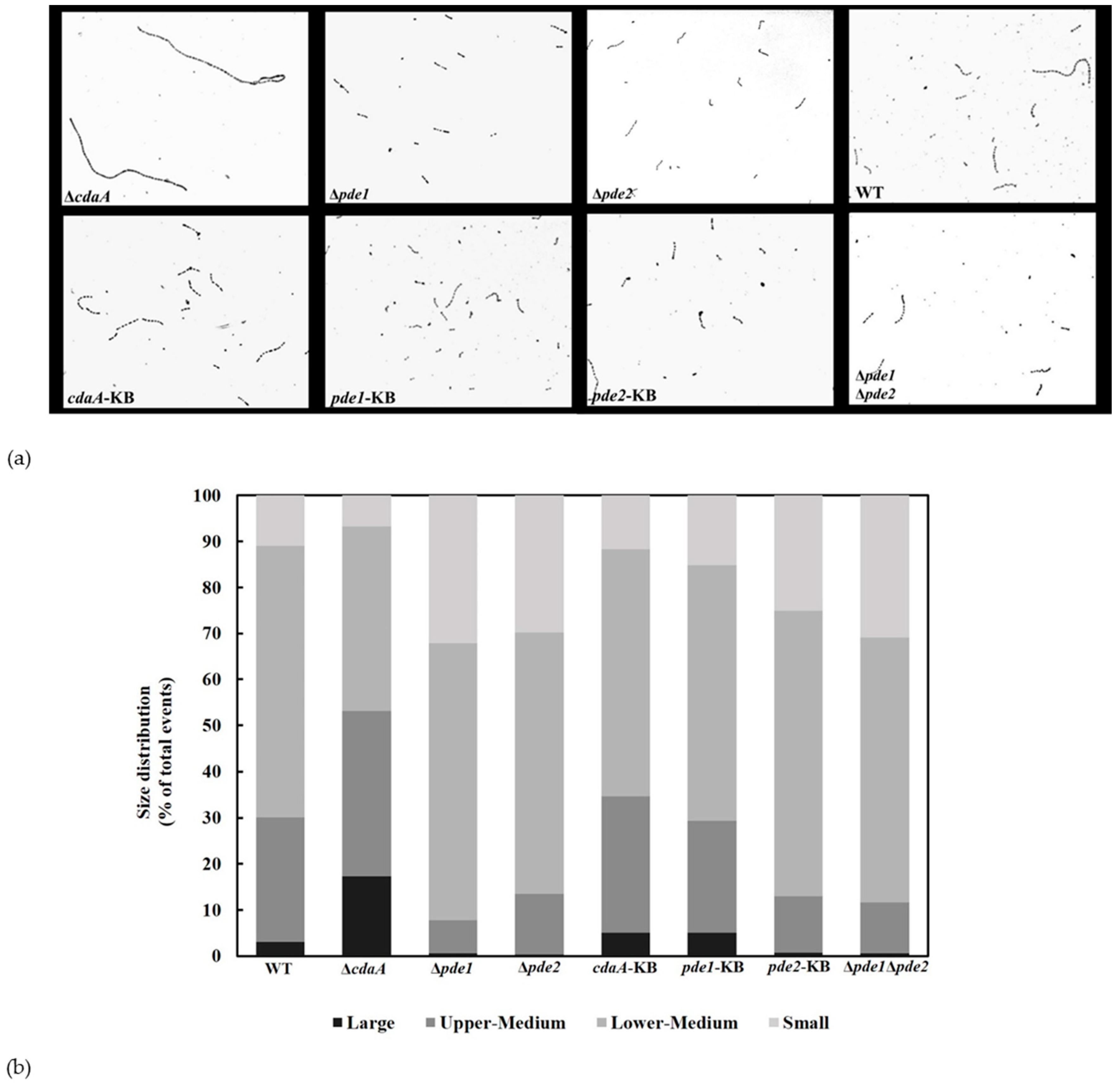
3.6. Interference with c-di-AMP Signaling Affects Chain Length
3.7. Transformation Efficiency Was not Affected by Disruption of the c-di-AMP Signalling System
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Witte, G.; Hartung, S.; Büttner, K.; Hopfner, K.-P. Structural Biochemistry of a Bacterial Checkpoint Protein Reveals Diadenylate Cyclase Activity Regulated by DNA Recombination Intermediates. Mol. Cell 2008, 30, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H.; Kang, S.O. Streptococcus pyogenes c-di-AMP Phosphodiesterase, GdpP, Influences SpeB Processing and Virulence. PLoS ONE 2013, 8, e69425. [Google Scholar] [CrossRef] [PubMed]
- Fahmi, T.; Faozia, S.; Port, G.C.; Cho, K.H. The Second Messenger c-di-AMP Regulates Diverse Cellular Pathways Involved in Stress Response, Biofilm Formation, Cell Wall Homeostasis, SpeB Expression, and Virulence in Streptococcus pyogenes. Infect. Immun. 2019, 87, e00147-19. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, R.M.; Abbott, J.; Burhenne, H.; Kaever, V.; Gründling, A. c-di-AMP Is a New Second Messenger in Staphylococcus aureus with a Role in Controlling Cell Size and Envelope Stress. PLoS Pathog. 2011, 7, e1002217. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Yang, J.; Zarrella, T.M.; Zhang, Y.; Metzger, D.W.; Bai, G. Cyclic Di-AMP Impairs Potassium Uptake Mediated by a Cyclic Di-AMP Binding Protein in Streptococcus pneumoniae. J. Bacteriol. 2013, 196, 614–623. [Google Scholar] [CrossRef]
- Whiteley, A.T.; Garelis, N.E.; Peterson, B.N.; Choi, P.H.; Tong, L.; Woodward, J.J.; Portnoy, D.A. c-di-AMP modulates Listeria monocytogenes central metabolism to regulate growth, antibiotic resistance and osmoregulation. Mol. Microbiol. 2017, 104, 212–233. [Google Scholar] [CrossRef]
- Zarrella, T.M.; Yang, J.; Metzger, D.W.; Bai, G. Bacterial Second Messenger Cyclic di-AMP Modulates the Competence State in Streptococcus pneumoniae. J. Bacteriol. 2019, 202, e00691-19. [Google Scholar] [CrossRef]
- Woodward, J.J.; Iavarone, A.T.; Portnoy, D.A. c-di-AMP Secreted by Intracellular Listeria monocytogenes Activates a Host Type I Interferon Response. Science 2010, 328, 1703–1705. [Google Scholar] [CrossRef]
- Barker, J.R.; Koestler, B.J.; Carpenter, V.K.; Burdette, D.L.; Waters, C.M.; Vance, R.E.; Valdivia, R. STING-Dependent Recognition of Cyclic di-AMP Mediates Type I Interferon Responses during Chlamydia trachomatis Infection. mBio 2013, 4, e00018-13. [Google Scholar] [CrossRef]
- McFarland, A.P.; Luo, S.; Qadri, F.A.; Zuck, M.; Thayer, E.F.; Goo, Y.A.; Hybiske, K.; Tong, L.; Woodward, J.J. Sensing of Bacterial Cyclic Dinucleotides by the Oxidoreductase RECON Promotes NF-kappaB Activation and Shapes a Proinflammatory Antibacterial State. Immunity 2017, 46, 433–445. [Google Scholar] [CrossRef]
