Molecular Characterization and Antimicrobial Susceptibilities of Nocardia Species Isolated from the Soil; A Comparison with Species Isolated from Humans

 ,
,
Abstract
Importance
1. Introduction
2. Materials and Methods
2.1. Molecular Identification of Species
2.2. Multilocus Sequence Analysis (MLSA)
2.3. Genetic Similarities Among Soil and Clinical Nocardia cyriacigeorgica Strains
2.4. Antimicrobial Susceptibilities
2.5. Bioinformatic Analysis
2.6. Accession Number(s)
3. Results
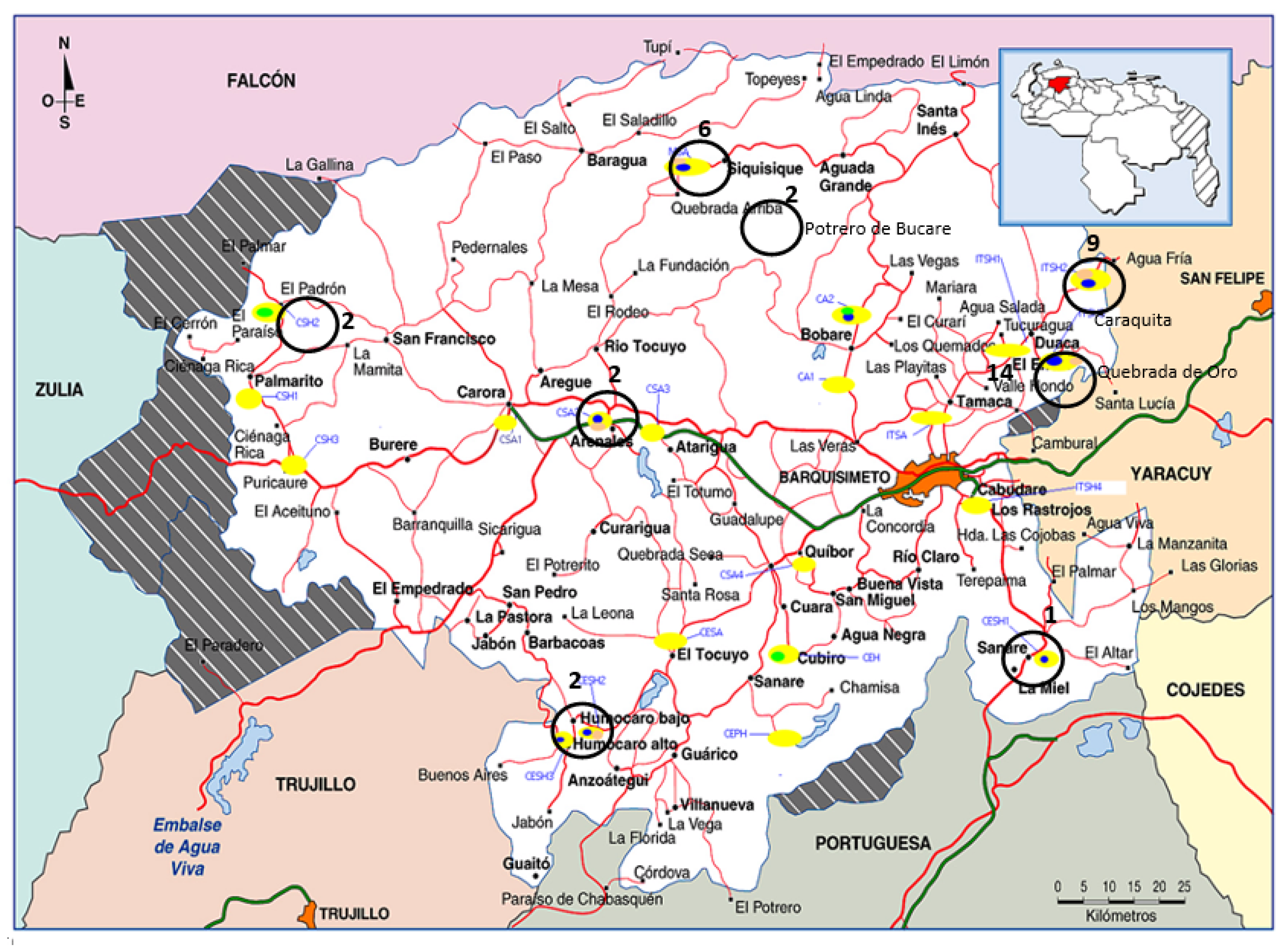
3.1. Distribution of Nocardia Species in the Soil
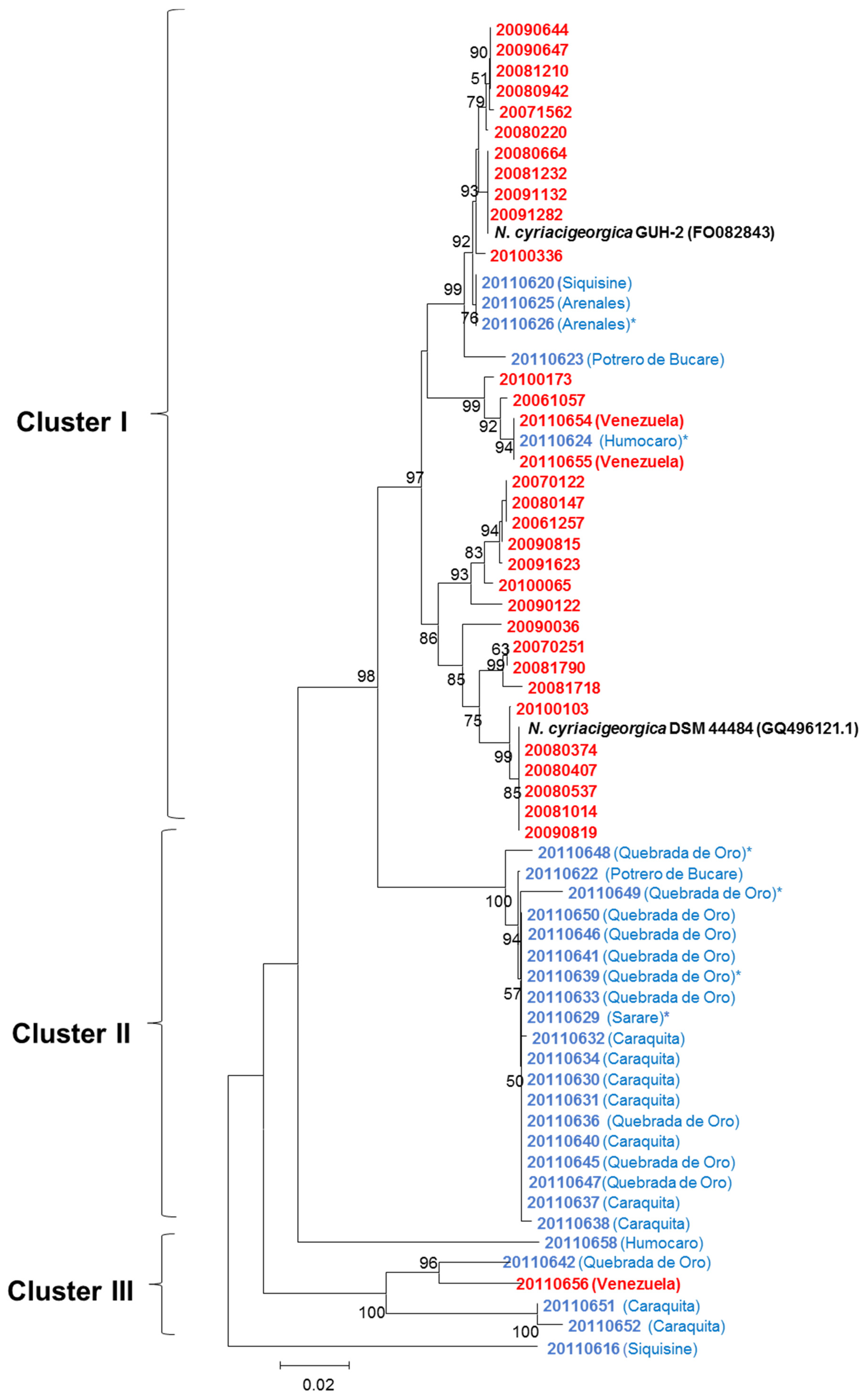
3.2. Phylogenetic Analysis by MLSA
3.3. Antimicrobial Susceptibilities
3.4. Comparison of Soil and Human N. cyriacigeorgica Strains
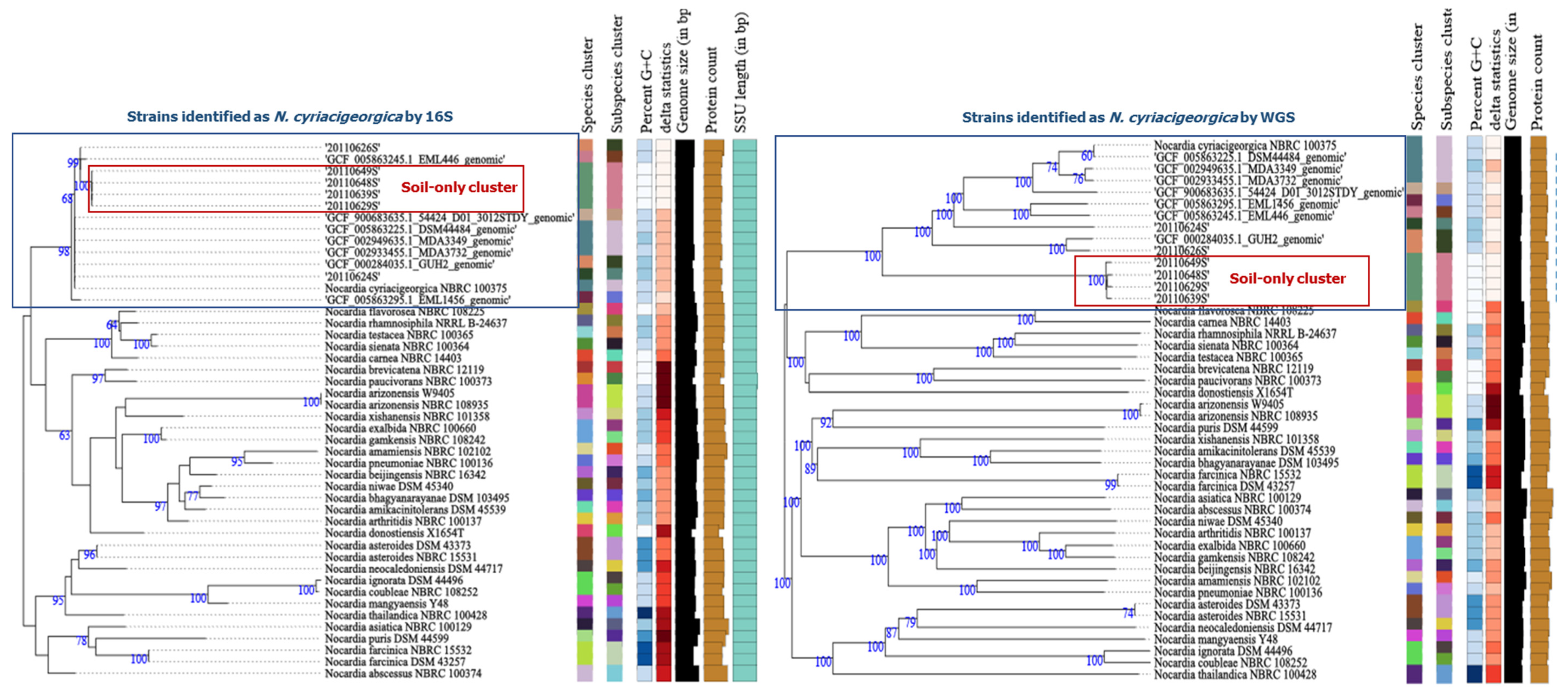
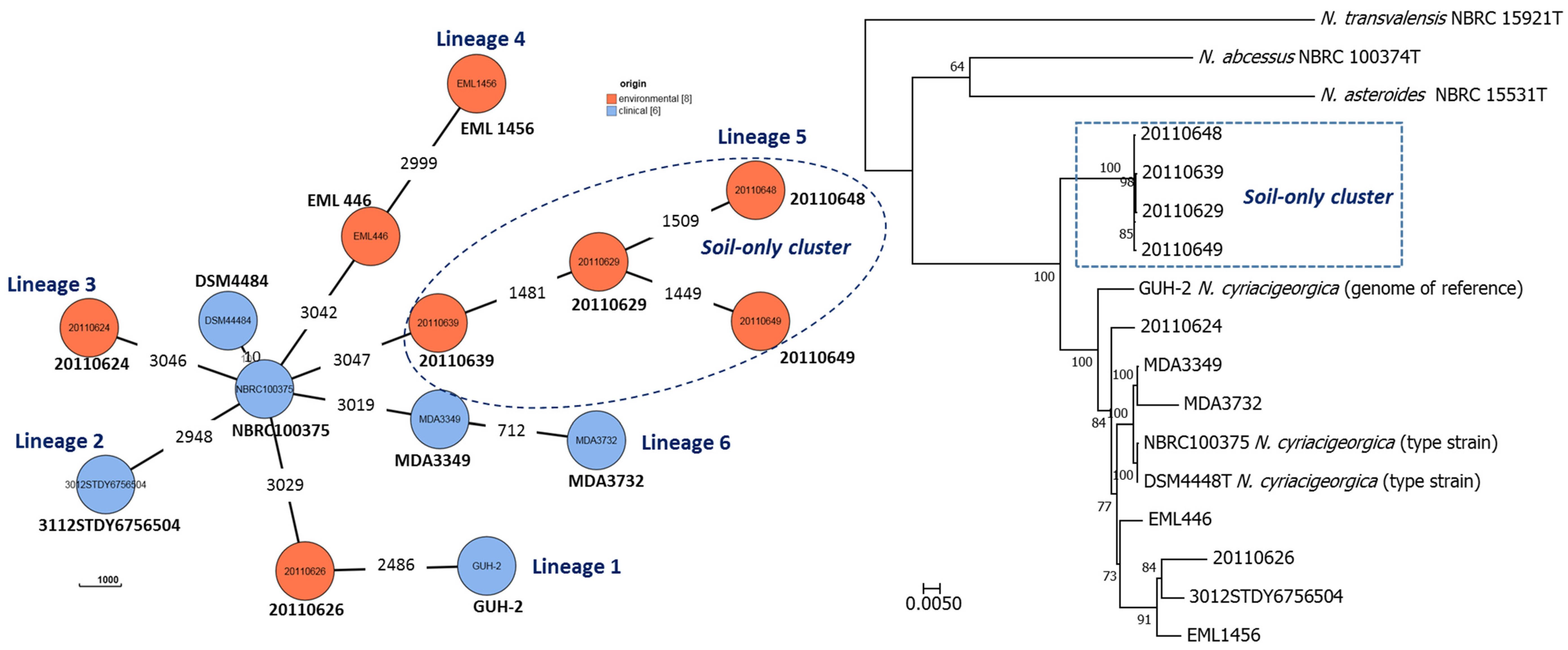
3.5. Whole-Genome Sequencing of the soil N. cyriacigeorgica Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goodfellow, M. “Family IV. Nocardiaceae”. In Bergey’s Manual of Systematic Bacteriology; Whitman, W., Goodfellow, M., Kämpfer, P., Busse, H.J., Trujillo, M., Ludwig, W., Suzuki, K., Eds.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Mohammadipanah, F.; Wink, J. Actinobacteria from Arid and Desert Habitats: Diversity and Biological Activity. Front Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef] [PubMed]
- Yaemsiri, S.; Sykes, J.E. Successful Treatment of Disseminated Nocardiosis Caused by Nocardia veterana in a Dog. J. Vet. Intern Med. 2018, 32, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tan, W.; Wang, W.; Hou, S.; Chen, G.; Xia, L.; Lu, Y. Identification of common antigens of three pathogenic Nocardia species and development of DNA vaccine against fish nocardiosis. Fish Shellfish Immunol. 2019, 95, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Yasuike, M.; Nishiki, I.; Iwasaki, Y.; Nakamura, Y.; Fujiwara, A.; Shimahara, Y.; Kamaishi, T.; Yoshida, T.; Nagai, S.; Kobayashi, T.; et al. Analysis of the complete genome sequence of Nocardia seriolae UTF1, the causative agent of fish nocardiosis: The first reference genome sequence of the fish pathogenic Nocardia species. PLoS ONE 2017, 3, 12. [Google Scholar] [CrossRef]
- Bull, A.T.; Asenjo, J.A. Microbiology of hyper-arid environments: Recent insights from the Atacama Desert, Chile. Van Leeuw J. Microb. 2013, 103, 1173–1179. [Google Scholar] [CrossRef]
- Luo, Q.; Hiessl, S.; Steinbüchel, A. Functional diversity of Nocardia in metabolism. Environ. Microbiol. 2014, 16, 29–48. [Google Scholar] [CrossRef]
- Sharma, P.; Kalita, M.C.; Thakur, D. Broad Spectrum Antimicrobial Activity of Forest-Derived Soil Actinomycete, Nocardia sp. PB-52. Front Microbiol. 2016, 7, 347. [Google Scholar] [CrossRef]
- Shivlata, L.; Satyanarayana, T. Thermophilic and alkaliphilic Actinobacteria: Biology and potential applications. Front Microbiol. 2015, 6, 1014. [Google Scholar] [CrossRef]
