Heterogeneity of Molecular Characteristics among Staphylococcus argenteus Clinical Isolates (ST2250, ST2793, ST1223, and ST2198) in Northern Taiwan

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Identification of S. argenteus by Detection of MALDI-TOF MS, MLST Typing, and Detection of CrtM
2.2. Pulsed-Field Gel Electrophoresis and Spa Typing
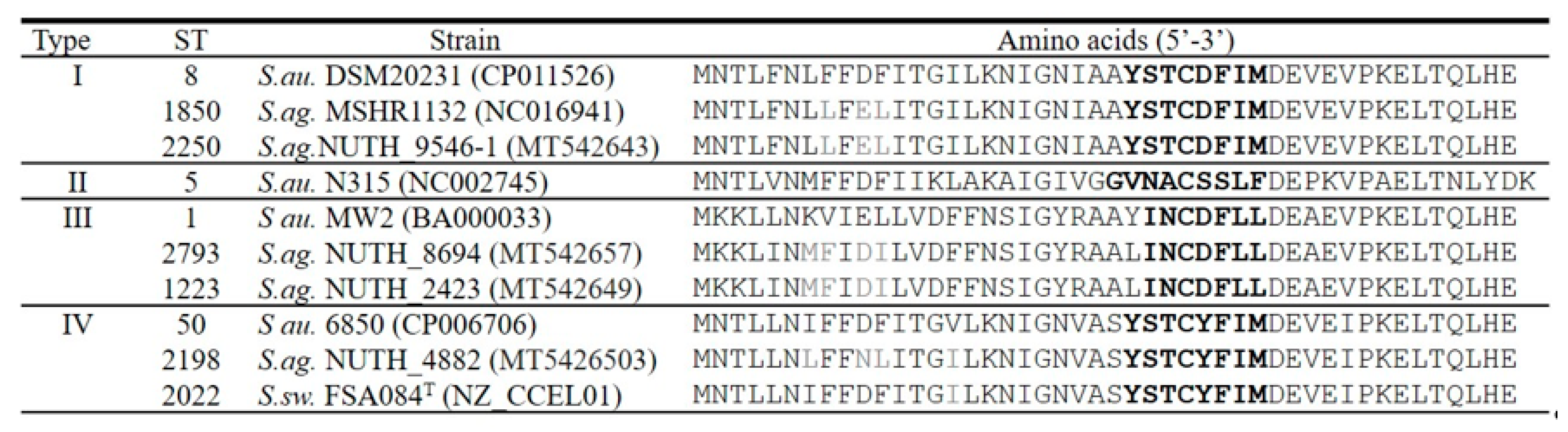
2.3. Coa, DnaJ, GroEL Gene and Spacer Sequencing, and AgrD Type
2.4. Detection of Virulence Factors
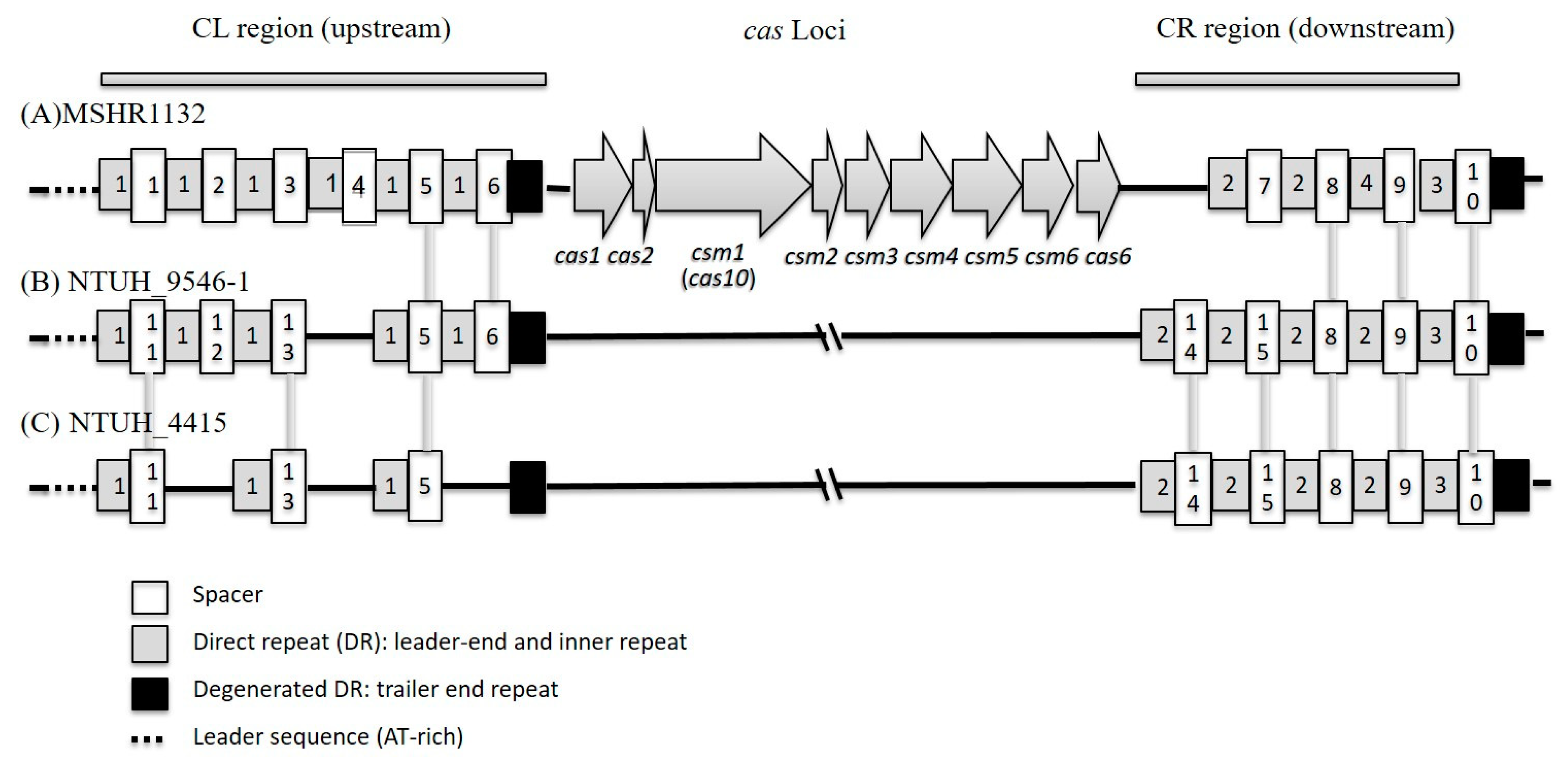
2.5. Sequencing of CRISPR/cas Loci of S. argenteus ST2250
2.6. Phylogenetic Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
3.1. Identification of S. argenteus by MALDI-TOF MS, MLST Typing and Lack of CrtM
3.2. Antimicrobial Susceptibility and Biochemical Characteristics
3.3. Genotyping by Pulsed-Field Gel Electrophoresis (PFGE) and Spa Types
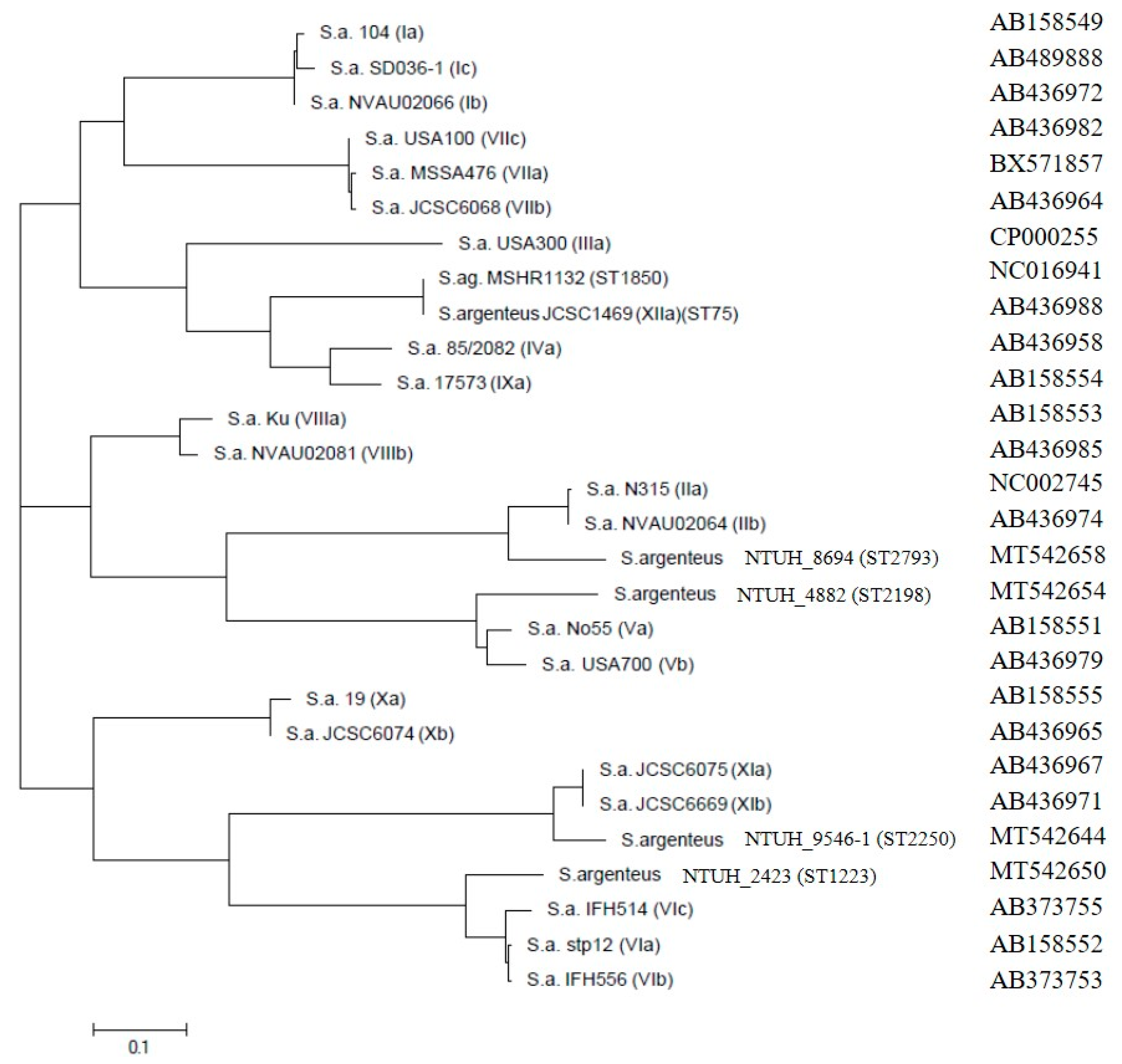
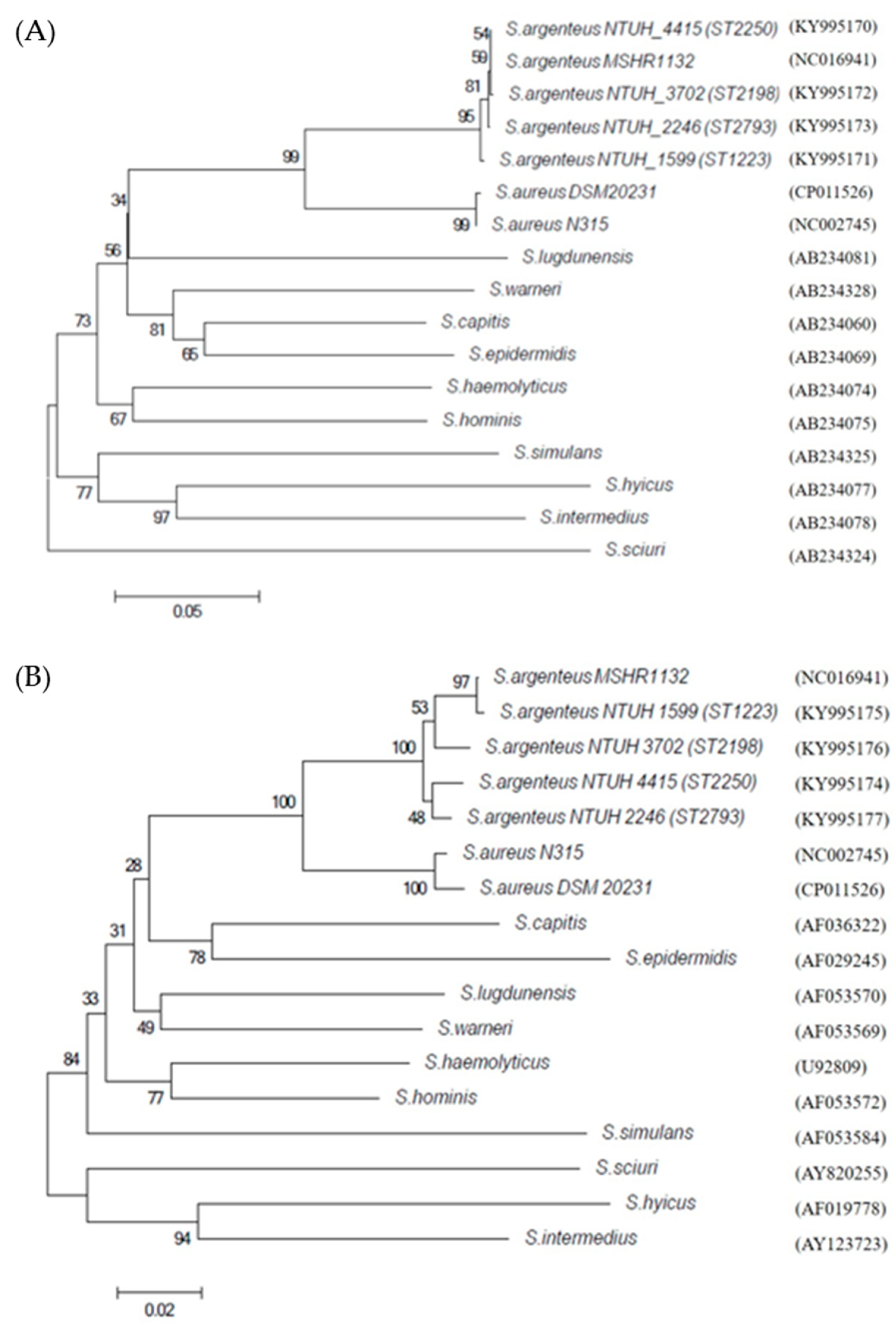
3.4. Phylogenetic Trees of Coa, DnaJ and GroESL and Agr Type
3.5. Examination of Virulence Factors
3.6. Polymorphism of CRISPR/Cas System in ST2250 Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Holt, D.C.; Holden, M.T.; Tong, S.Y.; Castillo-Ramirez, S.; Clarke, L.; Quail, M.A.; Currie, B.J.; Parkhill, J.; Bentley, S.D.; Feil, E.J.; et al. A very early-branching Staphylococcus aureus lineage lacking the carotenoid pigment staphyloxanthin. Genome Biol. Evol. 2011, 3, 881–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.; Schaumburg, F.; Ellington, M.J.; Corander, J.; Pichon, B.; Leendertz, F.; Bentley, S.D.; Parkhill, J.; Holt, D.C.; Peters, G.; et al. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: The non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Argudin, M.A.; Dodemont, M.; Vandendriessche, S.; Rottiers, S.; Tribes, C.; Roisin, S.; de Mendonca, R.; Nonhoff, C.; Deplano, A.; Denis, O. Low occurrence of the new species Staphylococcus argenteus in a Staphylococcus aureus collection of human isolates from Belgium. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1017–1022. [Google Scholar] [CrossRef]
- Dupieux, C.; Blonde, R.; Bouchiat, C.; Meugnier, H.; Bes, M.; Laurent, S.; Vandenesch, F.; Laurent, F.; Tristan, A. Community-acquired infections due to Staphylococcus argenteus lineage isolates harbouring the Panton-Valentine leucocidin, France, 2014. Eurosurveillance 2015, 20, 21154. [Google Scholar] [CrossRef] [PubMed]