- Parvatiyar, K.; Zhang, Z.; Teles, R.M.B.; Ouyang, S.; Jiang, Y.; Iyer, S.S.; Zaver, S.A.; Schenk, M.; Zeng, S.; Zhong, W.; et al. The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nat. Immunol. 2012, 13, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Wang, S.; Xiong, Z.; Zhu, X.; Ye, B.; Du, Y.; Meng, S.; Qu, Y.; Liu, J.; Gao, G.; et al. The ER membrane adaptor ERAdP senses the bacterial second messenger c-di-AMP and initiates anti-bacterial immunity. Nat. Immunol. 2018, 19, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Andrade, W.A.; Firon, A.; Schmidt, T.; Hornung, V.; Fitzgerald, K.A.; Kurt-Jones, E.A.; Trieu-Cuot, P.; Golenbock, D.T.; Kaminski, P.A. Group B Streptococcus Degrades Cyclic-di-AMP to Modulate STING-Dependent Type I Interferon Production. Cell Host Microbe 2016, 20, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, G.; Liang, L.; Lei, C.; Sun, X. Increased Excess Intracellular Cyclic di-AMP Levels Impair Growth and Virulence of Bacillus anthracis. J. Bacteriol. 2020, 202, 32071095. [Google Scholar] [CrossRef]
- Bai, Y.; Yang, J.; Eisele, L.E.; Underwood, A.J.; Koestler, B.J.; Waters, C.M.; Metzger, D.W.; Bai, G. Two DHH Subfamily 1 Proteins in Streptococcus pneumoniae Possess Cyclic Di-AMP Phosphodiesterase Activity and Affect Bacterial Growth and Virulence. J. Bacteriol. 2013, 195, 5123–5132. [Google Scholar] [CrossRef]
- Huynh, T.N.; Luo, S.; Pensinger, D.A.; Sauer, J.-D.; Tong, L.; Woodward, J.J. An HD-domain phosphodiesterase mediates cooperative hydrolysis of c-di-AMP to affect bacterial growth and virulence. Proc. Natl. Acad. Sci. USA 2015, 112, E747–E756. [Google Scholar] [CrossRef]
- Du, B.; Ji, W.; An, H.; Shi, Y.; Huang, Q.; Cheng, Y.; Fu, Q.; Wang, H.; Yan, Y.; Sun, J. Functional analysis of c-di-AMP phosphodiesterase, GdpP, in Streptococcus suis serotype 2. Microbiol. Res. 2014, 169, 749–758. [Google Scholar] [CrossRef]
- Kamegaya, T.; Kuroda, K.; Hayakawa, Y. Identification of a Streptococcus pyogenes SF370 gene involved in production of c-di-AMP. Nagoya J. Med. Sci. 2011, 73, 49–57. [Google Scholar]
- Bai, Y.; Yang, J.; Zhou, X.; Ding, X.; Eisele, L.E.; Bai, G. Mycobacterium tuberculosis Rv3586 (DacA) Is a Diadenylate Cyclase That Converts ATP or ADP into c-di-AMP. PLoS ONE 2012, 7, e35206. [Google Scholar] [CrossRef]
- Römling, U. Great Times for Small Molecules: C-di-AMP, a Second Messenger Candidate in Bacteria and Archaea. Sci. Signal. 2008, 1, pe39. [Google Scholar] [CrossRef]
- Mehne, F.M.P.; Gunka, K.; Eilers, H.; Herzberg, C.; Kaever, V.; Stulke, J. Cyclic di-AMP homeostasis in bacillus subtilis: Both lack and high level accumulation of the nucleotide are detrimental for cell growth. J. Biol. Chem. 2013, 288, 2004–2017. [Google Scholar] [CrossRef] [PubMed]