- Dhakal, D.; Pokhrel, A.R.; Shrestha, B.; Sohng, J.K. Marine Rare Actinobacteria: Isolation, Characterization, and Strategies for Harnessing Bioactive Compounds. Front Microbiol. 2017, 8, 1106. [Google Scholar] [CrossRef]
- Lara-Severino, R.D.; Camacho-López, M.A.; Casanova-González, E.; Gómez-Oliván, L.M.; Sandoval-Trujillo, Á.H.; Isaac-Olivé, K.; Ramírez-Durán, N. Haloalkalitolerant Actinobacteria with capacity for anthracene degradation isolated from soils close to areas with oil activity in the State of Veracruz, Mexico. Int. Microbiol. 2016, 19, 15–26. [Google Scholar] [CrossRef]
- Rodrigues, E.M.; Vidigal, P.M.P.; Pylro, V.S.; Morais, D.K.; Leite, L.R.; Roesch, L.F.W.; Tótola, M.R. Draft genome of Nocardia farcinica TRH1, a linear and polycyclic aromatic hydrocarbon-degrading bacterium isolated from the coast of Trindade Island, Brazil. Braz. J. Microbiol. 2017, 48, 391–392. [Google Scholar] [CrossRef] [PubMed]
- Brown-Elliott, B.A.; Brown, J.M.; Conville, P.S.; Wallace, R.J. Clinical and laboratory features of the Nocardia spp. based on current molecular taxonomy. Clin. Microbiol. Rev. 2006, 19, 259–282. [Google Scholar] [CrossRef] [PubMed]
- Ambrosioni, J.; Lew, D.; Garbino, J. Nocardiosis: Updated clinical review and experience at a tertiary center. Infection 2010, 38, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.; Thibert, L.; Alarie, I.; Valiquette, L.; Pépin, J. Nocardiosis in Quebec, Canada, 1988–2008. Clin. Microbiol. Infect. 2011, 17, 690–696. [Google Scholar] [CrossRef]
- Ramírez, A.; Blanco, M.; García, E. Biogeografía de Nocardia: Estudio de la población edáfica de Nocardia en diversas zonas climáticas del Estado de Lara. Rev. Soc. Ven. Microbiol. 2003, 23, 1–7. [Google Scholar]
- Carrasco, G.; Valdezate, S.; Garrido, N.; Villalón, P.; Medina-Pascual, M.J.; Sáez-Nieto, J.A. Identification, typing, and phylogenetic relationships of the main clinical Nocardia species in Spain according to their gyrB and rpoB genes. J. Clin. Microbiol. 2013, 51, 3602–3608. [Google Scholar] [CrossRef]
- Drancourt, M.; Bollet, C.; Carlioz, A.; Martelin, R.; Gayral, J.P.; Raoult, D. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol. 2000, 38, 3623–3630. [Google Scholar] [CrossRef]
- Takeda, K.; Kang, Y.; Yazawa, K.; Gonoi, T.; Mikami, Y. Phylogenetic studies of Nocardia species based on gyrB gene analyses. J. Med. Microbiol. 2010, 59, 165–171. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Susceptibility Testing of Mycobacteria, Nocardia spp., and Other Aerobic Actinomycetes, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999, 41, 95–98. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Gascuel, O. BIONJ: An improved version of the NJ algorithm based on a simple model of sequence data. Mol. Biol. Evol. 1997, 14, 685–695. [Google Scholar] [CrossRef] [PubMed]