- Thaipadungpanit, J.; Amornchai, P.; Nickerson, E.K.; Wongsuvan, G.; Wuthiekanun, V.; Limmathurotsakul, D.; Peacock, S.J. Clinical and molecular epidemiology of Staphylococcus argenteus infections in Thailand. J. Clin. Microbiol. 2015, 53, 1005–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, T.; Shinjoh, M.; Ohara, H.; Kawai, T.; Kamimaki, I.; Mizushima, R.; Kamada, K.; Itakura, Y.; Iguchi, S.; Uzawa, Y.; et al. Purulent lymphadenitis caused by Staphylococcus argenteus, representing the first Japanese case of Staphylococcus argenteus (multilocus sequence type 2250) infection in a 12-year-old boy. J. Infect. Chemother. 2018, 24, 925–927. [Google Scholar] [CrossRef]
- Zhang, D.-F.; Xu, X.; Song, Q.; Bai, Y.; Zhang, Y.; Song, M.; Shi, C.; Shi, X. Identification of Staphylococcus argenteus in Eastern China based on a nonribosomal peptide synthetase (NRPS) gene. Future Microbiol. 2016, 11, 1113–1121. [Google Scholar] [CrossRef]
- Chen, S.Y.; Lee, H.; Wang, X.M.; Lee, T.F.; Liao, C.H.; Teng, L.J.; Hsueh, P.R. High mortality impact of Staphylococcus argenteus on patients with community-onset staphylococcal bacteraemia. Int. J. Antimicrob Agents 2018, 52, 747–753. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Lee, H.; Teng, S.-H.; Wang, X.-M.; Lee, T.-F.; Huang, Y.-C.; Liao, C.-H.; Teng, L.-J.; Hsueh, P.-R. Accurate differentiation of novel Staphylococcus argenteus from Staphylococcus aureus using MALDI-TOF MS. Future Microbiol. 2018, 13, 997–1006. [Google Scholar] [CrossRef]
- Hansen, T.A.; Bartels, M.D.; Hogh, S.V.; Dons, L.E.; Pedersen, M.; Jensen, T.G.; Kemp, M.; Skov, M.N.; Gumpert, H.; Worning, P.; et al. Whole Genome Sequencing of Danish Staphylococcus argenteus Reveals a Genetically Diverse Collection with Clear Separation from Staphylococcus aureus. Front. Microbiol. 2017, 8, 1512. [Google Scholar] [CrossRef]
- Moradigaravand, D.; Jamrozy, D.; Mostowy, R.; Anderson, A.; Nickerson, E.K.; Thaipadungpanit, J.; Wuthiekanun, V.; Limmathurotsakul, D.; Tandhavanant, S.; Wikraiphat, C.; et al. Evolution of the Staphylococcus argenteus ST2250 Clone in Northeastern Thailand Is Linked with the Acquisition of Livestock-Associated Staphylococcal Genes. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.F.; Zhi, X.Y.; Zhang, J.; Paoli, G.C.; Cui, Y.; Shi, C.; Shi, X. Preliminary comparative genomics revealed pathogenic potential and international spread of Staphylococcus argenteus. BMC Genom. 2017, 18, 808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giske, C.G.; Dyrkell, F.; Arnellos, D.; Vestberg, N.; Fang, H. Transmission events and antimicrobial susceptibilities of methicillin-resistant Staphylococcus argenteus in Stockholm. Clin. Microbiol. Infect. 