- Kellenberger, C.A.; Chen, C.; Whiteley, A.T.; Portnoy, D.A.; Hammond, M.C. RNA-Based Fluorescent Biosensors for Live Cell Imaging of Second Messenger Cyclic di-AMP. J. Am. Chem. Soc. 2015, 137, 6432–6435. [Google Scholar] [CrossRef] [PubMed]
- Blötz, C.; Treffon, K.; Kaever, V.; Schwede, F.; Hammer, E.; Stülke, J. Identification of the Components Involved in Cyclic Di-AMP Signaling in Mycoplasma pneumoniae. Front. Microbiol. 2017, 8, 1328. [Google Scholar] [CrossRef] [PubMed]
- Rao, F.; See, R.Y.; Zhang, D.; Toh, D.C.; Ji, Q.; Liang, Z.-X. YybT is a signaling protein that contains a cyclic dinucleotide phosphodiesterase domain and a GGDEF domain with ATPase activity. J. Biol. Chem. 2010, 285, 473–482. [Google Scholar] [CrossRef]
- Latoscha, A.; Drexler, D.J.; Al-Bassam, M.M.; Bandera, A.M.; Kaever, V.; Findlay, K.C.; Witte, G.; Tschowri, N. c-di-AMP hydrolysis by the phosphodiesterase AtaC promotes differentiation of multicellular bacteria. Proc. Natl. Acad. Sci. USA 2020, 117, 7392–7400. [Google Scholar] [CrossRef]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the Normal Bacterial Flora of the Oral Cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef]
- Sulyanto, R.M.; Thompson, Z.A.; Beall, C.J.; Leys, E.J.; Griffen, A.L. The Predominant Oral Microbiota Is Acquired Early in an Organized Pattern. Sci. Rep. 2019, 9, 10550. [Google Scholar] [CrossRef]
- Shelburne, S.A.; Sahasrabhojane, P.; Saldaña, M.; Yao, H.; Su, X.; Horstmann, N.; Thompson, E.; Flores, A.R. Streptococcus mitis Strains Causing Severe Clinical Disease in Cancer Patients. Emerg. Infect. Dis. 2014, 20, 762–771. [Google Scholar] [CrossRef]
- Basaranoglu, S.T.; Ozsurekci, Y.; Aykac, K.; Aycan, A.E.; Bıcakcigil, A.; Altun, B.; Sancak, B.; Cengiz, A.B.; Kara, A.; Ceyhan, M. Streptococcus mitis/oralis Causing Blood Stream Infections in Pediatric Patients. Jpn. J. Infect. Dis. 2019, 72, 1–6. [Google Scholar] [CrossRef]
- Marron, A.; Carratalà, J.; Gonzalez-Barca, E.; Fernández-Sevilla, A.; Alcaide, F.; Gudiol, F. Serious Complications of Bacteremia Caused by Viridans Streptococci in Neutropenic Patients with Cancer. Clin. Infect. Dis. 2000, 31, 1126–1130. [Google Scholar] [CrossRef]
- Kilian, M.; Poulsen, K.; Blomqvist, T.; Havarstein, L.S.; Bek-Thomsen, M.; Tettelin, H.; Sørensen, U.B.S. Evolution of Streptococcus pneumoniae and Its Close Commensal Relatives. PLoS ONE 2008, 3, e2683. [Google Scholar] [CrossRef] [PubMed]
- Kilian, M.; Riley, D.R.; Jensen, A.; Brüggemann, H.; Tettelin, H. Parallel Evolution of Streptococcus pneumoniae and Streptococcus mitis to Pathogenic and Mutualistic Lifestyles. mBio 2014, 5, e01490-14. [Google Scholar] [CrossRef] [PubMed]