- McTaggart, L.R.; Richardson, S.E.; Witkowska, M.; Zhang, S.X. Phylogeny and identification of Nocardia species on the basis of multilocus sequence analysis. J. Clin. Microbiol. 2010, 48, 4525–4533. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute [CLSI]. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes. Approved Standard-M24-A2, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Ant. V. Leeuw. 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Rodriguez, R.L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes (No. e1900v1). PeerJ Preprints 2016. [Google Scholar]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinf. 2013, 14, 60. [Google Scholar] [CrossRef]
- Vautrin, F.; Bergeron, E.; Dubost, A.; Abrouk, D.; Martin, C.; Cournoyer, B.; Louzier, V.; Winiarski, T.; Rodriguez-Nava, V.; Pujic, P. Genome Sequences of Three Nocardia cyriacigeorgica Strains and One Nocardia asteroides Strain. Microbiol. Resour. Announc. 2019, 15, 8. [Google Scholar] [CrossRef] [PubMed]
- Medlar, A.J.; Törönen, P.; Holm, L. AAI-profiler: Fast proteome-wide exploratory analysis reveals taxonomic identity, misclassification and contamination. Nucleic Acids Res. 2018, 46, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.M.; Kim, C.K.; Roh, J.; Byun, J.H.; Yang, S.J.; Choi, S.B.; Chun, J.; Yong, D. Application of the Whole Genome-Based Bacterial Identification System, TrueBac ID, Using Clinical Isolates That Were Not Identified With Three Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) Systems. Ann. Lab. Med. 2019, 39, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Ankenbrand, M.J.; Keller, A. bcgTree: Automatized phylogenetic tree building from bacterial core genomes. Genome 2016, 59, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Alanjary, M.; Kronmiller, B.; Adamek, M.; Blin, K.; Weber, T.; Huson, D.; Philmus, B.; Ziemert, N. The Antibiotic Resistant Target Seeker (ARTS), an exploration engine for antibiotic cluster prioritization and novel drug target discovery. Nucleic Acids Res. 2017, 45, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Guitor, A.K.; Raphenya, A.R.; Klunk, J.; Kuch, M.; Alcock, B.; Surette, M.G.; McArthur, A.G.; Poinar, H.N.; Wright, G.D. Capturing the Resistome: A Targeted Capture Method To Reveal Antibiotic Resistance Determinants in Metagenomes. Antimicrob. Agents Chemother. 2019, 20, 64. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Inouye, M.; Dashnow, H.; Raven, L.A.; Schultz, M.B.; Pope, B.J.; Tomita, T.; Zolber, J.; Holt, K.E. SRST2: Rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014, 6, 90. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, 16–21. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Interpretive Criteria for Identification of Bacteria and Fungi by DNA Target Sequencing: Approved Guideline MM18-A; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Brown-Elliott, B.A.; Conville, P.; Wallace, R.J. Current Status of Nocardia Taxonomy and Recommended Identification Methods. Clin. Microbiol. Newsl. 2015, 37, 25–32. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef]