2019, 25, 1289.e5–1289.e8. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Sumi, A.; Takahashi, S.; Ike, M.; Ito, M.; Habadera, S.; Kobayashi, N. Molecular Epidemiological Characterization of Staphylococcus argenteus Clinical Isolates in Japan: Identification of Three Clones (ST1223, ST2198, and ST2550) and a Novel Staphylocoagulase Genotype XV. Microorganisms 2019, 7, 389. [Google Scholar] [CrossRef] [Green Version]
- Chantratita, N.; Wikraiphat, C.; Tandhavanant, S.; Wongsuvan, G.; Ariyaprasert, P.; Suntornsut, P.; Thaipadungpanit, J.; Teerawattanasook, N.; Jutrakul, Y.; Srisurat, N.; et al. Comparison of community-onset Staphylococcus argenteus and Staphylococcus aureus sepsis in Thailand: A prospective multicentre observational study. Clin. Microbiol. Infect. 2016, 22, 458.e11–458.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, M.S.; San, T.; San, N.; Oo, W.M.; Ko, P.M.; Thet, K.T.; Urushibara, N.; Kawaguchiya, M.; Sumi, A.; Kobayashi, N. Molecular characterization of Staphylococcus argenteus in Myanmar: Identification of novel genotypes/clusters in staphylocoagulase, protein A, alpha-haemolysin and other virulence factors. J. Med. Microbiol. 2019, 68, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; San, T.; Aye, M.M.; Mya, S.; Maw, W.W.; Zan, K.N.; Htut, W.H.W.; Kawaguchiya, M.; Urushibara, N.; Kobayashi, N. Prevalence and Genetic Characteristics of Staphylococcus aureus and Staphylococcus argenteus Isolates Harboring Panton-Valentine Leukocidin, Enterotoxins, and TSST-1 Genes from Food Handlers in Myanmar. Toxins 2017, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.; Wong, M.Y.; Tseng, Y.H.; Lin, C.L.; Tung, C.W.; Kao, C.C.; Huang, Y.K. Vascular access infection by Staphylococcus aureus from removed dialysis accesses. Microbiologyopen 2019, 8, e800. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.W.; Holt, D.C.; Lilliebridge, R.A.; Stephens, A.J.; Huygens, F.; Tong, S.Y.; Currie, B.J.; Giffard, P.M. Phylogenetically distinct Staphylococcus aureus lineage prevalent among indigenous communities in northern Australia. J. Clin. Microbiol. 2009, 47, 2295–2300. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Essex, A.; Buchanan, J.T.; Datta, V.; Hoffman, H.M.; Bastian, J.F.; Fierer, J.; Nizet, V. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 2005, 202, 209–215. [Google Scholar] [CrossRef]