- Salvadori, G.; Junges, R.; Morrison, D.A.; Petersen, F.C. Overcoming the Barrier of Low Efficiency during Genetic Transformation of Streptococcus mitis. Front. Microbiol. 2016, 7, 1009. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2014, 43, D257–D260. [Google Scholar] [CrossRef]
- Stevens, K.; Chang, D.; Zwack, E.E.; Sebert, M.E. Competence in Streptococcus pneumoniae Is Regulated by the Rate of Ribosomal Decoding Errors. mBio 2011, 2, e0007111. [Google Scholar] [CrossRef]
- Assev, S.; Stig, S.; Scheie, A.A. Cariogenic traits in xylitol-resistant and xylitol-sensitive mutans streptococci. Oral Microbiol. Immunol. 2002, 17, 95–99. [Google Scholar] [CrossRef]
- Burhenne, H.; Kaever, V. Quantification of Cyclic Dinucleotides by Reversed-Phase LC-MS/MS; Springer Science and Business Media LLC: Berlin, Germany, 2013; Volume 1016, pp. 27–37. [Google Scholar]
- Taylor, B.L.; Zhulin, I.B. PAS Domains: Internal Sensors of Oxygen, Redox Potential, and Light. Microbiol. Mol. Biol. Rev. 1999, 63, 479–506. [Google Scholar] [CrossRef]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Römling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef]
- Drexler, D.J.; Müller-Nurasyid, M.; Rojas-Cordova, C.A.; Bandera, A.M.; Witte, G. Structural and Biophysical Analysis of the Soluble DHH/DHHA1-Type Phosphodiesterase TM1595 from Thermotoga maritima. Structure 2017, 25, 1887.e4–1897.e4. [Google Scholar] [CrossRef]
- Tosi, T.; Hoshiga, F.; Millership, C.; Singh, R.; Eldrid, C.; Patin, D.; Mengin-Lecreulx, D.; Thalassinos, K.; Freemont, P.S.; Gründling, A. Inhibition of the Staphylococcus aureus c-di-AMP cyclase DacA by direct interaction with the phosphoglucosamine mutase GlmM. PLoS Pathog. 2019, 15, e1007537. [Google Scholar] [CrossRef]
- Whiteley, A.T.; Pollock, A.J.; Portnoy, D.A. The PAMP c-di-AMP Is Essential for Listeria monocytogenes Growth in Rich but Not Minimal Media due to a Toxic Increase in (p)ppGpp. Cell Host Microbe 2015, 18, 132. [Google Scholar] [CrossRef]
- Zeden, M.S.; Schuster, C.F.; Bowman, L.; Zhong, Q.; Williams, H.D.; Gründling, A. Cyclic di-adenosine monophosphate (c-di-AMP) is required for osmotic regulation inStaphylococcus aureusbut dispensable for viability in anaerobic conditions. J. Biol. Chem. 2018, 293, 3180–3200. [Google Scholar] [CrossRef] [PubMed]
- Devaux, L.; Sleiman, D.; Mazzuoli, M.-V.; Gominet, M.; Lanotte, P.; Trieu-Cuot, P.; Kaminski, P.A.; Firon, A. Cyclic di-AMP regulation of osmotic homeostasis is essential in Group B Streptococcus. PLoS Genet. 2018, 14, e1007342. [Google Scholar] [CrossRef]
- Peng, X.; Michalek, S.; Wu, H. Effects of diadenylate cyclase deficiency on synthesis of extracellular polysaccharide matrix of Streptococcus mutans revisit. Environ. Microbiol. 2016, 18, 3612–3619. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zheng, X.; Zhou, X.; Zeng, J.; Ren, Z.; Xu, X.; Cheng, L.; Li, M.; Li, J.; Li, Y. Regulation of oxidative response and extracellular polysaccharide synthesis by a diadenylate cyclase inStreptococcus mutans. Environ. Microbiol. 2015, 18, 904–922. [Google Scholar] [CrossRef] [PubMed]