- Ott, S.R.; Meier, N.; Kolditz, M.; Bauerd, T.T.; Rohde, G.; Presterl, E.; Schürmann, D.; Lepper, P.M.; Ringshausen, F.C.; Flick, H.; et al. Pulmonary nocardiosis in Western Europe-Clinical evaluation of 43 patients and population-based estimates of hospitalization rates. Int. J. Inf. Dis. 2019, 81, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Kachuei, R.; Emami, M.; Mirnejad, R.; Khoobdel, M. Diversity and frequency of Nocardia spp. in the soil of Isfahan province, Iran. Asian Pac. J. Trop. Biomed. 2012, 2, 474–478. [Google Scholar] [CrossRef]
- Aghamirian, M.; Ghiasian, S.A. Isolation and characterization of medically important aerobic actinomycetes in soil of Iran (2006-2007). Open Microbio. L J. 2009, 3, 53–57. [Google Scholar] [CrossRef][Green Version]
- Lebeaux, D.; Bergeron, E.; Berthet, J.; Djadi-Prat, J.; Mouniée, D.; Boiron, P.; Lortholary, O.; Rodriguez-Nava, V. Antibiotic susceptibility testing and species identification of Nocardia isolates: A retrospective analysis of data from a French expert laboratory, 2010–2015. Clin. Microbiol. Infec. 2019, 25, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, A.; Yazawa, K.; Ishikawa, J.; Hotta, K.; Nishimura, K.; Mikami, Y. Nocardial infections in Japan from 1992 to 2001, including the first report of infection by Nocardia transvalensis. Eur. J. Epidemiol. 2004, 19, 383–389. [Google Scholar] [CrossRef]
- Valdezate, S.; Garrido, N.; Carrasco, G.; Medina-Pascual, M.J.; Villalón, P.; Navarro, A.M.; Sáez-Nieto, J.A. Epidemiology and susceptibility to antimicrobial agents of the main Nocardia species in Spain. J. Antimicrob. Chemother. 2017, 72, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Conville, P.S.; Brown-Elliott, B.A.; Smith, T.; Zelazny, A.M. The Complexities of Nocardia Taxonomy and Identification. J. Clin. Microbiol. 2017, 56, e01419-17. [Google Scholar] [CrossRef]
- Valero-Guillén, P.L.; Martín-Luengo, F. Nocardia in soils of southeastern Spain: Abundance, distribution, and chemical characterisation. Can. J. Microbiol. 1984, 30, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.F.; Werth, J.T. Is the lower atmosphere a readily accessible reservoir of culturable, antimicrobial compound-producing Actinomycetales? Front Microbiol. 2015, 6, 802. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Wang, P.; Yang, C.; Gu, L. Molecular identification and phylogenetic relationships of clinical Nocardia isolates. Ant. V. Leeuw. 2019, 112, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.C.; Chimetto, L.; Edwards, R.A.; Swings, J.; Stackebrandt, E.; Thompson, F.L. Microbial genomic taxonomy. BMC Genom. 2013, 14, 913. [Google Scholar] [CrossRef] [PubMed]