- Koreen, L.; Ramaswamy, S.V.; Graviss, E.A.; Naidich, S.; Musser, J.M.; Kreiswirth, B.N. spa typing method for discriminating among Staphylococcus aureus isolates: Implications for use of a single marker to detect genetic micro-and macrovariation. J. Clin. Microbiol. 2004, 42, 792–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauschild, T.; Stepanovic, S. Identification of Staphylococcus spp. by PCR-restriction fragment length polymorphism analysis of dnaJ gene. J. Clin. Microbiol. 2008, 46, 3875–3879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, W.-C.; Tseng, S.-P.; Chen, H.-J.; Tsai, J.-C.; Chang, C.-H.; Lee, T.-F.; Hsueh, P.-R.; Teng, L.-J. Use of groESL as a target for identification of Abiotrophia, Granulicatella, and Gemella species. J. Clin. Microbiol. 2010, 48, 3532–3538. [Google Scholar] [CrossRef] [Green Version]
- Strommenger, B.; Cuny, C.; Werner, G.; Witte, W. Obvious lack of association between dynamics of epidemic methicillin-resistant Staphylococcus aureus in central Europe and agr specificity groups. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Kobayashi, N.; Nagashima, S.; Ishino, M.; Otokozawa, S.; Mise, K.; Sumi, A.; Tsutsumi, H.; Uehara, N.; Watanabe, N.; et al. Diversity of staphylocoagulase and identification of novel variants of staphylocoagulase gene in Staphylococcus aureus. Microbiol. Immunol. 2008, 52, 334–348. [Google Scholar] [CrossRef]
- Jarraud, S. Relationships between Staphylococcus aureus Genetic Background, Virulence Factors, agr Groups (Alleles), and Human Disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Peacock, S.J.; Moore, C.E.; Justice, A.; Kantzanou, M.; Story, L.; Mackie, K.; O’Neill, G.; Day, N.P. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect. Immun. 2002, 70, 4987–4996. [Google Scholar] [CrossRef] [Green Version]
- van Wamel, W.J.; Rooijakkers, S.H.; Ruyken, M.; van Kessel, K.P.; van Strijp, J.A. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Thoendel, M.; Kavanaugh, J.S.; Flack, C.E.; Horswill, A.R. Peptide signaling in the staphylococci. Chem. Rev. 2011, 111, 117–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Kubota, H.; Ono, H.K.; Kobayashi, M.; Murauchi, K.; Kato, R.; Hirai, A.; Sadamasu, K. Food poisoning outbreak in Tokyo, Japan caused by Staphylococcus argenteus. Int. J. Food Microbiol. 2017, 262, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Neyaz, L.; Karki, A.B.; Fakhr, M.K. The Whole-Genome Sequence of Plasmid-Bearing Staphylococcus argenteus Strain B3-25B from Retail Beef Liver Encodes the Type VII Secretion System and Several Virulence Factors. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Lia, Q.; Lia, Y.; Tang, Y.; Meng, C.; Ingmer, H.; Jiao, X. Prevalence and characterization of Staphylococcus aureus and Staphylococcus argenteus in chicken from retail markets in China. Food Control. 2019, 96, 158–164. [Google Scholar] [CrossRef]