- Teh, W.K.; Dramsi, S.; Tolker-Nielsen, T.; Yang, L.; Givskov, M. Increased Intracellular Cyclic di-AMP Levels Sensitize Streptococcus gallolyticus subsp. gallolyticus to Osmotic Stress and Reduce Biofilm Formation and Adherence on Intestinal Cells. J. Bacteriol. 2019, 201, e00597-18. [Google Scholar] [CrossRef]
- Du, B.; Sun, J.H. Diadenylate cyclase evaluation of ssDacA (SSU98_1483) in Streptococcus suis serotype 2. Genet. Mol. Res. 2015, 14, 6917–6924. [Google Scholar] [CrossRef]
- Heidemann, J.L.; Neumann, P.; Dickmanns, A.; Ficner, R. Crystal structures of the c-di-AMP–synthesizing enzyme CdaA. J. Biol. Chem. 2019, 294, 10463–10470. [Google Scholar] [CrossRef]
- Konno, H.; Yoshida, Y.; Nagano, K.; Takebe, J.; Hasegawa, Y. Biological and Biochemical Roles of Two Distinct Cyclic Dimeric Adenosine 3′,5′-Monophosphate- Associated Phosphodiesterases in Streptococcus mutans. Front. Microbiol. 2018, 9, 2347. [Google Scholar] [CrossRef]
- Wang, X.; Davlieva, M.; Reyes, J.; Panesso, D.; Arias, C.A.; Shamoo, Y. A Novel Phosphodiesterase of the GdpP Family Modulates Cyclic di-AMP Levels in Response to Cell Membrane Stress in Daptomycin-Resistant Enterococci. Antimicrob. Agents Chemother. 2017, 61, e01422-16. [Google Scholar] [CrossRef]
- Wang, F.; He, Q.; Su, K.; Wei, T.; Xu, S.; Gu, L. Structural and biochemical characterization of the catalytic domains of GdpP reveals a unified hydrolysis mechanism for the DHH/DHHA1 phosphodiesterase. Biochem. J. 2018, 475, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, K.; Sabareesh, V.; Singh, N.; Saigal, K.; Mechold, U.; Sinha, K.M. Two-Step Synthesis and Hydrolysis of Cyclic di-AMP in Mycobacterium tuberculosis. PLoS ONE 2014, 9, e86096. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhang, J.-J.; Fang, X.; Lawlis, G.B.; Troxell, B.; Zhou, Y.; Gomelsky, M.; Lou, Y.; Yang, X.F. DhhP, a Cyclic di-AMP Phosphodiesterase of Borrelia burgdorferi, Is Essential for Cell Growth and Virulence. Infect. Immun. 2014, 82, 1840–1849. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bai, Y.; Zhang, Y.; Gabrielle, V.D.; Jin, L.; Bai, G. Deletion of the cyclic di-AMP phosphodiesterase gene (cnpB) in Mycobacterium tuberculosis leads to reduced virulence in a mouse model of infection. Mol. Microbiol. 2014, 93, 65–79. [Google Scholar] [CrossRef]
- Kuipers, K.; Gallay, C.; Martínek, V.; Rohde, M.; Martínková, M.; Van Der Beek, S.L.; Jong, W.S.P.; Venselaar, H.; Zomer, A.; Bootsma, H.; et al. Highly conserved nucleotide phosphatase essential for membrane lipid homeostasis inStreptococcus pneumoniae. Mol. Microbiol. 2016, 101, 12–26. [Google Scholar] [CrossRef]