- Varghese, N.J.; Mukherjee, S.; Ivanova, N.; Konstantinidis, K.T.; Mavrommatis, K.; Kyrpides, N.K.; Pati, A. Microbial species delineation using whole genome sequences. Nucleic Acids Res. 2015, 43, 6761–6771. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.L.M.; Gunturu, S.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. The Microbial Genomes Atlas (MiGA) webserver: Taxonomic and gene diversity analysis of Archaea and Bacteria at the whole genome level. Nucleic Acids Res. 2018, 46, 282–288. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No. | Species (16S rDNA) | Percentage Identity with Respect to DMS 44484T 16S rDNA | Percentage Identity with Respect to DMS 44484T gyrB | Drug Resistance Phenotype | Location (Municipality) | Latitude (N)/Longitude(W) and Altitude | Temperature/Rainfall | Sample Time (m, yr) | Weather Type |
|---|---|---|---|---|---|---|---|---|---|
| 20110625 | N. cyriacigeorgica | 99.84% | 99.4% | CIP CLA | Arenales (Torres) | 10° 9′11" | 27 °C/400mm | August, 2002 | Semi-arid continental |
| 20110626 b,c,d | N. cyriacigeorgica | 100% | 95.9% | xl CIP CLA | Arenales | 69° 54´12" 517m | |||
| 20110630 | N. cyriacigeorgica | 99.84% | 93.1% | Xl CLA | Caraquita(Crespo) | 10° 41′ 11" 69° 05′ 11" 685m | 25 °C/743mm | April, 2006 | Subhumid interior (transitional) |
| 20110631 | N. cyriacigeorgica | 99.84% | 93.1% | Xl CLA | Caraquita | ||||
| 20110632 | N. cyriacigeorgica | 99.84% | 93.0% | XL cla | Caraquita | ||||
| 20110634 | N. cyriacigeorgica | 99.84% | 93.1% | XL CLA | Caraquita | ||||
| 20110637 | N. cyriacigeorgica | 99.84% | 93.1% | CLA | Caraquita | ||||
| 20110638 | N. cyriacigeorgica | 99.84% | 92.8% | xl | Caraquita | ||||
| 20110640 | N. cyriacigeorgica | 98.85% | 93.1% | CLA | Caraquita | ||||
| 20110651 a,c | N. cyriacigeorgica | 99.84% | 86.2% | XL CLA CIP | Caraquita | ||||
| 20110652 a,c | N. vermiculata | - | - | - | Caraquita | ||||
| 20110643 a,c | N. vermiculata | - | - | CIP | El Padrón (Torres) | 10° 20′ 44″ 70° 28′ 59′″ 643m | 26 °C/921mm | May, 2006 | Subhumid continental |
| 20110644 a,b,c | N. elegans | - | - | XL TOB CIP | El Padrón | ||||
| 20110624 d | N. cyriacigeorgica | 99.92% | 95.9% | CLA | Humocaro (Morán) | 9o 40´57" 69o 58 12" 964m | 24 °C/700mm | August, 2002 | Subhumid continental (seasonal) |
| 20110658 | cyriacigeorgica | 99.84% | 87.9% | Xl CLA | Humocaro | ||||
| 20110622 | N. cyriacigeorgica | 99.84% | 93.3% | CLA CIP | Potrero de Bucare (Iribarren) | 10o 18´51" 69o 27´45" 711m | 24 °C/700mm 25 °C/339mm | Semi-arid continental | |
| 20110623 | N. cyriacigeorgica | 99.75% | 98.2% | CIP | August, 2002 | ||||
| 20110627 a,c | N. abcessus | - | - | IMI CIP | Quebrada de Oro (Crespo) | 10° 16′ 2" 69° 2′ 22" 1278m | 24 °C/1285mm | April, 2006 | Subhumid interior (transitional) |