- Pumipuntu, N.; Tunyong, W.; Chantratita, N.; Diraphat, P.; Pumirat, P.; Sookrung, N.; Chaicumpa, W.; Indrawattana, N. Staphylococcus spp. associated with subclinical bovine mastitis in central and northeast provinces of Thailand. PeerJ 2019, 7, e6587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, S.W.; Beres, S.B.; Olsen, R.J.; Musser, J.M. Absence of patient-to-patient intrahospital transmission of Staphylococcus aureus as determined by whole-genome sequencing. mBio 2014, 5, e01692-14. [Google Scholar] [CrossRef] [Green Version]
- Kwok, A.Y.; Chow, A.W. Phylogenetic study of Staphylococcus and Macrococcus species based on partial hsp60 gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.F.; Yang, X.Y.; Zhang, J.; Qin, X.; Huang, X.; Cui, Y.; Zhou, M.; Shi, C.; French, N.P.; Shi, X. Identification and characterization of two novel superantigens among Staphylococcus aureus complex. Int. J. Med. Microbiol. 2018, 308, 438–446. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Umeda, K.; Yonogi, S.; Nakamura, H.; Yamamoto, K.; Kumeda, Y.; Kawatsu, K. Staphylococcal food poisoning caused by Staphylococcus argenteus harboring staphylococcal enterotoxin genes. Int. J. Food Microbiol. 2018, 265, 23–29. [Google Scholar] [CrossRef]
- Schuster, D.; Rickmeyer, J.; Gajdiss, M.; Thye, T.; Lorenzen, S.; Reif, M.; Josten, M.; Szekat, C.; Melo, L.D.; Schmithausen, R.M.; et al. Differentiation of Staphylococcus argenteus (formerly: Staphylococcus aureus clonal complex 75) by mass spectrometry from S. aureus using the first strain isolated from a wild African great ape. Int. J. Med. Microbiol. 2017, 307, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Gao, C.H.; Zhu, J.; Zhao, L.; Wu, Q.; Li, M.; Sun, B. Identification and functional study of type III-A CRISPR-Cas systems in clinical isolates of Staphylococcus aureus. Int. J. Med. Microbiol. 2016, 306, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Mao, T.; Long, J.; Duan, G.; Yang, H. Commentary: Study the Features of 57 Confirmed CRISPR Loci in 38 Strains of Staphylococcus aureus. Front. Microbiol. 2019, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, Z.; Xu, Z. Study the Features of 57 Confirmed CRISPR Loci in 38 Strains of Staphylococcus aureus. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ST | No. of Isolates Each Year | |||||