- Tang, Q.; Luo, Y.; Zheng, C.; Yin, K.; Ali, M.K.; Li, X.; He, J. Functional Analysis of a c-di-AMP-specific Phosphodiesterase MsPDE from Mycobacterium smegmatis. Int. J. Biol. Sci. 2015, 11, 813–824. [Google Scholar] [CrossRef]
- Bowman, L.; Zeden, M.S.; Schuster, C.F.; Kaever, V.; Gründling, A. New Insights into the Cyclic Di-adenosine Monophosphate (c-di-AMP) Degradation Pathway and the Requirement of the Cyclic Dinucleotide for Acid Stress Resistance inStaphylococcus aureus. J. Biol. Chem. 2016, 291, 26970–26986. [Google Scholar] [CrossRef]
- Sureka, K.; Choi, P.H.; Precit, M.; Delince, M.; Pensinger, D.A.; Huynh, T.N.; Jurado, A.R.; Goo, Y.A.; Sadílek, M.; Iavarone, A.T.; et al. The cyclic dinucleotide c-di-AMP is an allosteric regulator of metabolic enzyme function. Cell 2014, 158, 1389–1401. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Origin | Reference |
|---|---|---|---|
| S. mitis CCUG 31611 | Type strain Streptococcus mitis, corresponding to NCTC 12261 and ATCC 49456 | CCUG | |
| MI009 | ∆srtA::erm; ErmR,. Used to prepare an amplicon of the erythromycin resistance cassette and flanking regions used in transformation efficiency assays | S. mitis CCUG 31611 | [33] |
| S. mitis ΔcdaA | Markerless in-frame deletion of SM12261_1351 | S. mitis CCUG 31611 | This study |
| S. mitis Δpde1 | Markerless in-frame deletion of SM12261_1779 | S. mitis CCUG 31611 | This study |
| S. mitis Δpde2 | Markerless in-frame deletion of SM12261_1122 | S. mitis CCUG 31611 | This study |
| S. mitis Δpde1Δpde2 | Markerless in-frame deletion of SM12261_1779 and SM12261_1122 | S. mitis Δpde2 | This study |
| S. mitis cdaA-KB | SM12261_1351 re-introduced into the original locus | S. mitis ΔcdaA | This study |
| S. mitis pde1-KB | SM12261_1779 reintroduced into the original locus | S. mitis Δpde1 | This study |
| S. mitis pde2-KB | SM12261_1122 re-introduced into the original locus | S. mitis Δpde2 | This study |
| E. coli DH5α | Strain used for cloning | - | Thermo Fisher Scientific |
| E. coli BL21 (DE3) | Strain used for expression of recombinant proteins. | - | Thermo Fisher Scientific |
| pET28a(+) | Expression vector with IPTG-inducible T7-promoter; KanR | Novagen | |
| pET28a(+)KpnI-mut | pET28a(+)-vector with the NcoI restriction site mutated to KpnI; KanR | pET28a(+) | This study |
| pET28a(+)KpnI-mut_CdaA-N-His | pET28a(+)KpnI-mut-vector carrying the full-length S. mitis cdaA ORF; KanR | pET28(+)KpnI-mut | This study |
| pET28a(+)KpnI-mut_CdaA-T-N-His | pET28a(+)KpnI-mut-vector carrying the S. mitis cdaA including aa 94-285 with an N-terminal 6x His-tag; KanR | pET28(+)KpnI-mut | This study |