| 20110628 | N. abcessus | - | - | IMI CIP | Quebrada de Oro | ||||
| 20110633 | N. cyriacigeorgica | 99.84% | 93.1% | CLA | Quebrada de Oro | ||||
| 20110635 a,c | N. asteroides | - | - | - | Quebrada de Oro | ||||
| 20110636 | N. cyriacigeorgica | 99.92% | 93.1% | CIP cla | Quebrada de Oro | ||||
| 20110639 d | N. cyriacigeorgica | 99.92% | 92.6% | CIP CLA | Quebrada de Oro | ||||
| 20110641 | N. cyriacigeorgica | 99.84% | 93.1% | CLA | Quebrada de Oro | ||||
| 20110642 a,c | N. cyriacigeorgica | 99.84% | 85.1% | xl | Quebrada de Oro | ||||
| 20110645 | N. cyriacigeorgica | 99.84% | 93.1% | XL CLA min | Quebrada de Oro | ||||
| 20110646 | N. cyriacigeorgica | 99.84% | 93.1% | XL CLA CIP | Quebrada de Oro | ||||
| 20110647 | N. cyriacigeorgica | 100% | 93.1% | CLA | Quebrada de Oro | ||||
| 20110648 d | N. cyriacigeorgica | 99.84% | 92.8% | CLA | Quebrada de Oro | ||||
| 20110649 d | N. cyriacigeorgica | 99.84% | 91.2% | CLA | Quebrada de Oro | ||||
| 20110650 | N. cyriacigeorgica | 99.92% | 93.1% | XL, CLA | Quebrada de Oro | ||||
| 20110629 d | N. cyriacigeorgica | 99.75% | 92.4% | CLA | Sarare (Simón Planas) | 9° 47′ 2″ 69° 9′ 40″ 269m | 26 °C/1434mm | August, 2002 | Subhumid continental (seasonal) |
| 20110616 b,c | N. cyriacigeorgica | 100% | 85.0% | CIP, CLA | Siquisique (Urdancia) | 10o 34´24" 69o 42´ 5" 271m | 27 °C/358mm | August, 2002 | Semi-arid continental |
| 20110617 a,c | N. rhamnosiphila | - | - | CLA | Siquisique | ||||
| 20110618 a,c | N. rhamnosiphila | - | - | - | Siquisique | ||||
| 20110619 | N. cyriacigeorgica | 99.92% | 99.4% | XL CLA CIP | Siquisique | ||||
| 20110620 | N. cyriacigeorgica | 100% | 99.4% | XL CLA | Siquisique | ||||
| 20110621 a,b | N. mexicana | - | - | Xl IMI tob CLA min | Siquisique |
| Antimicrobial Agent | MIC (mg/L) 1 | Resistance (%) 2–4 | Sign. Difference | ||
|---|---|---|---|---|---|
| Range | MIC50 | MIC90 | (p ≤0.05) | ||
| Amoxicillin-clavulanic acid 4 | |||||
| Soil | ≤2–32 | 8 | 32 | 14 (48.27%) | yes |
| Clinical | ≤2–64 | 32 | 32 | 23 (76.7%) | |
| Cefoxitin | |||||
| Soil | ≤4–128 | 8 | 32 | 12 (41.4%) 5 | yes |
| Clinical | ≤4–≥128 | 128 | ≥128 | 28 (93.4%) 5 | |
| Ceftriaxone | |||||
| Soil | ≤4 | ≤4 | ≤4 | 0 | yes |
| Clinical | ≤4–16 | ≤4 | 8 | 5 (16.7%) | |
| Cefepime | |||||
| Soil | ≤1–16 | 2 | 8 | 1 (3.4%) | yes |
| Clinical | ≤1–32 | 16 | 32 | 18 (60.0%) | |
| Imipenem | |||||
| Soil | ≤2–4 | ≤2 | ≤2 | 0 | yes |
| Clinical | ≤2–32 | 8 | 32 | 24 (79.2%) | |
| Amikacin | |||||
| Soil | ≤1–16 | ≤1 | 2 | 1 (3.4%) | no |
| Clinical | ≤1–16 | ≤1 | ≤1 | 1 (3.0%) | |
| Tobramycin | |||||
| Soil | ≤1–2 | ≤1 | ≤1 | 0 | no |
| Clinical | ≤1–16 | ≤1 | ≤1 | 2 (6.6%) | |
| Ciprofloxacin | |||||
| Soil | ≤0.12–≥4 | 1 | ≥4 | 8 (28.5.0%) | yes |
| Clinical | 2–≥4 | ≥4 | ≥4 | 29 (96.7%) | |
| Moxifloxacin | |||||