|---|---|---|---|---|---|---|
| Spa Type | 2000 | 2005 | 2010 | 2011 | 2012 | Total (%) |
| ST2250 | 0 | 0 | 19 | 25 | 28 | 72 (75.0) |
| t5078 a | 0 | 0 | 15 | 18 | 22 | 55 (57.3) |
| t5787 | 0 | 0 | 0 | 1 | 0 | 1 (1.0) |
| t6675 | 0 | 0 | 0 | 2 | 3 | 5 (5.2) |
| t7960 | 0 | 0 | 1 | 1 | 1 | 3 (3.1) |
| t10900 | 0 | 0 | 0 | 0 | 1 | 1 (1.0) |
| t17928 | 0 | 0 | 0 | 2 | 0 | 2 (2.1) |
| Unknown b | 0 | 0 | 3 | 1 | 1 | 5 (5.3) |
| ST2793 | 1 | 2 | 4 | 4 | 1 | 12 (12.5) |
| Unknown c | 1 | 2 | 4 | 4 | 1 | 12 (12.5) |
| ST1223 | 0 | 3 | 5 | 1 | 1 | 10 (10.4) |
| t5142 | 0 | 2 | 2 | 0 | 0 | 4 (4.2) |
| t7463 | 0 | 1 | 0 | 0 | 0 | 1 (1.0) |
| t9791 | 0 | 0 | 3 | 0 | 1 | 4 (4.2) |
| t18323 | 0 | 0 | 0 | 1 | 0 | 1 (1.0) |
| ST2198 | 0 | 0 | 1 | 1 | 0 | 2 (2.1) |
| t9505 | 0 | 0 | 0 | 1 | 0 | 1 (1.0) |
| Unknown d | 0 | 0 | 1 | 0 | 0 | 1 (1.0) |
| Total (%) | 1 (1.0) | 5 (5.2) | 29 (30.2) | 31 (32.3) | 30 (31.3) | 96 (100) |
| ST Type | Spa Repeat Profile | No. of Isolates |
|---|---|---|
| ST2250 | 299-31-25-17-16-16-16-16 | 1 |
| ST2250 | 299-31-25-16-16-16-16-16 | 1 |
| ST2250 | 299-31-25-17-119-16-16-16-16 | 1 |
| ST2250 | 299-31-17-17-16-16-16-16-16-16-16 | 1 |
| ST2250 | 299-31-31-31-25-17-17-16-16-16-16 | 1 |
| ST2793 | 259-31-16-16-16-23-17-360-360-25 (t19483) | 8 |
| ST2793 | 259-31-16-16-23-307-360-360-25 | 1 |
| ST2793 | 259-31-307-16-23-17-360-360-25 | 1 |
| ST2793 | 259-25-25 | 1 |
| ST2793 | 259-25 | 1 |
| ST2198 | 259-23-23-17-17-17-23-23-23-17-17-16 | 1 |
| Strain | Spacer Length (bp) | Spacer Sequence (5′-3′) |
|---|---|---|
| S.ag. MSHR1132 (ST1850) (NC016941) | 71 | TACAGAACTTAATTCATAAATAAATTATTAAGAACAATAATCAAACA....TTAAAAAATGGAGGTTTATTATAT |
| S.ag. NTUH_9546-1 (ST2250) (MT542642) | 71 | -----------------------------------------------....------------------------ |
| S.ag. NTUH_8694 (ST2793) (MT542656) | 71 | --g---g----------------------------------------....------------------------ |
| S.ag. NTUH_2423 (ST1223) (MT542648) | 75 | ---------------------------aa------g-a---g-----caat-a--c------------------- |
| S.ag. NTUH_4882 (ST2198) (MT542652) | 71 | -----------------------------------------------....------------------------ |
| S.a. N315 (ST5) (NC002745) | 75 | --t-a--t-a--------g-------g-a------g-a---g---t-tgac-a--c--------------c--t- |
| S.a. DSM20231 (ST8) (CP011526) | 75 | ---------------------------aat-----g-a---g-----caac-a--c--------------c--t- |
| ST Type (No. of Isolates) | ST2250 (72) | ST2793 (12) | ST1223 (10) | ST2198 (2) |
|---|---|---|---|---|
| agr type b | I | III | III | IV |