| pET28a(+)KpnI-mut_CdaA-T2-N-His | pET28a(+)KpnI-mut-vector carrying the S. mitis cdaA including aa 103–285 with an N-terminal 6x His-tag; KanR | pET28(+)KpnI-mut | This study |
| pET28a(+)KpnI-mut_Pde1-N-His | pET28a(+)KpnI-mut-vector carrying the full-length S. mitis pde1 with an N-terminal 6x His-tag; KanR | pET28(+)KpnI-mut | This study |
| pET28a(+)KpnI-mut_Pde1-T-N-His | pET28a(+)KpnI-mut-vector carrying the S. mitis pde1 ORF including aa 53–657 with an N-terminal 6x His-tag; KanR | pET28a(+)KpnI-mut_PDE1-N-His | This study |
| pET28a(+)KpnI-mut_Pde2-N-His | pET28a(+)KpnI-mut-vector carrying the full-length S. mitis pde2 with an N-terminal 6x His-tag; KanR | pET28(+)KpnI-mut | This study |
| Enzyme | Substrate | Product | [Enzyme] (μM) | Km (μM) | Vmax (μM s−1) | Kcat (s−1) | kcat/Km (μM−1 s−1) |
|---|---|---|---|---|---|---|---|
| CdaA103–285 | ATP | c-di-AMP | 10 | 821.3 ± 10.5 | 0.280 ± 0.046 | 28 × 10−3 ± 4.6 × 10−3 | 3.41 × 10−5 |
| Pde153–657 | c-di-AMP | pApA | 2.5 | 241.3 ± 11.6 | 10.35 ± 4.98 | 4.14 ± 1.99 | 17 × 10−3 |
| pApA | AMP | 5 | 152.4 ± 03.6 | 12.47 ± 3.06 | 2.49 ± 0.61 | 16.4 × 10−3 | |
| Pde2 | c-di-AMP | AMP | 10 | 148.5 ± 34.8 | 0.78 ± 0.05 | 0.078 ± 0.005 | 5.25 × 10−4 |
| pApA | AMP | 0.05 | 877 ± 312 | 176.4 ± 31.2 | 3527 ± 623 | 4.02 |
| Strain | Generation Time (min) (Standard Error of the Mean) |
|---|---|
| WT | 39.6 ± 0.5 |
| ∆cdaA | 38.7 ± 2.3 |
| cdaA-KB | 37.3 ± 1.0 |
| ∆pde1 | 42.0 ± 0 |
| pde1-KB | 39.4 ± 1.1 |
| ∆pde2 | 54.8 ± 0.6 |
| pde2-kb | 34.2 ± 0.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rørvik, G.H.; Liskiewicz, K.A.; Kryuchkov, F.; Naemi, A.-O.; Aasheim, H.-C.; Petersen, F.C.; Küntziger, T.M.; Simm, R. Cyclic Di-adenosine Monophosphate Regulates Metabolism and Growth in the Oral Commensal Streptococcus mitis. Microorganisms 2020, 8, 1269. https://doi.org/10.3390/microorganisms8091269
Rørvik GH, Liskiewicz KA, Kryuchkov F, Naemi A-O, Aasheim H-C, Petersen FC, Küntziger TM, Simm R. Cyclic Di-adenosine Monophosphate Regulates Metabolism and Growth in the Oral Commensal Streptococcus mitis. Microorganisms. 2020; 8(9):1269. https://doi.org/10.3390/microorganisms8091269
Chicago/Turabian StyleRørvik, Gro Herredsvela, Krystyna Anna Liskiewicz, Fedor Kryuchkov, Ali-Oddin Naemi, Hans-Christian Aasheim, Fernanda C. Petersen, Thomas M. Küntziger, and Roger Simm. 2020. "Cyclic Di-adenosine Monophosphate Regulates Metabolism and Growth in the Oral Commensal Streptococcus mitis" Microorganisms 8, no. 9: 1269. https://doi.org/10.3390/microorganisms8091269
APA StyleRørvik, G. H., Liskiewicz, K. A., Kryuchkov, F., Naemi, A.-O., Aasheim, H.-C., Petersen, F. C., Küntziger, T. M., & Simm, R. (2020). Cyclic Di-adenosine Monophosphate Regulates Metabolism and Growth in the Oral Commensal Streptococcus mitis. Microorganisms, 8(9), 1269. https://doi.org/10.3390/microorganisms8091269