| Soil | ≤0.25–≥4 | 0.5 | 4 | 10 (34.5%) | yes |
| Clinical | 1–≥4 | 4 | ≥4 | 29 (96.7%) | |
| Clarithromycin | |||||
| Soil | 1–≥16 | 8 | 16 | 28 (96.5%) | yes |
| Clinical | ≤0.06–≥16 | ≥16 | ≥16 | 22 (73.3%) | |
| Doxycycline | |||||
| Soil | ≤0.25–8 | 2 | 4 | 17 (58.6%) | no |
| Clinical | ≤0.12–8 | 2 | 4 | 19 (62.7%) | |
| Minocycline | |||||
| Soil | ≤1–8 | ≤1 | 2 | 7 (24.1%) | yes |
| Clinical | ≤1–4 | 2 | 4 | 18 (59.4%) | |
| Tigecycline | |||||
| Soil | 0.06–4 | 0.25 | 1 | -6 | no |
| Clinical | ≤0.25–≥4 | 0.25 | 2 | -6 | |
| Co-trimoxazole 4 | |||||
| Soil | ≤0.25–0.5 | 0.25 | 0.5 | 0 | no |
| Clinical | ≤0.25–4 | 0.5 | 2 | 1 (3.3%) | |
| Linezolid | |||||
| Soil | ≤1–2 | ≤1 | ≤1 | 0 | no |
| Clinical | ≤1–4 | ≤1 | 2 | 0 | |
| Strain (ID/refSeq) | G + C% Length (no. of Contigs; Depth Coverage) | 16S rRNA (≥ 99.6%) 1,2 | gyrB (≥ 93.5%) 1,2 | ANI (≥ 95%) 1,2,3 | AAI (≥ 95%??) 1,2,4 | DDH-Estimate (GLM-based) (≥70% DDH-Estimate, Difference in <1% G + C) 1,2,5 and Interpretation | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain for Comparison | GUH-2 | DSM 44484T | GUH-2 | DSM 44484T | GUH-2 | DSM 44484T | GUH-2 | DSM 44484T | GUH-2 | DSM 44484T | |
| GUH-2 NC_016887 | 68.37% 6,194,645 (1) | -- | 100 | -- | 94.77 | -- | 90.14 | -- | 92.08 | -- | 99.9% (99.8–100%) 0.03 (either distinct or same species) |
| DSM 44484T NZ_VBUR00000000.1 | 68.19% 6,311,306 (1, 484x) | 100 | -- | 94.77 | -- | 90.14 | -- | 92.08 | - | 99.9% (99.8–100%) 0.03 (either distinct or same species) | -- |
| Soil Strains | |||||||||||
| 20110624 JAAGVC000000000 | 68.39% 6,326,508 (113;79x) | 100 | 100 | 95.32 | 95.98 | 89.84 | 91.12 | 91.12 | 93.11 | 38.0% (36.3–41.3%) 0.05 (either distinct or same species) | 43.6% (41.1–46.1%) 0.10 (either distinct or same species) |
| 20110626 JAAGVB000000000 | 68.29% 6,578,812 (158;100x) | 99.79 | 99.87 | 99.45 | 95.34 | 97.84 | 90.64 | 91.74 | 92.03 | 86.0% (83.4–88.3%) 0.1 (either distinct or same species) | 41.2% (38.7–3.8%) 0.04 (either distinct or same species) |
| 20110629 JAAGVA000000000 | 66.87% 6,251,294 (71;154x) | 99.79 | 99.87 | 93.12 | 92.37 | 86.77 | 86.38 | 88.21 | 88.23 | 31.5% (29.1–34%) 1.75 (distinct species) | 31.4% (29–33.9%) 1.61 (distinct species) |
| 20110639 JAAGUZ000000000 | 66.95% 6,200,016 (178;45x) | 99.79 | 99.87 | 93.12 | 92.6 | 86.57 | 85.85 | 88.22 | 88.30 | 31.6% (29–34.1%), 1.43 (distinct species) | 31.5% (29.1–34%) 1.29 (distinct species) |
| 20110648 JAAGUY000000000 | 66.96% 6,274,061 (57;126x) | 99.79 | 99.87 | 92.85 | 92.85 | 86.62 | 85.68 | 88.21 | 88.21 | 31.6% (29.2–34.1%) 1.42 (distinct species) | 31.5% (29.1–34%) 1.28 (distinct species) |
| 20110649 JAAGUX000000000 | 66.92% 6,258,095 (135;46x) | 99.79 | 99.87 | 92.16 | 91.20 | 86.72 | 85.81 | 88.21 | 88.22 | 31.6% (29.2–34.1%) 1.46 (distinct species) | 31.5% (29.1–34%) 1.31 (distinct species) |