| coa genotype b | XId | XVI | XV (serotype VI) | XIV |
| CRISPR | + (72) | NT | NT | NT |
| IEC type C | sak+, scn+ (60) (Type E) sak+ (1) scn+ (1) NT (10) | scn+ (11) NT(1) | scn+ (9) NT(1) | sak+, scn+, chp+ (1) scn+ (1) |
| hla & hld | + (72) | + (12) | + (10) | + (2) |
| cna | NT | + (12, type III) | NT | NT |
| bbp | NT | NT | NT | + (2) |
| seb | NT | NT | + (3) | + (1) |
| sec | NT | + (8) | NT | NT |
| egc cluster | NT | NT | + (10) | NT |
| Type | Sequence | Size(bp) | Note |
|---|---|---|---|
| DR1 | GATCGATAACTACCCCGAAGAATAGGGGACGAGAACX | 37 | upstream |
| DR2 | XATTCGATAACTACCCCGAAGAAGAGGGGACGAGAAC | 37 | downstream |
| DR3 | TATTCGATAACTACCCCGAAGAAAAGGGGACGAGAAC | 37 | downstream |
| DR4 | TATTCGATAACTACCCCGAAGAAGAGGAGACGAGAAC | 37 | downstream |
| DDR | TGATCGATAACTACCCCGAAGAATAGGGGACAGAGTG | 37 | Degenerated DR |
| DDR | TATTCGATAAATACCCCGGAGAACAGGGGGCGAAAAC | 37 | Degenerated DR |
| S1 | CTACTAAAAAGTTATATGTTTCAACAATTTCGTCA | 35 | upstream |
| S2 | GGTTTAAGTTTGTCATTATAATCAATCCTTTTTCTT | 36 | upstream |
| S3 | GATTAAAACGGTTTGCTTTATTTGCATTTAAAATAG | 36 | upstream |
| S4 | TTTTTCATAGTTAATCAATCCCTTTTCTTTTTT | 33 | upstream |
| S5 | TAAATCTTTGATTGCTCTTAGCTCTAGTTATGTAT | 35 | upstream |
| S6 | ACGCTGTAGTGAAGTATAGAAACGGCATGAGTACAAT | 37 | upstream |
| S7 | TTTTACTGTGTTTTTCATAATTAATCAATCCTTT | 34 | downstream |
| S8 | TGCCCACTTAATTAATTCATCTAGTCTCATTTCTT | 35 | downstream |
| S9 | CATCAACTGACTTTTTAACTGTTTTAGTGAATTCGTC | 37 | downstream |
| S10 | TTAAAGATCTCAACAATAGCGTCCCATATTTTCTG | 35 | downstream |
| S11 | TAATTGCATTATCAAATGTATATGCTGGATTCCAT | 35 | upstream |
| S12 | GTACTTAAGATTTCATCAACTTTCTTTTGTACT | 33 | upstream |
| S13 | CGAATTTTGATTCTTTGTTTGTAAATAATGCTCT | 34 | upstream |
| S14 | CTATAATAGTTACTGCTTTTGTAACCGTCCATAT | 34 | downstream |
| S15 | AAATGCTTATCCATTCTAATCATATTTTCAATTTGTTTA | 39 | downstream |
| S16 | TGCCCACTTAATTAATTCATCTAGTCTTATTTCTT | 35 | downstream similar to S8 |
| Isolate ID a (Accession No.) | Number of Spacers | Direct Repeat | |
|---|---|---|---|
| Upstream | Downstream | ||
| MSHR 1132 (ST1850) (NC016941) | 6 (S1~6) | 4 (S7~10) | DR1, 2, 3, 4 |
| TD13 (ST2854) (MH513583) | 5 (S11~13,5~6) | - | DR1 |
| TD162 (ST2250) (MF167423) | 5 (S11~13,5~6) | - a | DR1 |
| SH3 (ST2250) (PRJEB8900) | 5 (S11~13,5~6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 36 isolates (NTUH_9546-1) (MT542645) | 5 (S11~13,5~6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 9 isolates (MT542659~60) | 4 (S11~13,5) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 6 isolates (MT542661~62) | 4 (S11,13,5~6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 5 isolates (MT542663~64) | 3 (S13,5~6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 3 isolates (MT542665~66) | 5 (S11~13,5~6) | 4 (S14,8~10) | DR1, 2, 3 |