| Strains with available genome | |||||||||||
| 3012STDY6756504 NZ_LR215973.1 | 68.20% 6,476,621 (1535;100x) | 100 | 100 | 96.84 | 97.93 | 89.96 | 96.26 | 92.26 | 96.82 | 39.9% (37.4–42.4%) 0.13 (either distinct or same species) | 68.8% (65.8–71.6%) 0.04 (either distinct or same species) |
| EML 446 NZ_VBUT00000000.1 | 68.20% 6,520,205 (14;463x) | 100 | 100 | 97.08 | 97.03 | 90.34 | 91.99 | 92.42 | 93.72 | 41.1% (38.6–43.6%) 0.14 (either distinct or same species) | 47.0% (44.4–49.6%) 0.03 (either distinct or same species) |
| EML 1456 NZ_VBUU00000000.1 | 68.00% 6,830,276 (108;458x) | 100 | 100 | 96.94 | 96.95 | 90.26 | 92.07 | 92.37 | 93.73 | 40.9% (38.4–43.4%) 0.34 (either distinct or same species) | 47.2% (44.6–9.8%) 0.17 (either distinct or same species) |
| MDA3349 NZ_CP026746.1 | 68.30% 6,462,637 (9; 43x) | 100 | 100 | 96.55 | 99.84 | 90.21 | 97.83 | 92.30 | 98.00 | 41.3% (38.8–3.9%) 0.15 (either distinct or same species) | 81.2% (78.3–83.8%) 0.09 (either distinct or same species) |
| MDA3732 NZ_PSZF00000000.1 | 68.29% 6,592,249 (84;172x) | 100 | 100 | 94.32 | 96.97 | 90.56 | 97.62 | 92.46 | 97.87 | 39.9% (37.4–42.4%) 0.13 (either distinct or same species) | 80.3% (77.4–83%) 0.03 (either distinct or same species) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrasco, G.; Monzón, S.; San Segundo, M.; García, E.; Garrido, N.; Medina-Pascual, M.J.; Villalón, P.; Ramírez, A.; Jiménez, P.; Cuesta, I.; et al. Molecular Characterization and Antimicrobial Susceptibilities of Nocardia Species Isolated from the Soil; A Comparison with Species Isolated from Humans. Microorganisms 2020, 8, 900. https://doi.org/10.3390/microorganisms8060900
Carrasco G, Monzón S, San Segundo M, García E, Garrido N, Medina-Pascual MJ, Villalón P, Ramírez A, Jiménez P, Cuesta I, et al. Molecular Characterization and Antimicrobial Susceptibilities of Nocardia Species Isolated from the Soil; A Comparison with Species Isolated from Humans. Microorganisms. 2020; 8(6):900. https://doi.org/10.3390/microorganisms8060900
Chicago/Turabian StyleCarrasco, Gema, Sara Monzón, María San Segundo, Enrique García, Noelia Garrido, María J. Medina-Pascual, Pilar Villalón, Ana Ramírez, Pilar Jiménez, Isabel Cuesta, and et al. 2020. "Molecular Characterization and Antimicrobial Susceptibilities of Nocardia Species Isolated from the Soil; A Comparison with Species Isolated from Humans" Microorganisms 8, no. 6: 900. https://doi.org/10.3390/microorganisms8060900
APA StyleCarrasco, G., Monzón, S., San Segundo, M., García, E., Garrido, N., Medina-Pascual, M. J., Villalón, P., Ramírez, A., Jiménez, P., Cuesta, I., & Valdezate, S. (2020). Molecular Characterization and Antimicrobial Susceptibilities of Nocardia Species Isolated from the Soil; A Comparison with Species Isolated from Humans. Microorganisms, 8(6), 900. https://doi.org/10.3390/microorganisms8060900