| 2 isolates (MT542667~68) | 2 (S11,6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 2 isolates (MT542669~70) | 4 (S11~12,5~6) | 4 (S15,8~10) | DR1, 2, 3 |
| 1 isolate (MT542671~72) | 5 (S11~13,5~6) | 7 (S14~15, 8~9, 8~10) | DR1, 2, 3 |
| 1 isolate (MT542673~74) | 4 (S11~13,5) | 5 (S14~16, 9~10) | DR1, 2, 3 |
| 1 isolate (MT542675~76) | 4 (S11~13,6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 1 isolate (MT542677~78) | 3 (S12~13,5) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 1 isolate (NTUH_4415) (MT542646) | 3 (S11,13,5) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 1 isolate (MT542679~80) | 3 (S11,5~6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 1 isolate (MT542681~82) | 2 (S11,5) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 1 isolate (MT542683~84) | 2 (S5,6) | 5 (S14~15,8~10) | DR1, 2, 3 |
| 1 isolate (MT542685~86) | 4 (S11~13,6) | 3 (S8~10) | DR1, 2, 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, J.-C.; Wan, T.-W.; Lee, H.; Wang, X.-M.; Lin, Y.-T.; Jung, C.-J.; Lee, T.-F.; Hsueh, P.-R.; Teng, L.-J. Heterogeneity of Molecular Characteristics among Staphylococcus argenteus Clinical Isolates (ST2250, ST2793, ST1223, and ST2198) in Northern Taiwan. Microorganisms 2020, 8, 1157. https://doi.org/10.3390/microorganisms8081157
Hsu J-C, Wan T-W, Lee H, Wang X-M, Lin Y-T, Jung C-J, Lee T-F, Hsueh P-R, Teng L-J. Heterogeneity of Molecular Characteristics among Staphylococcus argenteus Clinical Isolates (ST2250, ST2793, ST1223, and ST2198) in Northern Taiwan. Microorganisms. 2020; 8(8):1157. https://doi.org/10.3390/microorganisms8081157
Chicago/Turabian StyleHsu, Jia-Chuan, Tsai-Wen Wan, Hao Lee, Xiao-Mei Wang, Yu-Tzu Lin, Chiau-Jing Jung, Tai-Fen Lee, Po-Ren Hsueh, and Lee-Jene Teng. 2020. "Heterogeneity of Molecular Characteristics among Staphylococcus argenteus Clinical Isolates (ST2250, ST2793, ST1223, and ST2198) in Northern Taiwan" Microorganisms 8, no. 8: 1157. https://doi.org/10.3390/microorganisms8081157
APA StyleHsu, J.-C., Wan, T.-W., Lee, H., Wang, X.-M., Lin, Y.-T., Jung, C.-J., Lee, T.-F., Hsueh, P.-R., & Teng, L.-J. (2020). Heterogeneity of Molecular Characteristics among Staphylococcus argenteus Clinical Isolates (ST2250, ST2793, ST1223, and ST2198) in Northern Taiwan. Microorganisms, 8(8), 1157. https://doi.org/10.3390/microorganisms8081157